QUICK SEARCH
MO PROJECTS:
Africa
Asia/Pacific
Mesoamerica
North America
South America
General Taxonomy
Photo Essays
Training in Latin
America
MO RESEARCH:
Wm. L. Brown Center
Bryology
GIS
Graduate Studies
Research Experiences
for Undergraduates
Imaging Lab
Library
MBG Press
Publications
Climate Change
Catalog Fossil Plants
MO DATABASES:
W³MOST
Image Index
Rare Books
Angiosperm
Phylogeny
Res Botanica
All Databases
INFORMATION:
What's New?
People at MO
Visitor's Guide
Herbarium
Jobs & Fellowships
Symposium
Research Links
Site Map
Search
Synopsis of Chrysochlamys (Clusiaceae: Clusioideae: Clusieae) in MesoamericaBarry E. Hammel
Abstract. Delimitation of genera in the Tovomita group of tribe Clusieae has fluctuated, in Central America, with almost every new floristic work. Nevertheless, with some aberrations, the theme of a unified Chrysochlamys Tovomitopsis has persisted, and is here supported by concurring with Bentham and Hooker's placement of Tovomitopsis in synonymy of Chrysochlamys. The strictly South American Balboa is also placed in synonymy of Chrysochlamys, which action necessitates a new name (here provided) for its only species. A key to the 13 species of Chrysochlamys recognized for Mexico and Central America is provided. An annotated list of all, including brief descriptions, taxonomic and nomenclatural notes, five new combinations, and two new species, brings forth that concept and rectifies the nomenclature to the practicalities of numerous regional floras. Tovomita croatii Maguire, a somewhat problematic species, transferred to Tovomitopsis by D'Arcy, is included in the key to Chrysochlamys and fully discussed. The Tovomita group of the tribe Clusieae (Clusiaceae, subfamily Clusioideae) comprises primarily arborescent, terrestrial genera with succulent-capsuled fruits and 1(rarely 2) large arillate seeds per locule. Three genera therein, Tovomita, Chrysochlamys, and Tovomitopsis have suffered a particularly tortuous taxonomic history. Planchon and Triana (1860) made the genus Tovomitopis to accommodate Sprengel's Bertolonia, a later homonym; Bertolonia Raddi (Melastomataceae) is now conserved over Bertolonia Spin (Myoporaceae) (Farr et al., 1979). Planchon and Triana distinguished Tovomitopsis from Chrysochlamys claiming it had an arilloid (originating from the micropyle) rather than the true aril (originating from the funicule) they claimed for Chrysochlamys. According to them, the so-called aril of Tovomita is neither, but rather the outer integument of the seed. These differences were further supported by floral structure: Tovomitopis, calyx 4-merous; Chrysochlamys, calyx 5-merous; Tovomita, calyx 2- or 4-parted, the outer two sepals overtopping the bud. Planchon and Triana also described Balboa in the same publication with Tovomitopsis, but they placed it in their subtribe "Euclusiae" rather than Tovomiteae. Bentham and Hooker (1862: 167--176) recognized Balboa, Chrysochlamys and Tovomita (as related) but treated Tovomitopsis as a synonym of Chrysochlamys. However, they transferred no species. Subsequently, Balboa was apparently forgotten in discussions relating to the Tovomita group. Hemsley (1879: 87) transferred all Central American species of Tovomitopsis to Chrysochlamys. Cuatrecasas (1950) also declared unjustified the separation between Chrysochlamys and Tovomitopsis and transferred two (South American) species to Chrysochlamys. Williams (1959) subsumed Tovomitopsis within Tovomita, but kept Chrysochlamys separate. Maguire (1977) tentatively recognized all three genera. D'Arcy (1980) expressed the same taxonomic opinion begun by Bentham and Hooker and put into practice by Hemsley and Cuatrecasas, i.e., a unified Chrysochlamys/Tovomitopsis, but used the later name, Tovomitopsis. D'Arcy also elevated Engler's section Dystovomita of Tovomita to the genus Dystovomita. Most recently, an attempt has been made to circumscribe Chrysochlamys as comprising species with succulent petals, echinate pollen exine, and black seeds with white arils, rather than membranaceous petals, psilate-foveolate pollen, and green seeds with orange arils (cf. Hammel, 1984, 1986). Ignoring whether or not these characters support a monophyletic group or even if they coincide, typologically, with Chrysochlamys, neither such a separation, nor one based on the considerations of Planchon and Triana is practical. Dystovomita, with axillary inflorescences (i.e., mostly lateral and below the leaves), large, adaxial petiolar pits and relatively small seeds, is easily recognized and may even lie closer to Clusia (P. Stevens, in prep.). Tovomita, with terminal inflorescences, leaves clustered at the ends of side shoots, leaves sometimes with petiolar pits, outer two sepals fused in bud or at least overtopping the bud, styles often evident, fruits usually with the inner wall and placenta dark reddish wine, can also be recognized, even with most herbarium material. The older name, Chrysochlamys, takes precedence over Tovomitopsis for the remaining species in the group, i.e., those with terminal inflorescences, leaves not clustered on side shoots, lacking adaxial petiolar pits, sepals (usually) not overtopping the bud, styles lacking, and the inner fruit wall and placenta mostly white or pink. Balboa is a synonym of Chrysochlamys. The above paragraph can be summarized by the following key:
Chrysochlamys Poeppig, Nov. Gen. Sp. Pl.. 3:13, t. 211. 1842. TYPE: Chrysochlamys multiflora Poeppig. Balboa Planchon & Triana, Ann. Sci. Nat. Bot., ser. 4, 13: 315, 14: 252. 1860. TYPE: Balboa membranacea Planchon & Triana. Tovomitopsis Planchon & Triana, Ann. Sci. Nat. Bot., ser 4, 14: 261. 1860. TYPE: T. paniculata (Sprengel) Planchon & Triana. Basionym: Bertolonia paniculata Sprengel. Dioecious, terrestrial trees or shrubs, sometimes epipetric rheophytes, the branching decussate and the leaves dispersed more or less evenly along the branches; resin mostly clear, occasionally white, rarely yellow; internodes mostly without bud-scale scars. Petiolar pits and interpetiolar stipuliform structures absent. Leaves opposite, the blades elliptic; major lateral veins mostly prominent, resin canals sometimes visible in dried material. Inflorescences terminal (rarely left pseudo-axillary by the dominant growth of an axillary, vegetative bud), many-flowered, panicles. Flowers 4- or 5-merous, corolla aestivation imbricate, petals thin, membranaceous, to thick and succulent, pink, or yellow to white. Staminate flowers with numerous (rarely few) ± separate stamens, or the filaments fused basally into a short column, or rarely with fertile stamens surrounding a central sterile capitulum, or very rarely the anthers all sessile on a resiniferous capitulum. Pistillate flowers with numerous separate staminodia, or staminodia mounted on a ring or rarely just a ring surrounding the ovary; ovary with mostly 4 or 5 stigmas, rarely 3. Fruits globose to ovoid, usually terete, rarely winged or star-shaped in cross section, succulent capsules mostly with 1 seed maturing per locule, sometimes only 1 seed per fruit; seeds green to nearly black with an orange or white aril. Distribution and ecology. Chrysochlamys is known from Mexico to Bolivia and Brazil, where it occurs in wet to very wet primary forest from near sea level to at least 2300 m. Chrysochlamys, in total, contains perhaps as many as 50 species, most of them South American, and is in much need of revision. At least two Central American species, C. glauca (Oersted, Planchon & Triana) Hemsley and C. nicaraguensis (Oersted, Planchon & Triana) Hemsley, have been erroneously reported from South America, and numerous South American names (e.g., C. membranacea Planchon & Triana, C. micrantha Engler, C. myrcioides Planchon & Triana) have been loosely (and erroneously) applied to Central American species, as well as within South America. The following key, distinguishing 13 species of Chrysochlamys for Mexico and Central America, and the subsequent annotated list, including citation of representative specimens and descriptions of two new species from Costa Rica and Panama, aim to clarify the species and names for the region. One or two possible new species, from among Panamanian material at hand, await resolution and are not included here.
Chrysochlamys allenii (Maguire) Hammel, comb. nov. Basionym: Tovomitopsis allenii Maguire, Phytologia 38: 214. 1978. TYPE: Panama. Bocas del Toro: Robalo Trail, northern slopes of Cerro Horqueta, 2000--2300 m, Allen 5014 (holotype, NY; isotype, MO). Tovomitopsis centistaminibus D'Arcy, Ann. Missouri Bot. Gard. 67: 1030. 1980. Syn. nov. TYPE: Panama. Chiriquí: 8 km N of Camp El Volcán, 2000 m, Little 6070 (holotype, MO; isotype, US). Trees 5--25 m, sometimes with stilt roots, occasionally reported as epiphytic; resin clear to creamy. Leaf blades elliptic to slightly obovate, widest just above the middle, 11--16 X 4--6 cm, the apex rounded to acuminate, the base acuminate, coriaceous; lateral veins ca. 0.8--1 cm apart, ca. 9--12 pairs, reaching to the margin; petiole 1--2 cm. Inflorescence 8--12 cm, erect, puberulent; flower buds ca. 5--7 mm; stamens 50 or more, anthers of both stamens and staminodia about as wide as long, < 1 mm, filaments free; stigmas large (ca. 2 mm), distinctly spreading. Fruits pyriform, 2--2.5 cm, yellow-green, the stigmas often mounted slightly to one side; calyx deciduous in fruit. Distribution. Very wet cloud forest, (700--)1450--2300 m; continuous from the Cordillera de Guanacaste in northwestern Costa Rica to Chiriquí province of western Panama, disjunct from Darién province of far eastern Panama. A larger tree than any other Chrysochlamys in Central America and growing at higher elevation, C. allenii is further distinguished by its coriaceous leaves, numerous stamens (more than 50), often very large stigmas, and pyriform, often greenish yellow fruits. Although D'Arcy (1980) compared Tovomitopsis centistaminibus with T. psychotriifolia, the type specimen's high-elevation habitat, large stature, coriaceous leaves, and numerous stamens conspire to alienate it from the latter species, and instead, to bring it into synonymy with C. allenii. This species, however, can at times be confused with C. psychotriifolia, which usually grows at somewhat lower elevations, has fewer (and often longer) stamens, and larger fruits. Material at hand, especially that from far eastern Panama, suggests that C. allenii may be closely related to, perhaps not distinct from, C. colombiana (Cuatrecasas) Cuatrecasas. Representative specimens. Costa Rica. Guanacaste: Volcán Cacao, 1100 m, Nov. 1990 (fr), Espinoza 24 (INB). Puntarenas: Monteverde Reserve, 1500 m, June 1985 (fl, fr), Hammel 13856 (MO); cloud forest above Wilson's finca S of San Vito de Java, 1600 m, Aug. 1967 (fr), Raven 21845 (MO). Heredia: 12 km above San Rafael, 1800 m, July 1967 (fl), Lent 1141 (MO). San José: La Palma, 1550 m, Aug 1898 (fl), Tondúz 7378 (MO). Panama. Chiriquí: 8 km N camp El Volcán, 2000 m, Mar. 1943 (fr), Little 6068 (MO); about 2 km NE of Guadelupe, 2 km N of Cerro Punta, Dec. 1971 (fr), Wilbur et al 15331 (DUKE, MO); Bajo Mono - Robalo trail, 1600--2300 m, July 1947 (fl),Allen 4839 (MO). Bocas del Toro: vicinity of Cerro Colorado, 1700 m, Apr. 1986 (fr), McPherson 8925 (MO); vicinity of Fortuna Dam, 1200 m, Apr. 1986 (fl), McPherson9049 (MO). Darién: Cerro Sapo, 800--1000 m, Mar. 1986 (fl, fr), Hammel et al 14877 (MO). Chrysochlamys angustifolia (Maguire) Hammel, comb. nov. Basionym: Tovomitopsis angustifolia Maguire, Phytologia 36: 399. 1977. TYPE: Panama. Veraguas, Mori & Kallunki 5357 (holotype, MO; isotype, NY). Small shrubs to 2 m, often on rocks along streams; resin milky. Leaf blades narrowly elliptic 7--13 X 1--2(2.5) cm, the apex acute to acuminate, base acute, membranaceous, drying light gray- or tannish green; lateral veins obscure, ca. 0.5--1 cm apart, ca. 5 or 6 pairs, reaching to the margin; petiole ca. 0.5--1 cm. Inflorescence ca. 6 cm, erect, glabrous; flower buds 6--9 mm; stamens 30--40, anthers of stamens and staminodia ± triangular, slightly longer than wide, ca. 1 mm, filaments free; stigmas spreading to often erect. Fruits pyriform, 1.5--2 cm, red or greenish red; calyx deciduous in fruit. Distribution. Very wet, mostly lowland forest, usually along streams, 100--1000 m; Bocas del Toro to Veraguas and Coclé provinces of central Panama. This distinctive, small, narrow-leaved, epipetric rheophyte with flower buds over 6 mm long is endemic to a few lowland or mid-elevation sites in west central Panama. Its habitat, larger flowers, and stouter fruiting rachis easily distinguish it from the similarly narrow-leaved new species, C. tenuis, described below. Among Central American species, C. angustifolia is one of the few with copious milky resin rather than the more common clear to tardily creamy resin. Representative specimens. PANAMA. Bocas del Toro: Upper Río San Pedro, 500 m, July 1979 (fr), Gordon 59Da (MO). Veraguas: vicinity of Santa Fe, dirt road past Escuela Alto de Piedra, 490 m, June 1987 (fl, fr), Croat 66871 (MO). Coclé: along Río San Juan near fork with Río Tife, 400 m, June 1978 (fl, fr), Hammel 3329 (MO). Chrysochlamys eclipes L. O. Williams, Trop. Woods 111: 15. 1959. TYPE: Panama. Canal Zone: Barro Colorado Island, Standley 31459 (holotype, US). Shrubs or trees 2--10 m; resin clear. Leaf blades elliptic to oblong, 12--26(--29) X 5--12 cm, the apex acute, base acute, membranaceous, drying dark reddish tan or rarely greenish tan; lateral veins obvious, 1--1.5 cm apart, 10--12 pairs, loop-connected to a submarginal vein ca. 1--3 mm from the margin; petiole ca. 1--3 cm. Inflorescence ca. 7--21 cm, erect or pendulous, minutely puberulous; flower buds ca. 4--5 mm; stamens ca. 25--30, anthers of stamens and staminodia wider than long, <1 mm, filaments basally fused into a succulent torus, the free part of filaments <1 mm, much shorter than the fused part; stigmas rounded, pressed against the ovary. Fruits mostly globose, 2--3 cm, green to greenish red or red; seeds black with a white aril; calyx persistent in fruit. Distribution. Wet to very wet forest, from near sea level to 1600 m; central to eastern Panama. A distinctive, usually dark-reddish brown-drying, dull-leaved, Panamanian endemic (from the Canal to San Blas and Darién) with succulent and yellow rather than membranaceous white or pink petals, stamens with short, free filaments mounted on a succulent, fused basal portion, white rather than orange arils, and black rather than green seeds, not at all closely related to C. nicaraguensis, under which it was synonymized by D'Arcy (1980). This is the only species, described from the region, with succulent, rather than membranaceous petals. Even in fruit it can be identified by the combination of large, dark-drying leaves and fused staminodia. The petals dry black or very dark burgundy. Some of this material with particularly large inflorescences from far eastern Panama has been identified as C. floribunda Cuatrecasas and C. weberbaueri Engler However, the abundant material at hand suggests that probably only one species is involved. Pending study of type material of the older, South American names, I here use C. eclipes for all Panamanian collections. Representative specimens. PANAMA. Colon: end of Río Boquerón road, 500--650 m, Apr. 1978 (fr), Hammel 2445 (MO); Río Guanche, Feb. 1974 (fl), Foster 2780 (MO); Santa Rita Ridge Road, 450 m, Mar. 1975 (fr), Mori & Kallunki 5049 (MO). Canal Zone: Barro Colorado Island, Mar. 1969 (fr), Croat 8320 (MO); Pipeline Road, Sep. 1971, (fl), Gentry 1946 (MO). Panamá: Cerro Jefe region, 750 m, Jan. 1986 (fr), McPherson & Merello 8134 (MO); Gorgas Memorial Labs yellow fever research camp, 25 km NE of Cerro Azul on Río Piedras, 550 m, Nov. 1974 (fr), Mori & Kallunki 3371 (MO); Llano-Carti road, 350 m, Aug. 1986 (fl), McPherson 9962 (MO). San Blas: Cangandi, 30 m, Feb. 1986 (fr), de Nevers & Herrera 7186 (MO). Darién: Cana-Cuasi trail, Chepijana, 1600 m, Mar. 1940 (fr), Terry & Terry 1603 (MO); Parque Nacional del Darién, ridge between Río Topalisa and Río Pucuro, ca. 17 km E of Pucuro, 850 m, Oct. 1987 (fl), de Nevers et al.8380 (MO); slopes of Cerro Mali, 1300--1400 m, Oct. 1987 (fl & fr), Cuadros et al. 3927 (MO); S of El Real on trail up Cerro Pirre, 550--1030 m, Mar. 1985 (fr), McPherson 7045 (MO); trail to Cerro Pirre, 20--600 m, Aug. 1986 (fl), J. F. McDonagh et al. 483 (MO). Chrysochlamys glauca (Oersted, Planchon & Triana) Hemsley, Biol. Cent.-Amer., Bot. 1: 87. 1879. Tovomitopsis glauca Oersted, Planchon & Triana, in Planchon & Triana, Ann. Sci. Nat. Bot., Ser. 4, 14: 264. 1860. Tovomita glauca (Oersted, Planchon & Triana) L. O. Williams, Fieldiana Bot. 29: 357. 1961. TYPE: Costa Rica. [San José]: sur le mont Jaris, [ca. 850 m], Oersted 3 [3589] (holotype, C). Tovomita gracilis L. O. Williams, Fieldiana Bot. 29: 357. 1961. TYPE: Costa Rica. Alajuela: San Pedro de San Ramón, 1050 m, Brenes 4217 (lectotype, designated by Hammel (1986, F)). Tovomitopsis multiflora Standley, Field Mus. Nat. Hist. Bot. Ser. 4: 234. 1929. TYPE: Panama. Chiriquí: Progreso, Cooper & Slater 171 (holotype, F--573,086; isotype, NY). Shrubs or rarely trees 1--6(--14?) m; resin clear to slightly milky. Leaf blades elliptic, 7--20 X 2--7 cm, the apex and the base acute to acuminate, drying gray-green; lateral veins 0.5--1.5 cm apart, 6--10 pairs, fading towards the margin; petiole 1--2.5 cm. Inflorescence 2--12 cm, often pendulous, glabrous; flower buds ca. 3--4 mm, outer 2 sepals equal to markedly unequal, shorter than the inner ones; stamens 20--30, anthers of stamens and staminodia ovoid to ± globose, < 1 mm. Fruits pyriform to globose, 1--2 cm, greenish cream to nearly red, often two-toned; calyx deciduous in fruit. Distribution. Wet to very wet forest, from near sea level to 1400 m; Costa Rica and Panama. Even segregating the material described below as Chrysochlamys tenuis, C. glauca remains a rather variable species, characterized by its usually dull, few-veined leaves and small flower buds. Some of that variation may be worthy of taxonomic recognition. Many of the collections from the Osa peninsula in Costa Rica (cf. Aguilar 431, Kernan 713) have leaves with especially obvious and numerous latex canals and flower buds near the lower end of the size range for the species, giving such material an aspect predictive of its origin. Allen (1956: 345), in fact, distinguished two species from the Osa region within the C. glauca complex. For one (vouchered by Skutch 5286, F, not seen), he used the name C. costaricana, stating that it was "nearly identical" to C. glauca, "differing principally in the smaller flowers." Although C. costaricana (discussed below under C. psychotriifolia) is not its correct name, the species distinguished by Allen may deserve recognition, not since given. Populations at the other end of Costa Rica, in the Cordillera de Guanacaste, have almost shiny leaves with relatively obscure lateral veins. The plants also appear to flower more on lateral shoots than material from elsewhere. The anthers are slightly longer and the stigmas are larger than those of C. glauca from elsewhere, usually at least 1/2 (vs. no more than 1/3--1/4) the length of the ovary in newly opened flowers. Several collections from Panama, Panama province and San Blas (de Nevers 7223, Herrera et al. 1137, Herrera et al. 1145), tentatively placed here, have unusually dense and much-branched resin lines, and fruits with persistent sepals. These may represent a distinct species, but flowering material is needed. Representative specimens. Costa Rica. Guanacaste: Volcán Cacao, Rancho Harold, 700--1200 m, July 1986 (fr), I. & A. Chacón 2154 (CR); Parque Nacional Guanacaste, Estación Cacao, 1100 m, Nov. 1990 (fr), Chávez 368 (INB). Alajuela: Río Cataratas, San Ramón, 600 m, Jan. 1984 (fl), Gómez-Laurito 9700 (CR). Limón: Cantón de Pococí, Barra del Colorado, 15--20 m, Jan. 1995 (fl), Araya 713 (CR, INB, MO) Puntarenas: airport 4 mi. W of Rincón de Osa, 40 m, Aug. 1967 (fr), Raven 21564 (CR); wooded slopes above Golfito, 0--80 m, Jan. 1967 (fl), Burger & Mata 4775 (CR, F); Las Cruces, Jardín Botánico Wilson, 1100 m, Sep. 1993 (fr), G. Vargas et al. 1369 (CR). PANAMA. Bocas del Toro: Río Teribe, alrededores de la estación Teribe II del IRHE, Oct. 1984, Carrasquilla 2026 (MO). Chiriquí: E of Cañas Gordas near Costa Rican border on road to Volcán, 800--1200 m, Feb. 1973 (fl, fr), Liesner 268 (MO); road from Nueva California to Río Serano, 1400 m, Apr. 1979 (fl), D'Arcy et al. 13057 (MO). Panamá: Llano-Cartí road, 400 m, Jan. 1974 (fl), Nee & Dressler 9371 (MO). San Blas: Cordillera de San Blas, cabecera del Río Piriati, 460--799 m, Feb. 1992 (fr), Herrera et al. 1137 (MO). Authorship of the four Central American Tovomitopsis names published in Planchon and Triana (1860) has been variously obfuscated in numerous publications, including my own. Oersted alone never published these names, as sometimes implied (cf. Williams, 1959), nor is there any basis for the even more common "Oersted ex Planchon & Triana," (cf. Standley, 1937; Croat, 1978; D'Arcy, 1980), let alone the terse permutation of the latter to just "Planchon & Triana" (e.g., Hammel, 1986). Since Anders Örsted (1812--1872) was professor of botany at the University of Copenhagen until 1862 (http://www.nathimus.ku.dk/bot/orsted.htm) during the time Planchon and Triana were working in Paris on their Guttiferae memoir, nothing suggests other than the three, "Oersted, Planchon & Triana," as co-authors, as originally indicated for all four species in Planchon and Triana (1860). The citation of two collection (?) numbers for the types of these same four species results from examination of the actual specimens. The single-digit numbers are cited in the original publication (e.g., Oersted 4), and are also written on or in packets with the specimens. The four-digit numbers printed on the labels (e.g., Plantae Centroamericanae Oersted No. 3585) are not mentioned in the publication. Neither series appears to come from normal consecutive field-note numbers; earlier dates do not correspond, consistently, to lower numbers within the series. Chrysochlamys grandifolia (L. O. Williams) Hammel, comb. nov. Basionym: Tovomita grandifolia L. O. Williams, Trop. Woods 112: 96. 1960, new name for T. macrophylla L. O. Williams, Trop. Woods 111: 16. 1959, non T. macrophylla (Poeppig) Walpers 1842. TYPE: Costa Rica. Puntarenas: Esquinas forest, between Río Esquinas and Palmar Sur, 30 m, Allen 5968 (holotype, US; isotypes, EAP, F). Shrubs or trees 2--10 m; resin clear to yellowish. Leaf blades elliptic, 18--35 X 10--18 cm, the apex acute to rarely rounded, the base acute to acuminate, drying reddish; lateral veins 1--2 cm apart, 14--18 pairs, looping to form a ± distinct submarginal vein; petiole (2--)3--4 cm. Inflorescence ca. 6--10 cm, erect, glabrous; flower buds ca. 7 mm, outer 2 sepals ± equal to each other and shorter than the inner ones; stamens ca. 50, anthers of stamens and staminodia longer than wide, ca. 2 mm. Fruits ovoid, strongly 5- or 6-ribbed (star-shaped in cross section), greenish cream to nearly red, often two-colored; calyx persistent in fruit. Distribution. Very wet forest, 10--600(--1300) m; Costa Rica to central Panama. This distinctive Central American species, clearly delimited by its protologue, is characterized by strongly ribbed fruits and monomorphic stamens with anthers up to 2--3 times longer than wide. Costa Rican material from the Pacific slope has glabrous inflorescences and anthers nearly 2 mm long, whereas that of the Caribbean slope (also occurring at higher elevation than that of the Pacific) and the Panamanian collections have markedly puberulent inflorescences and smaller anthers. This species was originally published as Tovomita macrophylla L. O. Williams, a homonym of the earlier T. macrophylla (Poeppig) Walpers Williams, therefore, made a new name, T. grandifolia, for the same species. Coincidentally, an earlier Chrysochlamys macrophylla Pax would prevent the use of that combination for the present species, in any case. Nothing, however, prevents the use of the combination C. grandifolia (L. O. Williams) Hammel. Although D'Arcy (1980) created the combination Tovomitopsis myrcioides (Planchon & Triana) D'Arcy for this material (with Tovomita grandifolia as synonym), the South American C. myrcioides Planchon & Triana is a quite different and poorly known species. It has very short petioles and the dimorphic stamens of section Heterandra "externa fertilia, libera, interna sterilia in corpus centrale concreta" (Planchon & Triana, vol. 14: 260. 1860) with nearly globose anthers. Furthermore, I know of no such rib-fruited Chrysochlamys species from South America. Chrysochlamys membrillensis (see below) of far eastern Panama also has somewhat ribbed fruits, but is otherwise quite different from the present species. Representative specimens. Costa Rica. Cartago: Turrialba; 2 km al oeste de Grano de Oro, 1200 m, July 1995 (fl), Cascante 639 (CR). Limón: Cord. Talamanca, entre Cerro Chimú y Cerro Matama, 1200 m, Abr. 1985 (fl, fr), Gómez & Herrera 23536 (CR); fila entre cuenca superior del Río Xichiari y cuenca superior del Río Boyei, 1300 m, Ago. 1995 (fl), Herrera 8427 (CR). San José: ca. 20 km S of San Isidro de El General, El Pilar de Cajón, 620 m, Nov. 1988 (fr), Zamora et al. 1521 (CR, MO). Puntarenas: Cantón de Osa, Fila al Oeste de Rancho Quemado, 300 m, Aug. 1991 (fr), J. Marín 127 (CR, MO); Sierpe, cabeceras de Quebrada Guerra, 200 m, June 1990 (fl), Herrera 4240 (CR, INB, MO); Corcovado National Park, trail from Los Chiles to base of hills, 20--400 m, July 1977 (fl), Liesner 3175 (CR, MO); Los Planes (La Gloria), 100 m, Feb. 1991 (fr), R. González 11 (CR, F, INB, MO); Cantón de Golfito, Refugio Nacional Golfito, S end of Fila Gamba, 160--200 m, Dec. 1988 (fr), Grayum et al. 9180 (CR, MO). PANAMA. Bocas del Toro: road from Fortuna Dam to Chiriquí Grande, 400 m, Apr. 1986 (fl), McPherson 9020 (MO). Chiriquí: Camino hacia la finca Landau, NE del campamento de Fortuna (Hornito), sitio de presa, 1100 m, Aug. 1976 (fr), Correa et al. 2339 (MO). Veraguas: 5 mi. W of Santa Fe on road past Escuela Agrícola Alto Piedra, 800--1200 m, Mar. 1973 (fr), Croat 23151 (MO). Coclé: El Copé, Atlantic slope E of sawmill, 900 m, Feb. 1979 (fr), Hammel 6271 (MO). Colón: Santa Rita Ridge Road, 600 m, Feb. 1980 (fl), Antonio 3756 (MO). Panamá: between peaks on Cerro Trinidad, May 1968 (fl), Kirkbride & Duke 1647 (MO); Cerro Campana, near tower, Mar. 1977 (fr), Folsom et al. 2311 (MO). Chrysochlamys guatemaltecana Donnell-Smith, Bot Gaz. 13: 26. 1888. Tovomitopsis guatemaltecana (Donnell-Smith) Standley & Steyermark, Publ. Field Mus. Nat. Hist., Bot. Ser. 23: 66. 1944. TYPE: Guatemala. Alta Verapaz: von Tuerckheim 989 (holotype, US; isotype, NY). Trees or shrubs 3.5--15 m; resin clear to pinkish. Leaf blades elliptic 12--25 X 5--9(--10) cm, the apex acute, the base ± acuminate; drying tan to grayish tan and usually with rather obvious resin lines; lateral veins 1--2 cm apart, ca. 8--12 pairs, fading to the margin; petiole 1.5--2 cm. Inflorescence 7--15 cm, erect, glabrous; flower buds ca. 5 mm, the outer 2 sepals ± equal, < 1/2 the length of the bud; stamens ca. 50, anthers of stamens and staminodia about as long as wide, < 0.5 mm. Fruits pyriform, 2.5--4 cm, red tinged with green; calyx deciduous in fruit. Distribution. Wet forest; 100--1450 m. Mexico to Belize. This name has been considered a synonym of Chrysochlamys nicaraguensis (cf. Standley & Williams, 1961) and C. psychotriifolia (cf. Hammel, 1984). However, a clearer picture of variation within the genus throughout the region now supports its reinstatement. The species is characterized by more or less shiny, usually reddish-brown-drying leaves with rather few (6--9) and distant (1--2 cm) major lateral veins and usually quite prominent intersecondaries and obvious resin lines. The rather large flower buds (5--7 mm) have visible resin lines. It has more stamens (ca. 50) than either C. nicaraguensis (35--40) or C. psychotriifolia (ca. 20). The fruits are often quite elongate, up to 4 cm, as in C. psychrotiifolia, and the sepals are early deciduous. Representative specimens. MEXICO. Oaxaca: 18 km al este de Cuahutemoc, por la carretera Cuahtemoc-Sarabia, Dec. 1974 (fr), Vazquez et al. 1482 (MO); Mpio. Matías Romero, 8.3 km al S de Esmeralda, luego 2.3 km al E, 110 m, May 1981 (fl, fr), Wendt et al. 3267 (MO); Mpio. Sta. María Chimalpa, Río Verde, 360--430 m, July 1986 (fl), Caletti et al. 210 (MO); Mpio. Minatitlán, lomas al S del Poblado 11, ca. 30 km al E del Campamento La Laguna, 180 m, July 1980 (fl, fr), Wendt et al. 2642 (MO). GUATEMALA. Alta Verapaz: 7 mi. up road to Oxec, 700 m, July 1977 (fl), Croat 41631 (MO). Izabal: Puerto Mendez, km 7, Sep. 1970 (fl), Contreras 10237 (MO). BELIZE. Toledo: Central camp, Edwards Road beyond Columbia, June 1951 (fl), Gentle 7354 (MO); SW Maya Mountains, Columbia River Forest Reserve, 750 m, Apr. 1992 (fl), Holst 4407 (MO); vicinity of San José Mayan Indian village, June 1973 (fl, fr), Croat 24450 (MO). HONDURAS. Copán: Parque Nacional Cerro Azul, 1440 m, Feb. 1992 (fr), Hawkins & Mejía 184 (MO). Olancho: Refugio de Vida Silvestre La Muralla, 1400 m, June 1992 (fl), Mejía 164 (MO). Chrysochlamys membrillensis (D'Arcy) Hammel, comb. nov. Basionym: Tovomitopsis membrillensis D'Arcy, Ann. Missouri Bot. Gard. 67: 1036. 1980. TYPE: Panama. Darién: Upper Río Membrillo, 100--800 m, Duke 10872 (holotype, MO). Shrubs or trees to ca. 8 m; resin clear. Leaf blades narrowly elliptic, 16--25 X 4--6 cm, the apex acuminate, the base acute and slightly decurrent, drying reddish to grayish tan and with numerous, closely spaced, often-branched, resin lines; lateral veins 1--2.5 cm apart, ca. 10 pairs, fading towards the margin and very indistinctly loop-connected; petiole 1--3.5 cm. Inflorescence ca. 10 cm, erect, glabrous; flower buds ca. 5--6 mm, the outer 2 sepals ± equal and as long as or shorter than the others; stamens ca. 40, anthers of stamens and staminodia slightly longer than wide, ca. 1 mm. Fruits ovoid-pyriform, ca. 3 cm, strongly alate but the wings thin, pink; calyx ± persistent in fruit. Distribution. Very wet primary forest, 100--850 m. Known for sure from just five collections, only from Panama, low to middle elevations of Colón province and extreme eastern Darién province. This species must certainly also occur in nearby Colombia, but no such specimens have been seen. This Panamanian endemic has narrowly elliptic and often falcate, coriaceous leaves, with very visible resin canals. The lateral leaf veins are without prominent intersecondaries, and the fruits are basically pyriform and winged. Although the winged fruits are very characteristic of this species, they are not obvious in young pistillate material nor even more mature, dried material. Specimens examined. PANAMA. Colón: Santa Rita Ridge Road, 600--700 m, Apr. 1981 (fl), Sytsma et al. 4176 (MO); 250--400 m, May 1982 (fl), Knapp & Schmalzel 5264 (MO). Darién: Parque Nacional del Darién, between Río Topalisa y Río Pucuro, 750--850 m, Oct. 1987 (fl, fr), Hammel et al. 16282 (MO); Upper Río Membrillo, 100--800 m, Apr. 1967 (fr), Duke 10863 (MO); Apr. 1967 (fr), Duke 10901 (MO). Chrysochlamys nicaraguensis (Oersted, Planchon & Triana) Hemsley, Biol. Cent.-Amer., Bot. 1: 87. 1879. Tovomitopsis nicaraguensis Oersted, Planchon & Triana, in Planchon & Triana, Ann. Sci. Nat. Bot., ser. 4. 14: 266. 1860. Tovomita nicaraguensis (Oersted, Planchon & Triana) L. O. Williams, Trop. Woods 111: 18. 1959. TYPE: Nicaragua. [Río San Juan]: along the Río San Juan, [ca. 10--50 m], Oersted 2 [3587] (holotype, C). Chrysochlamys standleyana L. O. Williams, Trop. Woods 111: 16. 1959, new name for C. pauciflora Standley, Ceiba 3: 214. 1953 non C. pauciflora Steyermark 1952. Tovomitopsis standleyana (L. O. Williams) D'Arcy, Ann. Missouri Bot. Gard. 67: 1040. 1980. TYPE: Panama. Bocas del Toro: Chiriquí Lagoon, Wedel 1019 (holotype, F; isotype, MO). Shrubs or trees, 3--8 m; resin clear. Leaf blades elliptic to obovate-oblong, 14--25(--27) X 6--10(--12.5) cm, the apex acute to rounded and apiculate, the base acute to acuminate, drying gray or tannish green, more or less glossy; lateral veins 1.5--3 cm apart, (5--)6--8(--10) pairs, fading to the margin; petiole 1.5--4 cm. Inflorescence 3--19 cm, usually erect; flower buds 5--7 mm, outer pair of sepals ± equal to each other and shorter than the inner; stamens 35--40, anthers of stamens and staminodia about as wide as long, < 1 mm. Fruits obovoid to globose, 1.5--3 cm, greenish pink to red; calyx deciduous in fruit. Distribution. Very wet forest of the Caribbean lowlands, from near sea level to 600(--1200) m; Nicaragua, Costa Rica, Panama. Chrysochlamys nicaraguensis, often erroneously interpreted as a wide-ranging species (cf. discussion under C. eclipes), is still only known for certain from the very wet Caribbean lowlands of Nicaragua, Costa Rica, and adjacent Panama. It is distinctive for its very glossy and distantly veined (2--3 cm) leaves almost totally lacking visible intersecondaries and resin canals. For discussion of authorship and type specimen citation of this species, see under C. glauca. Representative specimens. NICARAGUA. Río San Juan: Campamento La Lupe, ca. 15.5 km al Noreste del poblado El Castillo, 100 m, Dec. 1991 (fr), Grijalva 5633 (CR). COSTA RICA. Alajuela: Cantón de San Ramón, camino al Volcán Muerto, 1100--1200 m, May 1993 (fl), F. Araya et al. 294 (CR, INB, MO). Heredia: Cantón de Sarapiquí, La Selva Research Station, 100 m, Feb. 1981 (fr), Folsom 8775 (CR, DUKE). Limón: Cerro Coronel, 60--100 m, Mar. 1987 (fl), Stevens et al. 24851 (CR, MO); Parque Nacional Tortuguero, ca. 3 km al Sur del pueblo, 3 m, Nov. 1988 (fr), Robles 2175 (CR, MO); Cantón de Talamanca, Aguas arriba de Río Banano, Fila Asunción, 400 m, Feb. 1995 (fr), Rodríguez 580 (CR, INB, MO); hills between headwaters of Quebrada Mata de Limón and upper branches of Quebrada Tigre (Sixaola region), 25--30 m, Nov. 1984 (fr), Grayum et al. 4468 (CR, MO). PANAMA. Bocas del Toro: Fish Creek Mts., vicinity of Chiriquí Lagoon, Apr. 1941 (fl), von Wedel 2300(MO); Peninsula Valiente, on E side of Lagoon of Chiriquí, 25 m, Aug. 1987 (fl), McPherson 11453 (MO). Veraguas: valley of Río Dos Bocas along road between Escuela Agrícola Alto Piedra and Calovebora, 450--550 m, Aug 31 (fl), Croat 27581 (MO). Coclé: Alto Calvario, N of El Copé, 700--900 m, May 1977 (fr), Folsom 3274 (MO); region N of El Valle, 1000 m, Sep. 1946 (fl), Allen 3683 (MO). Panamá: Cerro Campana, 800 m, Aug. 1982 (fl), Hamilton & D'Arcy 662 (MO); Finca El Indio at headwaters of Río Indio, slopes of Cerro Jefe, 750 m, Nov. 1979 (fl), Antonio 2425 (MO). Colón: ca. 2--3 m up Río Guanche, 10--20 m, Jan. 1973 (fr), Kennedy & Foster 2193 (MO). San Blas: Cerro Habú, 500--850 m, Dec. 1980 (fr), Sytsma et al. 2676 (MO). Chrysochlamys psychotriifolia (Oersted, Planchon & Triana) Hemsley, Biol. Cent.-Amer., Bot. 1: 87. 1879. Tovomitopsis psychotriifolia Oersted, Planchon & Triana, in Planchon & Triana, Ann. Sci. Nat. Bot., Ser. 4. 14: 263. 1860. TYPE: Costa Rica. [Cartago]: Turrialba, [ca. 650 m], Oersted 4 [3585] (holotype, C). Tovomitopsis costaricana Oersted, Planchon & Triana, in Planchon & Triana, Ann. Sci. Nat. Bot., Ser. 4. 14: 265. 1860. Chrysochlamys costaricana (Oersted, Planchon & Triana) Hemsley, Biol. Cent.-Amer., Bot. 1: 87. 1879. TYPE: Costa Rica. [Cartago]: Turrialba [ca. 650 m], Oersted 1 [3588] (holotype, C). Tovomitopsis faucis D'Arcy, Ann. Missouri Bot. Gard. 67: 1031. 1980. Syn. nov. TYPE: Panama. Chiriquí: forests around El Boquete, 1000--1300 m, Pittier 3039 (holotype, US). Shrubs or trees, 3--10 m; resin clear or slightly creamy. Leaf blades oblong-elliptic, 12--21 X 3--8(--9) cm, the apex acute, the base acute to acuminate, dull, drying gray-green; lateral veins mostly 1(--1.5) cm apart, 10--15 distinct pairs, fading to the margin or somewhat loop-connected towards the leaf apex; petiole 1--2 cm. Inflorescence 6--16 cm, erect, minutely pubescent or glabrous; flower buds ca. 5--7 mm, outer 2 sepals ± equal to each other and shorter than the inner; stamens 15--20, anthers of stamens and staminodia about as long as wide, < 1 mm; stigmas ± reflexed. Fruits oblong and often apiculate, 3--5.5 cm, greenish yellow-pink to red; calyx deciduous in fruit. Distribution. Very wet montane forest, 600--1500 m; Costa Rica and Panama. Chrysochlamys psychotriifolia, well known in Costa Rica, is a species with numerous, rather close, lateral veins, few (ca. 20) monomorphic stamens, and usually large (3--5 cm), obovate-mammillate fruits. Tovomitopsis costaricana, T. faucis, and what D'Arcy (1980) called T. membranacea (Planchon & Triana) D'Arcy were based on very scant material that is, nevertheless, now seen to fall easily within the variation of C. psychotriifolia. For discussion of authorship and type specimen citation of this species, see under C. glauca. The South American Chrysochlamys membranacea Planchon & Triana, by its type and numerous more recent collections is clearly a different species, belonging to Planchon and Triana's section Heterandra, with dimorphic stamens in the staminate flowers and the staminodia connate into a ring or collar in the pistillate. Representative specimens. COSTA RICA. Guanacaste: Parque Nacional Rincón de La Vieja, 950--1100 m, Nov. 1987 (fr), Herrera 1446 (CR); Cantón de Tilarán, 1--2 km W of Lago Cote, 800--900 m, Aug. 1993 (fl), Haber & Zuchowski 11626 (CR, MO). Puntarenas: Reserva Monteverde, Vert. Pacific, 1520--1580 m, Sep. 1976 (fl), Dryer 757 (CR). Alajuela: La Palma de San Ramón, 1250 m, Nov. 1925 (fl), Brenes 4600 (385) (CR). Heredia: Horquetas de Sarapiquí, Reserva Rara Avis, 700 m, July 1995 (fr), Martén 1008 (CR). San José: Braulio Carrillo National Park, vicinity of Estación Carrillo, 600 m, July 1985 (fr), Hammel & Trainer 14276 (CR, MO). Cartago: Cantón de Turrialba, Valle del Reventazón, Grano de Oro, 1100 m, Sep. 1993 (fl), Campos & Campos 185 (CR, INB, MO). PANAMA. Bocas del Toro: road to Chiriquí Grande N of Fortuna Dam, 650--700 m, June 1987 (fl), McPherson 11130 (MO). Chiriquí: in forest along road to La Fortuna, 1400 m, Feb. 1979 (fl), Hammel 6232 (MO); ridge road at Cerro Colorado, 1450--1750 m, Aug. 1977 (fl, fr), Folsom et al. 4832 (MO); Veraguas: above Santa Fe beyond Escuela Agrícola Interamericana, Apr. 1976 (fr), Croat 34216 (MO). Chrysochlamys silvicola (Hammel) Hammel, comb. nov. Basionyym: Tovomitopsis silvicola Hammel, Selbyana 9: 113. 1986. TYPE: Costa Rica. Heredia: La Selva Biological Station [ca. 100 m], Hammel 9404 (holotype, DUKE; isotypes, CR, F, MO). Shrubs or trees 3--6 m; resin white. Leaf blades oblong-elliptic (13--)16--22 X 4--7(--8) cm, the apex acute to acuminate, the base acute, dull both surfaces, drying reddish tan; major lateral veins 0.5--2 cm apart, ca. 10--16 pairs, difficult to distinguish from the intersecondary veins, fading to the margin where indistinctly loop-connected; petiole 1--2.5 cm. Inflorescence 5--10 cm, ± pendent, minutely pubescent or glabrous; flower buds ca. 4--5 mm, outer 2 sepals often markedly unequal with the larger one ± overtopping the bud; stamens 20--30, anthers of stamens and staminodia about as long as wide, < 1 mm; stigmas ± reflexed. Fruits ovoid to globose, 1.5--2.5 cm, red throughout or whitish at base; calyx ± persistent in fruit. Distribution. Very wet, primary lowland forest, Caribbean slope, from near sea level to 700(--900) m; Costa Rica and Panama. This species, particularly as to its type, can be characterized by its primary lateral veins and intersecondaries being almost equally salient. Nevertheless, a few collections from Costa Rica included here (Heredia, Cantón de Sarapiquí, Rara Avis, 400--700 m, Martén 948 (CR); G. Vargas 1650 (CR); O. Vargas 2 (CR); O. Vargas 4 (CR)) have fewer and less prominent intersecondaries. Material from Panama appears to grade imperceptibly into an extreme with the intersecondaries very obscure. All the material has dull, yellowish to reddish tan-drying leaves, the outer pair of sepals often very unequal, the petals usually without obvious resin canals, and stamens about 20--30. At least in Costa Rica, the resin of the twigs is bright white, rather than clear or tardily creamy as in most other species in Central America. For its reddish tan-drying leaves, this species sometimes resembles the more northern C. guatemaltecana, which has more stamens and shiny leaves with fewer, more distant veins. Representative specimens. COSTA RICA. Alajuela: Cantón de San Carlos, (near) Boca Tapada, Finca La Monalisa, 100 m, May 1995 (fl), Rodríguez & Estrada 702 (CR, INB, MO). Heredia: between Río Peje & Río Sardinalito, Caribbean slope of Volcán Barva, 700--750 m, Apr. 1986 (fr), Grayum 6695 (MO); Finca La Selva, Puerto Viejo de Sarapiquí (ca. 100 m), July 1979 (fl, fr), Grayum 1902 (CR, DUKE). San José: Parque Nacional Braulio Carrillo, Carillo, 600 m, May 1982 (fl), Gómez-Laurito 8440 (CR). Limón: Ref. Nac. Barra del Colorado, forests and pastures between Chirripocito and Río Sardina, 12 m, Apr. 1990 (fr), Grayum 9794 (CR, MO, F); La Aurora, Guácima, ca. 5 km al E del pueblo, 100 m, Aug. 1988 (fl), Robles et al. 2087 (CR, MO, F); Cantón de Talamanca, Alto Urén, 700--900 m, Sep. 1994 (fr), Gallardo & Lépiz 313 (CR, INB, MO, F). PANAMA. Veraguas: NW of Santa Fe, 1 km from Escuela Agrícola Alto de Piedra, Feb. 1975 (fl), Mori & Kallunki 4794 (MO), 900--1200 m, May 1981 (fr), Sytsma & Andersson 4640 (MO). Coclé: El Copé, 1200 m, June 1979 (fr), Antonio 1189 (MO); N of Penonomé on road to Coclesito, 100--170 m, Feb. 1978 (fl), Hammel 1681 (MO). Panamá: between Cerro Azul and Cerro Jefe, July 1969 (fl), Foster 1157 (MO); El Llano-Cartí Road, 400--450 m, Jan. 1974 (fl), Nee & Dressler 9352 (MO), 350--400 m, Feb. 1973 (fr), Kennedy 2543 (MO). San Blas: Río Diablo y vecindad de Duque Sui, 80--110 m, July 1992 (fl), Herrera et al. 1174 (MO). Darién: vicinity of upper gold mining camp of Tyler Kittredge on headwaters of Río Tuquesa, Aug. 1974 (fr), Croat 27209 (MO). Chrysochlamys skutchii Hammel, sp. nov. TYPE: Costa Rica. San José: basin of El General, 675--900 m, Mar. 1940 (fl), Alexander Skutch 4759 (holotype, CR; isotype, MO). Figure 1. 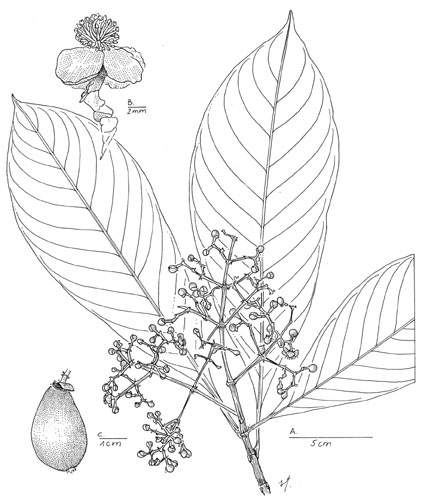 Figure 1. Chrysochlamys skutchii Hammel (A, B, Skutch 4759; C, Aguilar 1771). --A. Flowering branch. --B. Staminate flower. --C. Fruit. Altitudo altior C. allenii affinis a qua differt foliis majoribus, tenuioribus, par extimus sepala inaequalia, calyx ad maturiatem fructus persistens et inflorescentia glabra. Trees or shrubs 2--10 m; resin clear to creamy (red?). Leaf blades elliptic, (17--)21--35 X 6--12 cm, membranaceous, the apex and base acute to acuminate; lateral veins ca. 1--2 cm apart, ca. 10--17 pairs, mostly reaching to the margin or the apical veins looping into a ± distinct, submarginal vein; petiole 2--4 cm. Inflorescence 7--15 cm, erect, glabrous; flower buds ca. 5 mm, outer pair of sepals very unequal, one < 1/2 the length of the bud, the other ± equal to the bud; petals membranaceous, lacking distinctly contrasting resin canals; stamens ca. 50, varying in length centripetally, 1--4 mm, the anthers nearly as wide as long, ca. 0.3 X 0.4 mm; stigmas sessile, spreading or slightly elevated. Fruits ± pyriform or obovate, 2.5--4 cm, pale green-pink to red; seeds ca. 1 cm, the aril orange; calyx ± persistent in fruit. Distribution. This species is known only from a few collections from relatively low 100--750(--900) m elevations in wet forest on the Pacific slope of southern (and eastern) Costa Rica, from the regions of Acosta (Fila Bustamante), Valle del General, the Osa Peninsula and vicinity. Flowering or sterile collections of this species may be difficult to separate from Chrysochlamys grandifolia, a species most readily distinguished by its strongly ribbed fruits. The two grow sypatrically and have been confused on Costa Rica's Osa Peninsula. However, the leaves of C. grandifolia usually have a more distinct submarginal vein, it has fewer stamens, and the anthers are often 2--3 times longer than wide rather than about as long as wide. In many details, e.g., fruit shape and color, number and size of stamens, leaf coloration, C. skutchii comes much closer to C. allenii. Chrysochlamys skutchii is distinguished from C. allenii by its larger, thinner leaves, glabrous inflorescence, unequal outer sepal pair, persistent sepals in fruit, and lower elevation habitat. The epithet is in honor of the species' first collector, one of Costa Rica's primier natural historians, Alexander Skutch. Additional specimens. COSTA RICA. Puntarenas: Cantón de Golfito, La Gamba, "Bosque de los Austriacos," 300 m, Dec. 1993 (fl), Huber & Weissenhofer 68 (MO), (fl), Huber & Weissenhofer 90 (CR), (st.), Huber & Weissenhofer 94 (CR); Cantón de Osa, Parque Nacional Corcovado, Cerro Brujo, 600 m, Jan. 1991 (fl), Castro 268 (MO); Cerro Rincón, 745 m, Aug. 1990 (fr), Morales et al. 20 (CR, MO); Cerro Mueller, 150--650 m, May 1988 (fr), Kernan & Phillips 474 (CR, MO); Estación Los Patos, 250 m, May 1995 (fr), Moraga 191 (CR, MO), 500 m, July 1988 (fr), Kernan & Phillips 662 (CR); Los Planes, La Gloria, 170 m, Feb. 1991 (fr), Induni 266 (CR, INB, MO), 100 m, Feb. 1991 (fr), R. González 32 (CR); Rincón, Fila Casa Loma, 50--400 m, July 1990 (fr), Chacón 941 (CR, MO); Camino de Altura, 100--300 m, Aug. 1967 (fr), Raven 21512 (MO); Aguabuena, 3.5 km W of Rincón, 450 m, Nov. 1992 (fl), Thomsen 188 (CR); Rancho Quemado, 500 m, Nov. 1991 (fl), Marín et al. 256 (MO), 300--400 m, May 1988 (fr), Hammel et al. 16898 (CR, MO), 200--350 m, Apr. 1993 (fr), Aguilar 1771 (CR, INB, MO). Chrysochlamys tenuifolia Cuatrecasas, Revista Acad. Colomb. Ci. Exact. 29: 59. 1950. TYPE: Ecuador. Río San Miguel o Sucumbíos, 250 m, Cuatrecasas 10880 (holotype, F; isotype, COL). Shrubs or small trees to 2.5--7 m, most twigs with paired bud-scale scars just above the node; resin yellow to orange. Leaf blades elliptic, 8--14(--24) X 3--4.5(--9) cm, the apex acuminate, the base acute to obtuse, drying gray-green; major lateral veins ca. 3(--20) mm apart, ca. (12--)16 prominent pairs but with several nearly equally prominent intersecondaries, fading to the margin or looping to an obscure submarginal vein; petiole 0.6--1.2 cm. Inflorescence appearing axillary or terminal, 1--3 cm, erect, glabrous; flower buds 2--3 mm, the outer 2 sepals 1/3 the length of the bud; stamens ca. 60, apparently without filaments, anthers of stamens and staminodia broader than long, < 0.5 mm, surmounting a resiniferous capitulum . Fruits ellipsoid-falcate, bean-shaped, red, 1.5--2.5 cm, 3-valved but usually just 1-seeded, the seed black or brown with an orange aril; sepals persistent in fruit. Distribution. Very wet forest; 300--1400 m. Eastern Panama, Colombia, Ecuador. Because of its apparently axillary inflorescences, bud-scale scars, and resin-producing staminate flowers, this species's position in Chrysochlamys is somewhat dubious. It has many close and very prominent lateral veins, and small, essentially terminal inflorescences that are often left in an apparently axillary position (only one per node) by the dominant axillary vegetative shoot. It also sometimes has unambiguously terminal inflorescences, and has an androecium with the stamens more or less connivent into an apparently resiniferous capitulum, much as in the resin-producing flowers of some species of Clusia. The staminodia of the pistillate flowers are connate into a ring or collar around the base of the ovary as in Chrysochlamys membranacea. The small, narrowly ellipsoid-falcate, capsulate fruits mostly have only three sessile stigmas, and usually one seed with a markedly cellular aril. In leaf venation and bud scales it is very similar to Symphonia, but for numerous reasons would be even more out of place there than in Chrysochlamys. The species must certainly belong to the Tovomita group, but further study is in order. Although D'Arcy (1980) created the combination Tovomitopsis micrantha (Engler) D'Arcy for this material, the Peruvian Chrysochlamys micrantha Engler, by its original description and type photo, bears it no resemblance. Representative specimens. PANAMA. San Blas: forest SE of Puerto Obaldia, Aug. 1971 (fr), Croat 16754 (MO); mountains above Puerto Obaldia, Aug. 1971 (fr), Gentry 1487 (MO). Darién: Cerro Mali on Colombian border, 1400--1500 m, Jan. 1975 (fr), Gentry & Mori 13806 (MO); Parque Nacional del Darién, ridge between Río Topalisa and Río Pucuro, ca. 17 km E of Pucuro, 600-850 m, Oct. 1987 (fr), de Nevers et al. 8343 (MO), (fl), Hammel et al. 16379 (MO), 700--1400 m, Oct. 1987 (fl, fr), Hammel et al. 16357 (MO); Río Tacarcuna, vicinity of old Tacarcuna village, 580 m, Jan. 1975 (fl), Gentry & Mori 13586 (MO). Chrysochlamys tenuis Hammel, sp. nov. TYPE: Costa Rica. Limón: Cordillera de Talamanca, Fila Matama, 09°47'18"N, 83°08'45"W, 1350 m, Apr. 1989 (staminate fl), G. Herrera & A. Chacón 2673 (holotype, CR; isotypes, MO + 2). Figure 2.  Figure 2. Chrysochlamys tenuis Hammel (A--C, Herrera & Chacón 2673, D, Berrocal et al. 74). A. --Flowering branch. --B. Flower bud. --C. Staminate flower. --D. Fruit. Chrysochlamys glauca affinis a qua differet foliis angustioribus et plerumque in sicco ferruginis vice glaucorum, venatione magis inconspicuo, inflorescentia tenuiore. Shrubs or small trees 2--5 m; resin clear to tardily creamy. Petiole 0.5--2 cm. Leaf blades narrowly elliptic, 5--13 X 1--2.5(--3.5) cm, the apex acuminate, the base narrowly acute and shortly decurrent, both surfaces dull, drying reddish tan to gray-green; lateral veins ca. 0.5 cm apart, 6--8 pairs, very indistinct, often difficult to distinguish from intersecondaries, fading to the margin or looping to form an obscure submarginal vein; resin canals usually very obscure, ± parallel to the midrib or at least somewhere crossing the lateral veins. Inflorescence paniculate, very lax to slightly congested, ± globose, ca. 2--5(--8) cm long, 3- to ca. 30-flowered, the rachis very slender, often deflexed or pendulous, glabrous; flower buds 3--4 mm long, usually conspicuously striate with resin canals; outer 2 sepals ± equal and shorter than the bud, sometimes very narrowly triangular, rarely (in the type and one nearby paratype, Herrera 2495) the third and fourth sepals long-acuminate with the filiform tips overtopping the rest of the bud by ca. 2 mm; petals membranaceous; stamens ca. 20, free, 2--2.5 mm long, the anthers ± globose, ca. 0.3 mm long; stigmas 5. Fruits narrowly obvoid to pyriform, usually rostrate, 2--3 cm long reddish, often tinged with white or pale yellow, 1--3-seeded; seeds with an orange aril; calyx deciduous in fruit. Distribution. This species is known from scattered localities in very wet forest on the Atlantic slope of Costa Rica and Panama, and from a few collections from wet forest on the Burica Peninsula, shared by both countries along their Pacific border. Although most material is from below 800 m, the type is from nearly 1400 m. The type and all other Costa Rican, Atlantic slope collections, as well as one Panamanian collection from Bocas del Toro (Kirkbride & Duke 621), one from Veraguas (McPherson 10724), and two from the Río Blanco de Norte area of the Atlantic lowlands of Coclé, have particularly long, narrow leaves without obvious intersecondaries and relatively few-flowered, very lax inflorescences with the outer sepals slightly to very narrowly triangular. The Burica Peninsula outliers and all other collections from Panama tend to have somewhat broader and shorter leaves with more obvious intersecondaries, multi-flowered, more congested inflorescences, and broader outer sepals. Chrysochlamys tenuis is similar to C. glauca in its dull, few-veined leaves and small flower buds. The Río Guanche and Cerro Campana populations cited here were discussed by D'Arcy (1980, p. 1034) under that species as "suggestive of a distinct variety or even species." Rather than further blurring what then seemed only suggestive, more recent collections, particularly those from Atlantic Costa Rica, strengthen the case for recognizing this entity as a distinct species. Other than the narrower and usually reddish tan vs. gray-green-drying leaves, more obscure venation and smaller, more delicate inflorescences, C. tenuis is distinguished from C. glauca by the resin canals of the leaves, which, when visible, are more or less parallel to the midrib or at least somewhere crossing the lateral veins, rather than parallel to the lateral veins, as in C. glauca and most other species. The narrowly obovoid fruits of C. tenuis are usually only 1- or 2-seeded, whereas the more or less globose or broadly obovoid fruits of C. glauca are normally 5-seeded. Mori et al. 3950 recorded the seeds as "steel blue." The new species also superficially resembles the Panamanian C. angustifolia, which, however, is an epipetric rheophyte with milky resin, stouter inflorescence axes, and much larger flower buds. Original field notes for the type of this species indicated four duplicates. These were earlier distributed as either Tovomitopsis angustifolia or T. glauca, and their whereabouts, except as indicated above, are uncertain. Additional specimens. COSTA RICA. Limón: Cordillera de Talamanca, Fila Matama, Cerro Muchilla, 850 m, Apr. 1989 (fr), Robles & Chacón 2678 (CR, MO); Reserva Biol. Hitoy Cerere, Cerro Bitárkara, 800 m, Feb. 1989 (fl bud), Herrera & Solís 2495 (CR, MO); Río Tercero, 600 m, May 1985 (fr), Berrocal et al. 74 (CR); Suretka, 200 m, July 1982 (fr), Gómez-Laurito 8839 (CR). Puntarenas: Burica Peninsula, Punta Burica, M. M. Chavarría 306 et al. (CR, INB). PANAMA. Chiriquí: Burica Peninsula, 8--11 mi. W of Puerto Armuelles, 200--500 m, Feb. 1973 (fl, fr), Croat 21995 (MO), Mar. 1973 (fl, fr), Croat 22472 (MO); La Fortuna hydroelectric project, 1200 m, Mar. 1978 (fl, fr), Hammel 2181 (MO). Bocas del Toro: Quebrada Huron on Cerro Bonyic, 170--400 m, Apr. 1968 (fr), Kirkbride & Duke 621 (MO); Chiriquicito-Calderas trail, Apr. 1968 (fr), Kirkbride & Duke 765 (MO). Veraguas: Escuela Agrícola Alto de Piedra, road to Calovébora, 450--550 m, Aug. 1974 (fr), Croat 27665 (MO), Nov. 1974 (fr), Mori & Kallunki 3224 (MO), Dec. 1974 (fr), Mori et al. 3950 (MO); slopes of Cerro Tute, 600--750 m, Mar. 1987 (fl), McPherson 10724 (MO). Coclé: above El Copé, ca. 700--900 m, Apr. 1977 (fr), Folsom & Robinson 2371 (MO), Apr. 1977 (fr), Folsom & Kauke 2652 (MO), Oct. 1977 (fr), Folsom et al. 5707 (MO), Nov. 1977 (fr), Folsom & Collins 6516 (MO), Jan. 1978 (fr), Hammel 953 (MO), Apr. 1978 (fr), Hammel 2432 (MO), June 1978 (fr), Hammel 3587 (MO), Jan. 1989 (fr), McPherson 13627 (MO); Atlantic slope from El Copé to Río Blanco del Norte, ca. 150 m, Feb. 1980 (fl, fr), Antonio 3653 (MO), 350 m, Feb. 1982 (fl), Knapp 3675 (MO); near El Valle, ca. 700--1000 m, Aug. 1967 (fr), Duke & Dwyer 13936 (MO), Jan. 1972 (fr), Gentry & Dwyer 3654 (MO), Dec. 1972 (fr), Gentry 6848 (MO), Sep. 1978 (fr), Hammel 4794 (MO), May 1967 (fl), Lewis et al. 1769 (MO), 1785 (MO), Nov. 1974 (fr), Mori & Kallunki 2960 (MO), June 1975 (fl), Mori et al. 6565 (MO). Panamá: Cerro Campana, ca. 900 m, May 1971 (fl), Croat 14681 (MO), 14753 (MO), Sep. 1982 (fr), D'Arcy 15071 (MO), Mar. 1969 (fl), Porter et al. 4259 (MO); Cerro Trinidad, 800-1000 m, Oct. 1946 (fr), Allen 3773 (MO); El Llano-Cartí Road, Feb. 1977 (fl, fr), Foster 2770 (MO). Colón: Santa Rita Ridge, Jan. 1968 (fl), Dwyer 8564 (MO); Río Escandaloso near Cerro Bruja, 10--200 m, Apr. 1978 (fl), Hammel 2672 (MO), 2713 (MO), Mar. 1982 (fl), Knapp & Kress 4283 (MO), (fr), Knapp et al. 4461 (MO); Río Guanche near Portobelo, ca. 0--100 m, May 1984 (fl, fr), Churchill 5119 (MO), Oct. 1978 (fr), Dec. 1973 (fl), Gentry 8764 (MO), Oct. 1978 (fr), Hammel et al. 4932 (MO), July 1971 (fl), Kennedy 1174 (MO), Jan. 1973 (fr), Kennedy & Foster 2132 (MO), Aug. 1981 (fr), Knapp 1024 (MO), Mar. 1975 (fr), Mori & Kallunki 5194 (MO), Mar. 1975 (fl), 5212 (MO); June 1975 (fl), Mori et al. 6443 (MO), Aug. 1975 (fr), Mori & Witherspoon 7955 (MO), Dec. 1973 (fr), Nee & Gentry 8719 (MO), July 1976 (fr), Sullivan 155 (MO), Oct. 1980 (fr), Sytsma 1645 (MO). Although the strictly South American Balboa does not occur within the geographic focus of this paper, taxonomic clarity and logical consistency prescribe its mention here as an additional synonym of Chrysochlamys. Planchon and Triana described the genus and its single species, B. membranaceae based only on staminate material and placed it in their subtribe "Euclusieae" rather than Tovomiteae. Examination of type material and more recent collections, of both staminate and pistillate plants (e.g., Boyle & Boyle 3303 (MO), Croat 73004 (MO), Gentry et al. 14655 (MO), Hammel & Trainer 15831 (MO), Hoover 1287 (MO), Hoover et al. 3136 (MO), Romero-Castañeda 2739 (COL), Rubio & Quelal 1339 (MO)) support the view that B. membranacea is a species of Chrysochlamys: Chrysochlamys balboa Hammel, nomen nov. Basionym: Balboa membranacea Planchon & Triana, Ann. Sci. Nat. Bot., ser. 4. 14:252. 1860, non C. membranacea Planchon & Triana, 1860. TYPE: Colombia. Between Tuquerres and Barbacoa, 1176 m, Apr. 1853, Triana s.n. (holotype, P; isotype COL) Chrysochlamys clusiifolia subsp. panamensis Maguire, Phytologia 36: 395. 1977. TYPE: Panama. Darién: Cerro Mai, Serranía del Darién, Gentry & Mori 13769 (holotype, MO) = Dystovomita paniculata (J. D. Smith) Hammel, Ann. Missouri Bot. Gard. 76: 927. 1989. D'Arcy (1980) already showed this to belong in his newly elevated genus, Dystovomita. Tovomitopsis membranifolia (Standley) D'Arcy, Ann. Missouri Bot. Gard. 67: 1035. 1980. Tovomita membranifolia Standley, Field Mus. Nat. Hist. Bot. Ser. 22: 160. 1940. TYPE: Panama. Darién: Caná-Cuasí trail, Chepigana District, Terry & Terry 1478 (holotype, F; isotype, MO) = Tovomita stylosa Hemsley, Biol. Cent.-amer. 1: 88. 1879. Hammel (1986) already showed this to be a synonym of Tovomita stylosa. Tovomita croatii Maguire, Phytologia 36: 404. 1977. Tovomitopsis croatii (Maguire) D'Arcy, Ann. Missouri Bot. Gard. 67: 1031. 1980. TYPE: Panama. Veraguas: road to Calovébora Croat 14268A (holotype, MO). Shrubs 1--2 m, resin clear. Leaf blades obovate, widest above the middle, 20--60 X 7--25 cm, the apex acuminate, the base narrowing to an acute or cordulate base; lateral veins ca. 1--2 cm apart, ca. 16--22 pairs, looping to form a distinct submarginal vein 2--5 mm from the margin; petiole lacking. Inflorescence 10--26 cm, usually pendulous, glabrous or puberulent; flower buds ca. 6 mm, the outer sepals often overtopping the inner; stamens 25--30, anthers of both stamens and staminodia about twice as long as wide, ca. 1.5 mm; stigmas concave and spreading. Fruits pyriform, green to pink, ca. 2 cm; seeds ca. 1 cm; calyx ± persistent in fruit. Distribution. Very wet, mostly lowland forest, 30--350(--1500) m; Costa Rica, Panama, Ecuador. This rare, somewhat problematic species might appear to walk the generic borderline between Chrysochlamys and Tovomita. The outer pair of sepals, although thinly membranaceous, exceed the bud as in Tovomita, yet styles are lacking and the fruits apparently do not have the dark maroon placenta and inner fruit wall typical of Tovomita. Nevertheless, two Costa Rican collections (Grayum 3374, Grayum & Schatz 5276) clearly indicate the presence of side shoots with clustered leaves. With present knowledge, placement of this species in Tovomita, particularly because of its branching pattern, is more justifiable than placement in Chrysochlamys. As a species, it is very distinctive, not only for the nearly sessile and sometimes very large leaves, but also for the numerous (16--22) lateral veins that are loop-connected to a distinct submarginal vein. Probably because of the species' rarity, its short stature and large leaves, the typical Tovomita branching pattern of T. croatii has gone unobserved and unrecorded. Representative specimens. COSTA RICA. Limón:
Reserva Biol. Hitoy Cerere, 90--200 m, July 1985 (fr),
Grayum & Hammel 5763 (CR, M0);
Cantón de Talamanca, Sukut, 350 m, July 1989 (fr),
Herrera 3168 (CR, MO); Finca Anai, W of Mata de
Limón (Sixaola region), 30 m, May 1985 (fl), Grayum
& Schatz 5276 (CR, MO). PANAMA.
Chiriquí: Fortuna Dam Area, 1100 m, May 1984 (fl),
Chuchill 5243 (MO). Coclé: El Valle,
Cerro Gaital, 900--1000 m, July 1968 (fl, fr), Dwyer
& Correa 8913 (MO). ECUADOR.
Pichincha: Quito a Sto. Domingo de los Colorados, km 94,
1300--1500 m, Dec. 1986 (fr), Zak 1529 (MO).
|
{kind=link}
{kind=link}
© 1995-2025 Missouri Botanical Garden, All Rights Reserved
4344 Shaw Blvd.
St. Louis, MO 63110
(314) 577-5100
Technical Support