QUICK SEARCH
MO PROJECTS:
Africa
Asia/Pacific
Mesoamerica
North America
South America
General Taxonomy
Photo Essays
Training in Latin
America
MO RESEARCH:
Wm. L. Brown Center
Bryology
GIS
Graduate Studies
Research Experiences
for Undergraduates
Imaging Lab
Library
MBG Press
Publications
Climate Change
Catalog Fossil Plants
MO DATABASES:
W³MOST
Image Index
Rare Books
Angiosperm
Phylogeny
Res Botanica
All Databases
INFORMATION:
What's New?
People at MO
Visitor's Guide
Herbarium
Jobs & Fellowships
Symposium
Research Links
Site Map
Search
Three new species of Celastraceae from Costa Rica, one disjunct from MexicoBarry E. Hammel
This paper was reviewed and accepted in November of 1996 for publication in Novon, Vol. 7 (2). In preparation for and in keeping with the theme of electronic distribution of the
Manual to the Plants of Costa Rica, the mss. was made available on two Web sites (INBio and MO) by 14 April 1997. Abstract. Three species of Celastraceae, each represented by many collections from Costa Rica, are compared to near relatives and found to be new: Gymnosporia haberiana Hammel, disjunct from Mexico and Costa Rica, Maytenus recondita Hammel, of Costa Rica and Panama, and Crossopetalum enervium Hammel, of Costa Rica . The South American Gymnosporia magnifolia (Loesener) Lundell is here confirmed as a synonym of G. urbaniana (Loesener) Liesner, and Crossopetalum eucymosum (Loesener & Pitter) Lundell is placed in synonymy under C. parviflorum (Hemsley) Lundell. Keys are provided for all accepted neotropical species of Gymnosporia, and for all Costa Rican species of Crossopetalum and Maytenus. For nearly twenty years collections of two large, cloud forest trees in the family Celastraceae have been accumulating, primarily from the Monteverde region of Costa Rica. Much of this material has been distributed to various herbaria under tentative or approximate names. More recent exploration along the central cordillera has extended the range, within Costa Rica, of these two species and yielded many collections of a third, shrubby species in the genus Crossopetalum, also known from the Monteverde region. The two trees are known almost exclusively from mid-elevations on the Pacific slopes of the northern mountains, while the shrub extends nearly the entire length of the country and is known from numerous collections on both slopes. Although one of the trees has always been known to be a Maytenus presumably related to M. schippii Lundell, generic placement of the other has been more problematic. This species, while having alternate leaves and capsulate fruits with arillate seeds, as in Maytenus, has greener drying, entire leaves, more open and branched inflorescences with 4-merous flowers, and several seeded, 2--4 lobed fruits. In these respects the Costa Rican (and Mexican) tree can be distinguished from other alternate-leaved Celastraceae and coincides perfectly with two South American species recently described in or transferred to the genus Gymnosporia (Lundell, 1985; Liesner, 1993). Gymnosporia haberiana Hammel, sp. nov. TYPE: COSTA RICA. Guanacaste: Cordillera de Tilarán, La Cruz de Abangares, 1400 m, 15 Jul 1985 (fl.) Haber & Bello 2034 (holotype INB; isotypes, BM, C, CAS, COL, CR, DUKE, F, FLAS, GH, HNMN, K, MEXU, MICH, MO, NY, P, PMA, TEX, US, USJ, W, WIS). Figure 1. A G. gentryi foliorum nervis lateralibus minus prominentis et latiore divergentibus, ovario et capsula tantum lobato non anguloso, a G. urbaniana floribus majoribus, disco manifesto, ab ambobus sepalis latioribus quam longioribus.
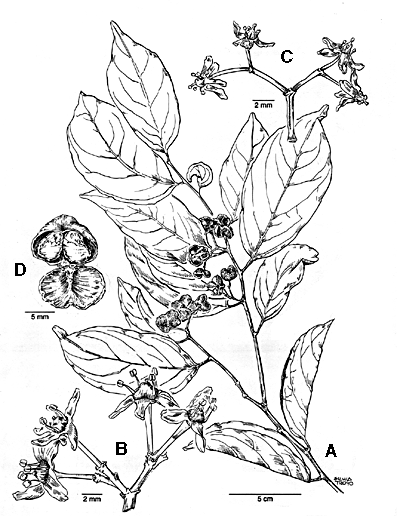
Figure 1. Gymnosporia haberiana. A. fruiting shoot; B. staminate flowers; C. pistillate flowers; D. fruit. (A, D from Haber & Zuchowski 11048; B from Hammel 17661; C from Haber 9303). Dioecious trees (3--) 5--30 m, the inner bark bright yellow; twigs terete; leaves and twigs drying pale to yellowish green, glabrous. Leaves alternate, 7-12 (--14) x 3--6 (--8) cm, elliptic, the apex acute to acuminate, the base acute to rounded, the margin entire and often curled under, all leaves often folded and curved along the midrib; main lateral veins mostly 4 or 5; petioles 5-10 mm; stipules small, deciduous. Inflorescences 2 or 3-branched, ca. 10--20-flowered axillary cymes, the rachis minutely and sparsely farinose-puberulent, the pedicels 2-4 mm. Flowers 4-merous, ca 4.5--7 mm wide, greenish; sepals ca. 0.4 x 1 mm, broadly rounded; petals imbricate, ca. 0.4 x 1 mm, spatulate or ligulate ca. 0.4 x 1 mm; stamens exceeding the style in staminate (pollen-bearing) plants, shorter than the style and without pollen in pistillate plants, anthers ± cordate, versatile; ovary ± globose, confluent with the conically raised disk, 2-celled with 2 ovules/cell. Fruits ca. 1 x 1--1.5 cm, globose or often wider than long and 2--4 lobed, (1) 2--4-seeded; seeds dark brown with white aril. Additional specimens examined. COSTA RICA. Alajuela: Cordillera de Guanacaste, P. N. Rincón de la Vieja, 10°47'50"N 85°18'19"W, 1500 m, 6 Jul 1991 (fl.), Rivera 1423 (CR, F, INB, MICH, MO, USJ, W). Guanacaste: Cordillera de Guanacaste, P. N. Guanacaste, Volcán Cacao, ca. 10°55'45"N 85°28'15"W, 1100-1400 m, 25 Nov 1989 (fr.), Chávez 7 (CR, F, MO); Dec 1990 (fr.), 478 (CR, F, INB, MO); Jul 1989 (fl.), Hammel 17661 (BM, CAS, COL, CR, F, INB, MEXU, MO, TEX, US); 9 Feb 1995 (fr.), Quirós 48 (CR, INB, MO); P. N. Rincón de La Vieja, sendero al Volcán, 10°45'50"N 85°18'50"W, 820 m, Jan 1991 (fr.), Rivera 974 (CAS, CR, F, INB, MO). Puntarenas: Cordillera de Tilarán, Monteverde Reserve and vicinity, ca. 10°20'N 84°50'W, 1200--1550 m, 4 Mar 1990 (fr.), Bello 2110 (F, INB, MO, MV); 28 Jul 1977 (fl.), Dryer 1589 (CR, MO); 4 Apr 1985 (fr.), Haber & Bello 1458 (BM, CAS, CR, F, LL, MEXU, MO, US); 12 Jul 1985 (fl.), Haber & Bello 1984 (BM, COL, CR, F, LL, MEXU, MICH, MO, P, UPS, W); 29 Nov 1985 (fr.), Haber ex Bello 3591 (BM, CR, LL, MO); 17 Dec 1985 (fr.), Haber ex Bello 3935 (CAS, CR, DUKE, F, LL, MEXU, MO); 22 Jul 1986 (fl.), Haber ex Clagget 5738 (CR, F, GH, LL, MO); 27 Feb 1987 (fr.), Haber & Bello 6728 (INB, LL, MO); Feb 1988 (fr.), Haber & Bello 8213 (F, INB, LL, MO, US); 20 Jul 1989 (fl.), Haber 9303 (CR, F, INB, MEXU, MO, TEX), 26 Mar 1990 (fr.), Haber 9815 (CR, INB, MO, US); 30 Jul 1991 (fl.), Haber & Zuchowski 10789 (CR, INB, MO), 12 Mar 1992 (fr.), Haber & Zuchowski 11048 (CR, F, INB, MO); 27 Jul 1977 (fl.), Hartshorn 1901 (CR, F); 25 Jan 1984 (fr.), Pennington 11432 (CR, MO). MEXICO. Veracruz: Estación Biológica Los Tuxtlas, 18°35'N 95°05'W, 300 m; no date (st.), Calzada 79 (F); 29 Nov 1974 (fr.), Cedillo 436 (MO); 4 Feb 1986 (fl.), Cedillo 3518 (MO); 24 Jun 1984 (fl.), Ibarra & Sinaca 1787 (MO); 15 Nov 1984 (fr.), Ibarra et al 2124 (MO); 20 Jan 1985 (fr.?) Ibarra & Sinaca 2240, 2241 (MO); 4 Jul 1985 (fl.), Sinaca 122 (MO). Distribution and biogeography. Gymnosporia haberiana is disjunct between one site at about 300 m elevation in Veracruz, México, and several wet-forest, mid-elevation sites in NW Costa Rica (Fig. 2). However, its nearest relatives, as discussed below, are from low- and mid-elevation South America, a common pattern for tropical Mexican and Central American taxa (eg., Hammel, 1986 and references therein). Nevertheless, G. haberiana is not known from southern Costa Rica or from Panama. This disjunct distribution between Mexico (or northern Central America) and Costa Rica, then skipping Panama to nearest relatives in South America, is not unique (eg. Ziziphus chloroxylon (L.) Oliv.), and if not an artifact of collecting, may itself reflect the complex geological history of the isthmian region. 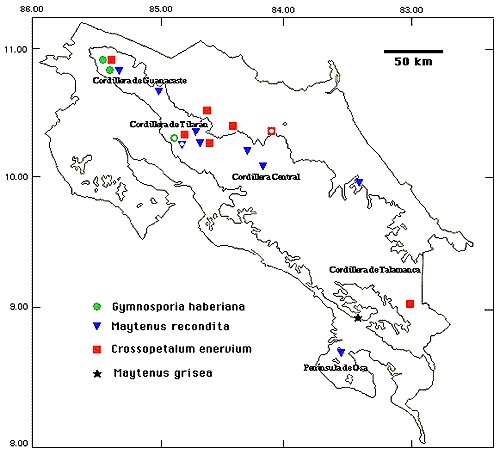 Figure 2. Distribution of four species of Celastraceae in Costa Rica (hollow symbols mark type localities; 500 m contour is indicated). Relationships. The closest relatives of Gymnosporia
haberiana are its two South American congeners. In 1985
Lundell transferred Maytenus magnifolia Loesener to
Gymnosporia and described a second neotropical species,
G. gentryi Lundell, thus reinstating for the New World
an essentially African genus that had come to be considered
congeneric with Maytenus. In contrast to species of
Maytenus, most of which (at least in the neotropics) have
small fasciculate inflorescences, 5-merous flowers, 1 or rarely
2-seeded fruits, and often toothed, gray- or tan-drying leaves, the
New World Gymnosporia have relatively large, open cymose
inflorescences, 4-merous flowers, usually 2--4-seeded fruits, and
entire, greenish-drying leaves (see also Hou,
1955). Study of specimens and bibliographic
material at MO supports Liesner's conjecture
(1993) that G. urbaniana (the
older name) and G. magnifolia are conspecific: Gymnosporia urbaniana (Loesener) Liesner Monogr. Syst. Bot. Missouri Bot. Gard. 45: 1254. 1993: Rhacoma urbaniana Loesener Repert. Spec. Nov. Regni Veg.Vol. 1(11): 162. 1905. TYPE: Perú. Weberbauer 1875 (photo MO). Syn. nov. Gymnosporia magnifolia (Loesener) Lundell Phytologia 57: 314. 1985: Maytenus magnifolia Loesener, Vernh. Bot. Vereins Prov. Brandenburg 48: 176. 1906 (1907). TYPE: Brazil. Amazonas: Flusse Juruá Miry, Ule 5721 (photo MO). The new species differs from G. gentryi of Colombia and Ecuador and G. urbaniana of Amazonian Brazil, Peru and Bolivia by its generally smaller leaves with less conspicuous lateral veins, usually fewer-flowered inflorescences, and sepals that are much broader than long. Gymnosporia urbaniana has smaller flowers with almost no nectar disk and is found mostly in lowland Amazonia. Gymnosporia gentryi, from 1200--2800 m in the northern Andes, has flowers nearly as large as those of G. haberiana, but the ovary (and fruit) is distinctly 4-angled and its leaves have very prominent (below) lateral veins that depart from the midrib at a much narrower angle. By these observations, G. haberiana and G. gentryi appear to be sister species. The three neotropical species of Gymnosporia can be separated according to the following key:
Mexican material of this species has sometimes been tentatively identified as Maytenus grisea Lundell, but examination of the Guatemalan type of that species (Contreras 6944, LL!) shows G. haberiana to be quite distinct. Although the two species are superficially similar because of their somewhat inflated and relatively thin-walled fruits, in contrast to the open cymes and lobed fruits of G. haberiana, the inflorescences of M. grisea are only pedicellate clusters on a very short or obsolete peduncle, the fruits are not lobed, and the leaves are toothed and dry gray rather than green. Unexpectedly, this study has revealed that a rarely collected Costa Rican plant from hills near Palmar Norte (Allen 6327, F [as "Maytenus pallidifolius Standl. & L. O. Williams ined."]; Bohlke 20, F; Hammel et al. 20296, INB; Poveda & Hoet 3050, CR) is certainly conspecific with M. grisea. Thus, M. grisea, known only from the type in Guatemala and now from southern Costa Rica (Fig. 2), is similar to G. haberiana in being disjunct within the region. Etymology and history. This new species is dedicated to my friend and colleague, Dr. William Haber, long-time student and professor of the rich flora of Monteverde, Costa Rica, locality of the type and most other collections cited here. Bill's keen observation and field notes brought to my attention the dioecious nature of this species. The earliest known collection of this species is from Mexico (Cedillo 436, Nov 1974). In Costa Rica it was apparently first collected in Jul 1977 (Dryer 1589). Maytenus recondita Hammel, sp. nov. TYPE: COSTA RICA. Puntarenas: Cordillera de Tilarán, Monteverde community, 10°18'N 84°48'W, 1350 m elev., 11 Jul 1989 (fl.), Haber & Zuchowski 9286 (holotype, INB; isotypes, CR, F, MICH, MO, US). Figure 3. A M. schippii atque M. guyanense inflorescentiis cymoso-paniculatis, foliis in sicco viridi-griseis aut fuscis, praesentia altitudinis majoris differet.
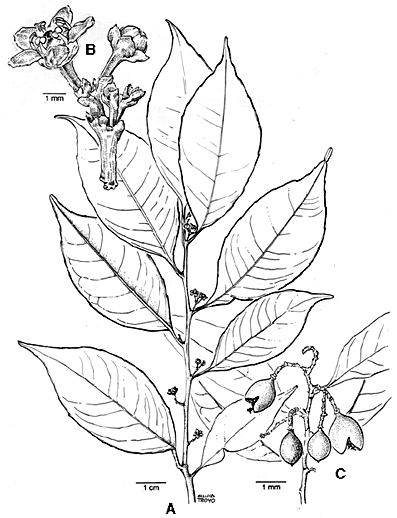
Figure 3. Maytenus recondita. A. flowering shoot; B. pistillate flowers; C. fruits. (A, B from Hartshorn 1895; C from Haber & Zuchowski 8737). Dioecious trees 5--25 m; twigs usually zig-zag, terete. Leaves alternate, (5--) 7--11 x 2--5 cm, elliptic, the apex acuminate, the base decurrent, shallowly toothed in the distal 1/2 or without teeth, with 4--7 indistinct lateral veins, these looping and not reaching the margin, glabrous, drying dark, gray-brown above, gray-green below; petioles 2--5 mm; stipules small, triangular, ± persistent. Inflorescences small, usually 1--4 (--8)-flowered, axillary cymose, panicles, the rachis 1--7 mm, the pedicels 1--2 mm, occasionally proliferating on ± leafless branchlets up to 4 cm long. Flowers 5-merous, ca. 3--4 mm across, pale green; sepals ca. 1/2 or less the length of the petals, rounded, ± erose-margined; petals 1 x 1 mm at base, more or less triangular; stamens yellow, born on the margin of a flat disc ca. 1.5 mm wide, the filaments ca. 0.5--0.6 mm. Capsules bivalvate mostly 1 or 2 (to 6) per axis, obovoid, 10--15 mm, bright orange, usually 1- or occasionally 2-seeded; seeds with a white aril. Additional specimens examined. COSTA RICA. Alajuela: Cordillera de Guanacaste, entre Río Celeste y cabeceras del Río Chimurria 10°43'15"N 85°00'20"W, 700--800 m, Jul. (fl. & fr.), Herrera 2008 (F, INB, MO); Cordillera de Tilarán, altos del Río Caño Negro, 10°21'N 84°48'W, 1300 m, 17 Sep 1989 (fl.), Bello 1376 (INB, MO); (Puntarenas, by error, on labels) Bosque Eterno De Los Niños, 10°23'N 84°42'W, 1100 m, fl. Apr., Bello et al 2156 (CAS, CR, F, INB, MO, W); 10°24'N 84°39'W, 800--900 m, 17 Jul 1993 (fr.), Haber et al 11544 (CAS, CR, F, INB, MICH, MO, W); Parque Nacional Juan Castro Blanco, 10°15'30"N 84°15'30", 1200 m, 26 Jun 1993 (fl., fr.), Jiménez 1327 (COL, CR, F, INB, MO); Parque Nacional Arenal, 10°25'N 84°43'W, 1200 m, 18 Sep 1990 (fl.), Bello 2403 (INB); Peñas Blancas River Valley, 10°20'N 84°50'W, 1250 m, 12 Oct 1985 (fr.), Haber ex Bello 3100 (CR, F, MO); Reserva Biológica de San Ramón, 1017'N 8436'W, 1000 m, 16 Feb 1994 (fl), Herrera 6894 (CR, USJ). Guanacaste: Cordillera de Guanacaste, Parque Nacional Rincón de La Vieja, cabeceras de Quebrada Rancho Grande, 10°46'N 85°49'W, 1350--1400 m, 2 Dec 1987 (fr.), Herrera 1474 (MO); Cordillera de Tilarán, 3 km N Santa Elena, ca. 1020'N 8450'W, 1500 m, near 20 Dec 1985 (fr.), Haber ex Bello 3824, (CR, F, MO), 3831, (CR, F, MO), 3863 (CR, MO); 5 km N Santa Elena on road to Las Nubes, 10°22'N 84°49'W, 1400 m, 10 Nov 1988 (fr.), Haber & Zuchowski 8737 (INB, MO); Río Negro de Tilarán, 1500 m, 4 Oct 1985 (fl., fr.), Haber & Bello 2989 (CR, F, MO); 10°21'N 84°49'W, 1400 m, 27 May 1987 (fl.), Haber & Bello 7125 (F, MO). Heredia: Cordillera Central, vicinity of Vara Blanca, 1710 m, Apr 1938 (fl.), Skutch 3765 (MO, US); Volcán Barva, 1700--2000 m, 24 May 1972 (fl., fr.), Stone 3261 (CR, F, MO). Limón: Cordillera de Talamanca, Z. P. Barbilla, frente a confluencia entre Río Caño Seco y Río Dantas, 10N 8326'W, 150--350 m, 3 Nov 1988 (fl.), Herrera 2278 (F, MO). Puntarenas: Cordillera de Tilarán, Monteverde Reserve and vicinity, ca. 10°20'N 84°50'W, 1000--1700 m elev., 25 May 1989 (fl.), Bello 922 (MICH, INB, MO, TEX, US); 21 Nov 1991 (fl.), Bello 4164 (INB); 17 to 20 Mar 1973 (fr.), Burger & J. Gentry 8601 (CR, F); 30 Oct 1976 (fl., fr.), Dryer 933 (CR, F, MO); 15 Dec 1976 (fr.), Dryer 1082 (CR); 13 Jan 1977 (fl., fr.), Dryer 1125 (CR, F, MO); Gentry & Haber 48761 (F, MO); 20 Mar 1976 (fl., fr.), Gómez-Laurito et al. 1402 (USJ); 5 Aug 1978 (fl., fr.), Haber 162 (CR); Haber 446 (F, MO), 692 (F, MO), 693 (F, MO); 20 Oct 1985 (fl.), Haber ex Bello 3108 (CR, MO); 20 Oct 1985 (fr.), Haber ex Bello 3117 (CR, F, MO); 10 Dec 1985 (fr.), Haber ex Bello 3673 (CR, MO); 16 Jan 1986 (fl., fr.), Haber ex Bello 4266 (CR, F, MO); 2 May 1974 (fl.), Hartshorn 1470 (CR 4 sheets, F, MO); 19 Feb 1976, (fr.), Hartshorn 1820 (F); 26 Jul 1977 (fl.), Hartshorn 1895 (CR 3 sheets, F, MO); 19 Aug 1995 (fl., fr.), Penneys & Zuchowski 683 (CR, F, INB, MO, TEX); 8 Aug 1975 (fl., fr.), Poveda 1110 (CR, F, MO, USJ); Cordillera de Tilarán, Ojo de Agua, Río Aranjuez, 10°17'N 84°46'W, 1550 m, 14 Nov 1987 Haber & Bello 7738 (F, MO); San Luis, Cerro Amapola, 10°16'33"N 84°47'45", 1100 m, 23 Nov 1993 (fr.), Fuentes 583 (INB, MO); Península de Osa, Quebrada Agua Buena, 0842'40"N 8331'00"W, 500 m, 13 Mar 1996 (fl.), Aguilar 4524 (CR, INB, F, MO). PANAMA: Chiriquí: Bajo Mono-Robalo trail ca. 1500--2100 m, 27 Jul 1947 (st.), Allen 4844 (F). Distribution. Except for one sterile collection from western Panama, Maytenus recondita is restricted to wet forests of central Costa Rica (Fig. 2), primarily at 700--1700 m elevation on the Pacific slope from the Cordillera de Guanacaste to the Cordillera Central, with one low-elevation, Atlantic-slope collection from the Río Barbilla region, Cordillera de Talamanca, (Herrera 2278) and another isolated collection from the Osa Peninsula (Aguilar 4524). Characterization. This species is recognized by the acuminate and decurrent, gray-green drying leaves and the paniculate inflorescence of reduced or aborted cymes. It is generally a larger tree as compared to M. segoviarum Standl. & Steyerm., with which it is sympatric in Costa Rica. It has been identified in the past as M. schippii Lundell "vel. sp. aff.," but differs from M. schippii by its shorter petioles, branched, pedunculate (rather than fasciculate) inflorescences, and higher-elevation habitat. Another similar species, currently identified in Costa Rica as M. guyanensis Klotzsch ex Reissek, differs by its much larger, usually darker drying leaves, fasciculate inflorescences, and lower-elevation habitat. The five species of Maytenus recognized for Costa Rica can be distinguished by the following key:
Anthers on flowering material of M. recondita also bearing young fruits appear (on close examination) not to bear pollen. Although the relative length of stamens and style does not differ strikingly in staminate and pistillate flowers, as in the above-discussed species of Gymnosporia, M. recondita must also be dioecious. A series of collections from Costa Rica (U. Chavarría 179, INB; Chávez 248, INB; Ezpinoza 1186, INB; G. Herrera 850, INB; A. Rodríguez 188, INB) is here specifically excluded from the concept of M. recondita. This entity shares the elevational range, inflorescence type, and leaf color of M. segoviarum, but has a leaf shape more similar to that of M. recondita. It is not yet decided whether these collections represent a sixth species for Costa Rica, hybrids between M. recondita and M. segoviarum, or simply variation within M. segoviarum. Etymology and history. The epithet recondita ("hidden,
unpretentious") was chosen for this species because its primary
distinction is its reduced but nevertheless branched inflorescence.
In addition, although the species has been well-collected and
distributed to various herbaria for many years, it is unremarkable
and has remained undescribed. This species was first collected in Apr
1938 (Skutch 3765). Crossopetalum enervium Hammel, sp. nov. TYPE: COSTA RICA. Heredia: Llanura de San Carlos, S base of Cerros Sardinal, Chilamate de Sarapiquí, 10°27'N 84°04'W, 70--100 m, 2 Jun 1985 (fl., fr.), Grayum & Jacobs 5351 (holotype CR; isotypes F, MO, LL + 3, dist. from MO as C. eucymosum). Figure 4. Differt ab species affinibus foliorum nervis lateralibus inconspicuis, apprime a C. standleyi foliis latioribus sed brevioribus, inflorescentiis ut videtur simplicioribus, a C. gomezii foliis multo minoribus.
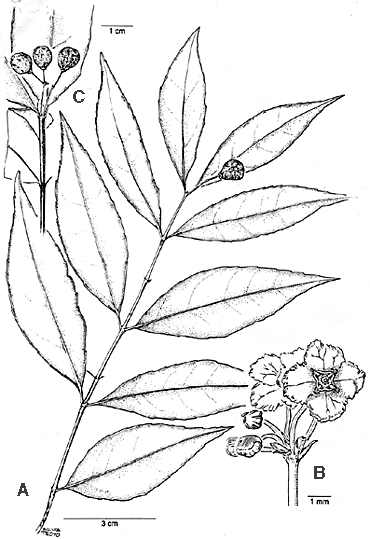
Figure 4. Crossopetalum enervium. A. shoot with flowers and immature fruit; B. flowers; C. fruits. (A, B from Gamboa 48; C from Bello 2232). Shrubs or small trees 1--5 (--8) m; twigs sharply quadrate-ribbed, glabrous. Leaves opposite, 6--10 x 1.8--4 cm, elliptic, the apex acute to acuminate, apiculate, the base acute and decurrent, distantly but sharply serrate, glabrous; main lateral veins 4--6, loop-connected well below the margin, very indistinct above and below; petioles ca. 3--5 mm; stipules minute, caducous extensions of the twig angles. Inflorescences ca. 1--1.5 cm long, mostly 3--7 (--25) flowered, usually only 1-branched, axillary to extra-axillary cymes, glabrous or rarely puberulent; bracts and bracteoles small but ± foliaceous, gland-tipped; peduncles 7--13 mm, the pedicels 1.5--3 mm. Flowers small, 4-merous; sepals ca. 1/2 the size of the petals, rounded; petals ca. 1--1.5 mm, ± orbicular, the usually wine-red margins slightly crenate; stamens yellow, borne from just inside the margin of a flat, circular or quadrate disc; filaments ca. 0.1--0.2 mm. Fruits drupaceous, 0.7--1.4 x 0.6--0.7 cm, obovoid, red to black; seeds tuberculate. Additional specimens examined. COSTA RICA. Alajuela: Cordillera de Tilarán, Monteverde Reserve, Peñas Blancas river valley, ca. 10°20'N 84°45'W, 800-1100 m, 22 Feb 1989 (fl.), Bello & Cruz 724 (INB, MV); 29 Nov 1986 (fl., fr.), Haber ex Bello 6480 (CR); Reserva Forestal de San Ramón, 800--1000 m, 4 May 1985 (fr.), E. Rojas s. n. (USJ); Llanura de San Carlos, Pital, Yucatán 10°34'40"N 84°11'00"W, 100 m, 4 Oct 1994 (fr.), Estrada 246 (INB); Llanura de Guatuso, 3 km NW of Florencia 10°23'N 84°28'W, 250 m, 28 Dec 1993 (fl., fr.), Haber & Guindon 11759 (CR, F, INB, LL, MO, US); 8 km NE of Villa Quesada, near Artezalea, 550 m, 17 Feb 1966 (fr.), Molina et al. 17271 (CR, F); Pueblo Nuevo, 1100 m, 15 Apr 1939 (fl., fr.), A. Smith 1902 (F). Guanacaste: Cordillera de Guanacaste, P. N. Guanacaste, Volcán Cacao, ca. 10°55'45"N 85°28'15", 1100 m, 11 Feb 1995 (fl.), Alfaro 114 (INB); 2 Jun 1990 (fr.), Bello 2232 (INB); 11 Apr 1991 (fl., fr.), Chávez 543 (CR, INB); 14 Jul 1991 (fr.), Chávez 585 (CR, INB); 8 Feb 1995 (fl., fr.), B. Gamboa 48 (CR, INB, MO); 10 Feb 1995 (fl., fr.), Lobo 50 (INB); 3 Jun 1990 (fr.), Maass 14 (INB); 2 Jun 1990 (fr.), Obando 15 (CR,); P. N. Rincón de La Vieja, 800 m, 27 Jan 1983 (fl.), Garwood et al 726 (F). Heredia: Llanura de San Carlos, near Tirimbina E of the Río Sarapiquí, 10°24'N 84°07'W, 150--250 m, 12--15 Aug 1971 (fr.), Burger & Burger 8053 (CR, F). Puntarenas: Cordillera de Tilarán, Monteverde area, Hoge Middle, 13 Mar. 1979 (fl.), Koptur SK-102 (CR); Cordillera de Talamanca, P. I. La Amistad, Estación Altamira, 09°02'10"N 83°01'20"W, 1350 m, 14 Apr 1995 (fl.), Angulo 164 (INB); Las Alturas and vicinity, 1700 m, 26 Aug 1974 (fr.), Maas 1484 (CR, F). Distribution. Crossopetalum enervium is apparently endemic to Costa Rica, where it is known from both the northern and southern parts of the country in wet to very wet forest from ca. 100 to 1700 m elevation (Fig. 2). A study of shrub and treelet species at Monteverde (Koptur et al., 1988), includes phenological data about this species (as C. eucymosum), vouchered by the above cited Koptur collection (Haber, pers. comm); fruits take about 3 months to mature and then may remain on the plant up to 5 months. Most collections are from the northern half of the country in the Cordilleras de Guanacaste and Tilarán and from the Caribbean lowlands to the E. A few collections with slightly more prominent venation, approaching that of C. standleyi, come from an outlying population in the extreme S of the country on the Pacific slopes of the Cordillera de Talamanca very close to Panama. Although not yet known from Panama, this new species must certainly occur there. Characterization and relationships. This species is distinctive for the very faint (when dry) lateral veins and the delicate, subumbellate inflorescences. In leaf size and certain aspects of the inflorescence and fruit, this species is most like C. standleyi (Lundell) Lundell (basionym Myginda standleyi; [Standley 68938, istotype F!]), from which it differs by its wider and slightly shorter leaves, markedly obscure venation, and the subumbellate cymes with relatively long primary peduncles. Both C. standleyi and C. enervium have obovate fruits and short anther filaments. Crossopetalum enervium is also similar to the type of C. riparium (Lundell 1476, F!) in its smallish leaves and delicate, glabrous inflorescences. The inflorescences of the latter species, however, are more distinctly branched, and its leaves dry brownish (rather than gray-green), and are less serrate and more prominently nerved than those of C. enervium. The stamens are nearly sessile on the type of C. riparium. Both C. riparium and C. standleyi (originally described from Guatemala) are restricted to northern Central American. That the close relatives of this new species are from N of Costa Rica is to be expected; the genus is basically Central American and West Indian, with only one species reported from South America (Gentry, 1993). One collection of C. enervium, Chávez 543, has slightly puberulent inflorescences. Among Costa Rican Crossopetalum, the new species is most similar to C. gomezii Lundell, which differs by having much larger leaves with very prominent venation. It has been confused with the most common Costa Rican species, formerly known as C. eucymosum (Loesener & Pittier) Lundell, here considered a synonym of the older C. parviflorum (Hemsley) Lundell: C. parviflorum (Hemsley) Lundell Wrightia3(1): 8. 1961: Euonymus parviflorus Hemsley, Diag. Pl. Nov. Mex. p. 6. 1878. TYPE: Nicaragua, Chontales, R. Tate 292 (photo! US). Syn. nov. Crossopetalum eucymosum (Loesener & Pittier) Lundell Wrightia3(1): 7. 1961: Myginda eucymosa Loesener & Pittier, Contr. US. Natl. Herb. 12:175. 1909, Pl. 18. TYPE: Guatemala. Alta Verapaz: Cahabon River, Pittier 239 (US!). Crossopetalum parviflorum has much larger, more open, pubescent inflorescences, larger flowers with longer stamens, and leaves that dry brownish instead of gray and have more obvious secondary venation. The five Costa Rican species of Crossopetalum can be distinguished by the following key:
Etymology and history. The epithet "enervium" is chosen in reference to the inconspicuous lateral venation of the leaves in dry specimens. Here again "reconditum" was a serious contender for the honors; this rarely collected species has actually been in collections for quite long, the earliest known gathering being that of Austin Smith 1902, in 1939. That collection and others, including the type, were distributed as Crossopetalum eucymosum, wherein the new species has, until now, remained hidden. Acknowledgements. I thank Michael Grayum for his ever careful reading of an early draft of the mss, two anonymous reviewers for further refinements, Silvia Troyo for the line drawings, and Henk van der Werff for fine tuning the latin descriptions. The photographs of Gymnosporia haberiana and Maytenus recondita were supplied by William Haber. Field work resulting in many of the collections here cited was supported by various National Geographic Society grants to William Haber and to Michael Grayum and by National Science Foundation (NSF) grant BSR-8700068 to M. H. Grayum and the author. Publication was supported by NSF grant DEB-9300814 to the author and M. H. Grayum.
|
{kind=link}
{kind=link}
{kind=link}
© 1995-2025 Missouri Botanical Garden, All Rights Reserved
4344 Shaw Blvd.
St. Louis, MO 63110
(314) 577-5100
Technical Support