QUICK SEARCH
MO PROJECTS:
Africa
Asia/Pacific
Mesoamerica
North America
South America
General Taxonomy
Photo Essays
Training in Latin
America
MO RESEARCH:
Wm. L. Brown Center
Bryology
GIS
Graduate Studies
Research Experiences
for Undergraduates
Imaging Lab
Library
MBG Press
Publications
Climate Change
Catalog Fossil Plants
MO DATABASES:
W³MOST
Image Index
Rare Books
Angiosperm
Phylogeny
Res Botanica
All Databases
INFORMATION:
What's New?
People at MO
Visitor's Guide
Herbarium
Jobs & Fellowships
Symposium
Research Links
Site Map
Search
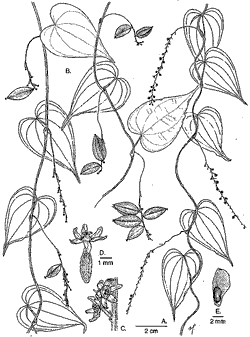
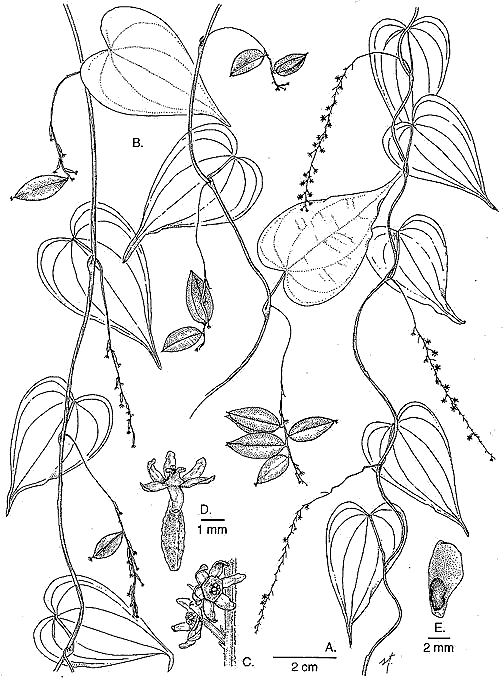
A New Species of Dioscorea from Costa RicaBarry E. Hammel
Novon 10: 378--381. 2000. Published on 14 December 2000. © Missouri Botanical Garden 2000. Abstract. Dioscorea natalia (Dioscoreaceae) is described from wet to very wet areas on both Pacific and Caribbean slopes of Costa Rica, from the Monteverde area to Tapantí National Park. The species' confusion with the related D. lepida C. V. Morton is discussed and a key is provided to distinguish it from that species and the other two closely related Costa Rican species, D. racemosa (Klotzsch) Uline and D. standlyei C. V. Morton. During the course of work on a treatment of Dioscorea for the Manual de Plantas de Costa Rica, material of three possible new species was set aside for further study. Careful examination and review demonstrated that one of them would not be separated from the recently described panamanian D. davidsei O. Téllez, and that the other corresponds to D. lepida. The third, here described as D. natalia, had previously been identified as D. lepida and with that species, is closely related to two other Costa Rican species, D. racemosa and D. standleyi. Dioscorea natalia Hammel sp. nov. TYPE: Costa Rica. Cartago: Parque Nacional Tapantí, Sendero Oropendula, 1400 m, 26 Sep 1997 (stam. & pist. fl., fr.) Hammel & Troyo 21068 (holotype INB; isotypes CR, F, MEXU, MO). Figure 1.
Species subtilis D. racemosae affinis, a qua imprimis differt inflorescentiis brevioribus, paucior-floribus rachidibus exilioribus. Stems delicate, twisting to the right but also very rarely to the left. Leaves alternate, ovate to narrowly triangular-cordate, 4.5--6 (--9.5) X 2.5--3.5 (--5) cm, 7 (9)-nerved, glabrous, very faintly glandular-dotted on the lower surface. Staminate inflorescences 4--8 (--10) cm, , simple racemes to racemose, usually 1 to rarely as many as 5 per axil; the rachis ± verruculose, very delicate, up to only twice the diameter of the pedicels; the cymes of 1 (rarely 2) flowers, subtended by a pair of narrowly triangular bracts 1--1.5 mm; pedicels 1.5--2 (--4) mm. Staminate flowers with tepals appearing nearly valvate in bud, broadly triangular, 0.8--1.1 mm, patent, pale yellowish-green or cream; stamens 3, connivent in the center of a +- hexagonal torus; filaments ca. 0.1--0.15 mm, united only at the very base; anther cells ± ovoid ca. 0.15--0.2 mm, touching those of adjacent anthers, dehiscing upwards; staminodia none. Pistillate inflorescences 5--8 cm, spikes. Pistillate flowers with tepals as in the staminate ones but to 1.25 mm; styles 3, united in a conical stylar column 0--0.25 mm, the free portion, including stigmas, ca. 0.4 mm; stigmas 3, simple; staminodia lacking. Fruits perpendicular to the rachis, elliptic, 1.2--2.0 x 0.5--0.9 cm, apically acute; seeds 1--1.4 x 0.3--0.4 cm, winged on the posterior end. Paratypes. Alajuela: Reserva Forestal San Ramón, 1100 m, Apr 1991 (fl.), Bittner 972 (CR); between San Miguel and La Palma de San Ramón, Nov 1926, Brenes (278) 5124; Monteverde Reserve, Río Peńas Blancas, 900 m, Aug 1988 (fl.) Bello 248 (CR); Guanacaste, Monteverde, 1 km al N de Las Nubes, 1300 m, Sep 1988 (stam., pist. fl., fr.), Haber & Zuchowski 8675 (INB, MO); road to Las Nubes, 1400 m, Nov 1988 (stam., pist. fl.) Haber 8749 (INB); Puntarenas: Monteverde Reserve, 1 km SW of Station, 1500 m; Feb 1992 (fl.), Ingram & Ferrell 1298 (CR); Monteverde vicinity, Upper Río Negro valley, 1500 m, Sep 1985 (fl, fr.), Haber ex Bello 3020 (CR); Monteverde vicinity, San Gerardo Biological Station, 1200 m, Feb 1995 (pist. fl., fr.), Penneys 167 (CR, F, INB, MO, US); San Jose: Río Hondura, 1100 m, Feb 1974 (fl), Lent 3791 (CR); Cartago: Cańón del Río Grande de Orosí, Oct 1983 (fl., fr.), Chacón et. al. 1483 (CR--6 mounted duplicates!); Parque (Refugio) Nacional Tapantí, 1250 m, Aug 1991 (stam. fl.), Hammel et al 18324 (INB, MO); 1400 m, Dec 1992 (fl.), Herrera 5813 (CR); Jun 1994 (stam. fl.), Morales & Carnevali 2873(INB); Valle del Reventazón, Moravia de Chirripó, 1100 m, Sep 1993 (stam. fl.), Campos & Campos 186 (INB). Distribution. This species, apparently endemic to Costa Rica, is known from some of the very wettest parts of the country, especially P. N. Tapantí. However, it is also known from the Las Nubes area of Monteverde, which is relatively dry. So far it is known only from 900--1500 m, from Monteverde and Tapanti, and several intervening localities. Etymology. It was my pleasure to defer to Silvia Troyo, illustrator of this new species, the honor of choosing its name. When she came across the species for the first time in the field, she was struck by the same joy of discovery as any taxonomist upon finding such a new species, so tiny and precious. For all good reason, she chose the name Natalia, one often given here to first-born daughters, and that of her own. The epithet "natalia" is used intentionally as a word in apposition with no Latin termination as sanctioned by the Saint Louis Code, Art. 23.1 (Greuter et al., 2000) and therefore not to be corrected (Art. 60.11) by well-meaning Latinists. Discussion. Dioscorea natalia is distinct from the closely related Costa Rican species (D. lepida, D. racemosa, D. standleyi) by its consistently very small leaves and small, delicate inflorescences whose slender rachis is usually less than twice the diameter of the pedicels. The flowers are almost always solitary along the rachis, and are born on rather long pedicels up to 4 mm. Some specimens of D. racemosa have nearly identical, also solitary flowers. However, the leaves are much larger, the inflorescences longer and thicker, and the flowers are more densely clustered. Although specimens of this species, examined for Flora Mesoamericana, were originally identified as D. lepida, the two are easy to distinguish; D. lepida always has much larger leaves, much-branched staminate inflorescences, and +- ligulate, rather than broadly triangular tepals. Since D. natalia had originally been determined as D. lepida, a discussion concerning our understanding of the latter is in order. Partly because the original description was based only on fruiting material (Morton, 1936), D. lepida has often been wrongly identified and poorly characterized (cf. Morton 1945, Standley 1937). Early in my study of the Costa Rican Dioscorea I had considered this material to be a new species distinct from related species by virtue of its abundantly branched staminate inflorescences, very broadly cordate, many-veined and often opposite leaves. None of these features was mentioned in the original description of D. lepida, nor used to distinguish it in intervening floristic treatments. The one feature, visible in the type, that taxonomists have used to form an (erroneous) concept of this species is the apically acute fruits (cf. Téllez & Morton, 1994). It is now known that material grouped by that character alone can include several species; D. lepida, D. natalia, D. racemosa, and D. standleyi all have rather small, often or always apically acute fruits. If one additional character should be chosen as that which allowed a clear delimitation of D. lepida, it would be the position of the mature fruits, i.e., reflexed (see key) by the pedicels towards the base of the inflorescence so that they lie nearly parallel to the rachis. This is consistent in all fruiting collections and all observed populations of D. lepida, and contrasts with the ± perpendicular or even apically directed fruits of the related species here discussed. That, in conjunction with the narrowly acute fruits, much-branched staminate inflorescences, broad, many-veined, and often opposite or even ternate leaves (at lower nodes), as well as the rather high and narrow elevational range, help distinguish D. lepida from its close relatives in Costa Rica. In addition, D. lepida is unusual in that plants of all populations examined were monoecious instead of dioecious, as most species of Dioscorea are considered to be. Dioscorea lepida stems apparently die back during the dry season and the new shoots, appearing at the beginning of the rainy season, flower precociously with staminate flowers. As the stems elongate they begin producing pistillate flowers towards the apex with a few intermediate inflorescences bearing both sexes and even some apparently hermaphroditic flowers. By the time the plants are in late flowering stage or fruiting, there may be little or no evidence of the staminate inflorescences lower down. Such a stituation can easily give rise to herbarium specimens that bear only one sex. Field or greenhouse observations of other species known only from collections, might demonstrate monoecy to be more widespread in Dioscorea than previously thought. The terminology used here, reflexed (downturned) versus upturned, relative to the axil, would be reversed relative to gravity since the infructescences are pendent in living plants.
Acknowledgments. This work was supported, in part, by funds from the National Science Foundation through a grant (DEB-9300814) to the author and co-PI M. H. Grayum for the Manual to the Plants of Costa Rica. I thank Osvaldo Téllez for his useful comments on an early draft of the manuscript and Silvia Troyo for her excellent line drawings of the new species.
|
{kind=link}
{kind=link}
© 1995-2025 Missouri Botanical Garden, All Rights Reserved
4344 Shaw Blvd.
St. Louis, MO 63110
(314) 577-5100
Technical Support