A revision of the genus Orthostichella (Neckeraceae)
Orthostichella C. Müll. is a genus
of predominantly epiphytic, frequently pendulous mosses of tropical and
subtropical American-African distribution. Schwägrichen (1816) described the
oldest species now placed in Orthostichella (as Hypnum hexastichum Schwägr.),
but the species generally accepted in the genus were first brought together by
Müller (1850) under Neckera Hedw. sect. Pseudopilotrichum. C.
Müll. subsection Orthostichella C. Müll. Müller (1879) began using his
subsection Orthostichella at the generic level, however, he did not
indicate this at the time, and because three names were placed in the genus, it
is considerd invalid. Müller (1890) again used Orthostichella at the
generic level, but once more this name is invalid because he described five
species in the genus. In 1897 Müller yet again used Orthostichella at
the generic level, but because he described only one new species the genus
becomes validated under Article 42 (dealing with descriptio generico-specifica
provisions) of the International Code of Botanical Nomenclature (Greuter 2000).
Orthostichella has long been
associated with Pilotrichella (C. Müll.) Besch. The relationship between
the two taxa began when Bescherelle (1872) elevated Müller’s (1850) Neckera
Hedw. sect. Pseudopilotrichum. C. Müll subsect. Pilotrichella to
generic rank. The newly established Pilotrichella was broadly conceived
and included four sections: Orthostichella (C. Müll.) Besch., Eupilotrichella
(C. Müll.) Besch., Papillaria (C. Müll.) Besch., and Meteoridium
(C. Müll.) Besch. Papillaria had previously been removed from this group
by Lorentz (1864). Jaeger & Sauerbeck (1877) refined Pilotrichella
by dividing it into two unranked groupings: Eupilotrichella (including
Bescherelle’s sect. Meteoridium) and Orthostichella. Section Meteoridium
was removed from Pilotrichella by Brotherus (1906) who also positioned
the genus (with sections Orthostichella and Eupilotrichella) in
the tribe Meteorieae. This association of Orthostichella with Pilotrichella
and the placement of Pilotrichella in the Meteoriaceae was followed
by Fleischer (1908), Brotherus (1925), Bartram (1949), Florschütz (1964),
Walther (1983), Vitt (1984), Spessard-Schueth (1994), Churchill & Linares
(1995), Duarte-Bello (1997), and Magill & van Rooy (1998).
Orthostichella differs
significantly from Pilotrichella in its smaller plant size, and in often
having primary stolons as well as stipitate stems. It also differs from Pilotrichella
in having spirally ranked leaves with sparsely developed alar cells, and often
its leaves have a single or ddouble costae. Sporophytically Orthostichella
differs from Pilotrichella in having shorter setae and smooth to
papillose exostome teeth. Also the presence of large spores (to 64 μm) in Pilotrichella distinguishes the two taxa.
In addition to Müller’s treatments (1879, 1890, 1897), Orthostichella
has been recognized as a genus distinct from Pilotrichella by Buck
(1994, 1994a), Buck & Goffinet (2000), Gradstein et al. (2001),
Allen & Magill (2003), and Goffinet & Buck (2004).
The Meteoriaceae are usually placed in the
Leucodontales (Fleischer 1908, Brotherus 1925, Walther 1983, Vitt 1984). The
family, however, was transferred to the Hypnales by Buck (1994) and placed near
the Brachytheciaceae in part because its exostome teeth are often horizontally
striate at base and it lacks stolon-like primary stems that are tightly
adherent to the substrate as well as greatly reduced stolon leaves. Buck (1994,
1994a) also reconsidered Pilotrichella and its systematic placement. As
a result Orthostichella, Pilotrichella, Weymouthia Broth.
and Squamidium (C. Müll.) Broth. were transferred to the
Lembophyllaceae.
There are a number of tropical and
subtropical pleurocarpous genera that grow pendulous in predominately epiphytic
habitats. These genera present classification problems because they appear to
represent several phylogenetic lines as judged by their very different
peristomial forms, but they exhibit considerable convergence in their gametophytic
features. Orthostichella is one of these problematic genera. It is
difficult to decide if it belongs in the Meteoriaceae or Lembophyllaceae
because its reduced peristome shows affinities to both families and it is hard
to determine whether its gametophytic features are indicative of propinquity of
descent or convergence. This situation is further complicated because the
Meteoriaceae and Lembophyllaceae are so similar (e.g., compare the
family descriptions of the Lembophyllaceae and Meteoriaceae in Buck &
Goffinet 2000).
The leaves and stems of most genera placed
in the Meteoriaceae often have parts that are intensely black. This odd feature
is usually only noticed in passing, but it is so distinctive that when present
one can immediately assign unknown specimens to the Meteoriaceae. Genera placed
in the Lembophyllaceae can be green, yellowish red or brown, but they never
exhibit this intense, at times shiny, black color. Furthermore, all of the
genera now placed in the Meteoriaceae that can produce this intense black
coloration also have single costae. On the basis of these two features it
appears that Squamidium should be returned to the Meteoriaceae. Pilotrichella
and Weymouthia which are ecostate or have short double costae and a
green, yellowish red or brown coloration seem properly placed in the
Lembophyllaceae. Orthostichella on the other hand seems better placed in
the Neckeraceae by virture of its stolon/stipe morphology, poorly and variably
developed leaf costae, and yellowish
white neckeroid peristome. Within the Neckeraceae the genus is isolated by
virtue of its non-complanate leaves that are often arranged in spiral rows.
Orthostichella
has a complex morphology with creeping primary stems and erect or pendent secondary stems.
Its stolons, stems, and branches are identical in morphological structure. It lacks a stem central
strand, and its axillary hairs are usually reddish throughout. The secondary stems can be strongly
stipitate or evenly foliate throughout. Remarkably, stipitate and evenly foliate secondary stems
can sometimes be found on the same primary stem. Its branches often end in filiform attenuations
or stolons. These different structures can be morphologically distinct, but more
often they intergrade one into another or they can be abruptly transformed from
one structure to another. As a rule the leaves on the branches, and sometimes
the secondary stems are variously spirally ranked while those on the stolons
and primary stems are unranked. But, this rule is often violated. Most leaves
are cuspidate, have distinctive incurved, serrulate upper leaf margins,
elongate, smooth, firm-walled leaf cells, and weakly developed, reddish yellow,
firm-walled alar cells. The costa in O. versicolor is one of its oddest
features. In some species most leaves are ecostate, however, here and there
leaves can be found with short double, short single, or long single costae. In
other species most leaves have a long single or long double costa but here and
there ecostate leaves can be found. Gametophytic features of Orthostichella
that show significant variation include: 1. relative plant size; 2. leaf shape;
3. leaf stance; 4. costa development; and 5. leaf apex shape.
The sporophytes of Orthostichella are
rarely encountered but, they are uniform throughout the
genus. The setae are elongate-flexuose and variously papillose roughened above.
The genus has ovoid to short-cylindrical capsules, long-rostrate opercula, and
mostly hairy, cucullate calyptrae. The Orthostichella peristome is
diplolepideous and reduced with yellowish white exostomes and endostomes that
are nearly the same length as the exostome teeth. The more or less linear
exostome teeth are lightly horizontally striate on the dorsal (outer) surface
at base. The endostome has a low basal membrane with filamentous, narrowly
perforated segments, and cilia are usually absent. Even though the Orthostichella
peristome is significantly reduced in form, the presence at the base of the
exostome teeth of horizontal striae indicates the peristome is basically hypnoid.
The name Orthostichella combines the
Greek orthos (straight) and Stichos (row) with the Latin
substantival suffix -ella (diminutive). The name refers to the strong
tendency for ranked leaves in the genus, although the branch leaves are often
in spiral rather than straight rows.
Orthostichella
C. Müll., Bull. Herb. Boissier 5: 204. 1897.
Type: O. filamentosula C.
Müll. (Buck 1994).
Neckera sect. Pseudopilotrichu
C. Müll.,Syn. 2: 123. 1850. Pseudopilotrichum
(C. Müll.) Buck & Allen in Buck, J. Hattori Bot. Lab. 75: 69. 1994.
Lectotype: N. hexasticha Schwägr. C. Müll. (Buck 1994).
Neckera sect. Pseudopilotrichum
subsect. Orthostichella C. Müll., Syn. 2: 123. 1850. Lectotype: N.
hexasticha Schwägr. C. Müll. (Buck 1994).
Plants
slender or medium-sized, dull, yellow-green, reddish yellow or brownish red, in
dense or loose mats often with pendent strands. Stolons red, creeping; leaves
widely spaced, reduced, erect, broadly ovate or ovate-triangular, abruptly
long-pointed, ecostate or costa short and double. Primary stems occasionally
stipitate, horizontal or erect, frequently and irregularly branched, at times
with flagelliform tips, yellowish green, becoming dark red with age, in cross
section with sclerodermis, enlarged cortical cells and no central strand
absent; paraphyllia absent, pseudoparaphyllia absent, scale leaves present;
axillary hairs 3–4 cells long, basal 1 cell quadrate to subquadrate, reddish
brown, upper 2–3 cells broadly cylindrical, hyaline or reddish; rhizoids only
on stolons red, not or irregularly branched, from clusters of initials abaxial
to the leaf insertions. Stipe leaves ovate below, piliferous above, auriculate,
cordate, rounded at base or straight to the insertions. Stipe leaves ovate,;
costae short and double; alar cells well developed. Secondary stem and branch
apices turgid; stem leaves not seriately ranked, ovate, erect, variously
clasping below, rounded to the base, abruptly narrowly acuminate, at times
hair-pointed; branch leaves obscurely seriately ranked when dry, distinctly so
when wet, leaves panduriform, broadly oblong, obovate, oblong-obovate, or
broadly obovate, erect to erect-spreading, appressed and at times clasping
below, abruptly flexed and spreading above, stem leaves often auriculate, apex
abruptly acuminate, cuspidate to long-cuspidate; margins broadly incurved,
sharply serrulate above, plane or broadly incurved, serrulate below, entire or
weakly toothed on basal auriculation; costae short double, short single, long
single, or absent; leaf cells long flexuose, smooth to slightly porose, outer
basal cells shorter and broader, inner basal cells elongate, porose, alar cells
subquadrate to short-rectangular extending up the margins in 2–4 rows and across
the insertion in 2–4 rows, yellow across the insertion. Asexual reproduction by
deciduous leaves. Dioicous. Perigonia gemmate, lateral on secondary stems and
branches, perigonial leaves ovate, acuminate, costa absent; paraphyses present,
antheridia few. Perichaetia lateral, paraphyses and archegonia numerous;
perichaetial leaves lanceolate, vaginula often densely hairy. Setae short,
wavy, smooth below, roughened above, yellow. Capsules exserted, erect to
somewhat inclined, ovoid to short-cylindrical, neck moderately developed;
exothecial cells small, rounded in several rows below the mouth, subquadrate,
irregularly short-rectangular, or oblate, firm-walled below, cells in neck
smaller than the median cells; stomata superficial on neck; opercula obliquely long-rostrate;
annuli rudimentary, consisting of 2–3 rows of thin-walled enlarged cells; peristome
yellowish white, exostome teeth linear, dorsal (outer) surface horizontally
striate at base, smooth or finely papillose above, trabeculae weakly developed
on both sides, endostome nearly as long as exostome, basal membrane very low,
segments filamentous, narrowly perforated, not keeled, smooth to papillose,
cilia rudimentary or absent. Spores round, lightly roughened. Calyptrae,
cucullate, hairy or smooth.
Key to species of Orthostichella.
1. Branch leaves spreading from
insertions..........................................................................................
2
1. Branch leaves at base appressed
and clasping stems.......................................................................
5
2. Branch leaves cuspidate; leaves
consistently costate.......................................
4. O. pandurifolia
2. Branch leaves apiculate to
mucronate.......................................................................................
3
3. All leaves with a long, single
percurruct costa.........................................................
3. O. longinervis
3. Leaves mostly ecostate,
sporatically costae, the costa short and double or single and ending at
midleaf...................................................................
..............................................................
4
4. Stem leaves hair-pointed
.................................................................................
7. O. welwitschii
4. Stem leaves cuspidate
.......................................................................................
6. O. versicolor
5. Stipe and secondary
stem leaves rounded to the insertion; setae 1.0–1.5 mm long.............................
.....................................................................................................................
2. O. hexasticha
5. Stipe and secondary stem
leaves auriculate; setae 3.5–4.0 mm long................................................
4
6. Auriculate leaf bases
serrate or dentate.............................................................
1. O. calomicra
6. Auriculate leaf bases entire
...................................................................................
5. O. roseana
1.
Orthostichella
calomicra (Broth.) Allen & Magill, comb. nov.
Pilotrichella calomicra Broth., Bol. Soc. Brot. 8: 180. 1890. Protologue: Săo
Tomé and Príncipe. Ins. S. Tomé, ubi legit Fr. Quintas. Types: Afr.
occ., ins. S. Thomé, leg. Fr. Quintas (H, holotype); Afr. occ. ins. S.
Thomé, leg. Fr. Quintas (Comm. Broth.) (S, isotype); Ins. S. Thomé, leg.
Fr. Quintas (S, isotype).
Pilotrichella isleana Besch. var. virescens Besch., Ann. Sci. Nat.,
Bot. sér. 6, 10: 267. 1880. Protologue. Réunion: Plaine des Cafres, G. De L’isle; Cilaos, hauts du
Matarum, Valentin, 1876.
Types. La Reunion. G.-de l’Isle [Herb.
Mus. Paris] (H, lectotype designated here); La Reunion. M. g.-de l’Isle,
com. Brotherus (S, 2 isolectotypes).
Pilotrichella holstii Broth., Bot. Jahrb. Syst. 20: 197. 1894.
Protologue: Kenya/Tanzania. Usambara; ohne nähere Standorte (Holst n. 698).
Types: Usambara, leg. C. Holst 698 (H, holotype); .Usambara, leg. C.
Holst, com. Broth. (S, isotype).
Pilotrichella muelleri Dusén, Kongl. Svenska Vetensk. Acad. Handl. 28(2):
33. 1895. Protologue: Cameroon.
Habitat in Camerunia in ramulis arborum, ubi supra Beam pagum c. 1,600 metra
supra mare m. Julio a. 1891 legi [Dusén]. Types: Africa
occ. in monte Camerunensibus in ramulis arborum c. 1600 m. Julii 13 1891, P.
Dusén 262 (S, holotype); Musci Africani in Camerunia a P. Dusén collecti. 262.
Orthostichella Duseni C. M. In montibus Camerunensibus ad Bueam pagum c.
1600 metra supra mare in ramulis arborum die 13 m. Julii a. 1891 (H, NY, S [4
collections], isotypes).
Orthostichella duseni C. Müll., Kongl. Svenska Vetensk. Acad. Handl. 28(2):
33. 1895, invalid name listed in synonymy. Pilotrichella duseni (C.
Müll.) Par., Index Bryol. Suppl. 270. 1900, illegitimate name includes type of
species with priority. Based on Dusén Musci Camerunia n. 262 (H, S).
Plants
slender, dull, yellow-green, reddish yellow or brownish red, in dense mats.
Stolons red, creeping; leaves widely spaced, reduced, erect, broadly ovate,
abruptly long-pointed, ecostate or costa short and double, 0.6-0-8 mm long.
Primary stems occasionally stipitate, horizontal or erect, frequently and
irregularly branched, at times with flagelliform tips, to 5 cm long, yellowish
green, becoming dark red with age, in cross section with sclerodermis of 3–5
small, thick-walled, reddish orange cells, cortex cells enlarged, firm-walled,
pale yellow, red with age, central strand absent; paraphyllia absent,
pseudoparaphyllia absent, scale leaves present; axillary hairs 3–4 cells long,
basal 1 cell quadrate to subquadrate, reddish brown, upper 2–3 cells broadly
cylindrical, hyaline or reddish; rhizoids only on stolons red, not or
irregularly branched, from clusters of initials abaxial to the leaf insertions.
Secondary stem and branch apices turgid; stem and branch leaves differentiated
only in size, obscurely seriately ranked when dry, distinctly so when wet,
leaves panduriform, broadly oblong, appressed and clasping below, broadly obovate,
abruptly flexed and spreading above, stem leaves 1.0–1.5 mm long, strongly
auriculate, apex apbruptly acuminate, long-cuspidate, cuspid 80–140 µm long;
margins broadly incurved, sharply serrulate above, plane or broadly incurved,
serrulate below, weakly toothed on basal auriculation; costae short double,
short single, or absent; leaf cells long flexuose, smooth to slightly porose,
18–40 x 3–5 μm, outer basal cells shorter and broader, inner basal cells
elongate, porose, alar cells subquadrate to short-rectangular extending up the
margins in 2–4 rows and across the insertion in 2–4 rows, yellow across the
insertion. Dioicous. Setae short, wavy, 2–3 mm long, smooth below, roughened
above, yellow. Capsules exserted, erect to somewhat inclined, ovoid to short-cylindrical,
1.0–1.5 mm long, neck moderately developed; exothecial cells small, rounded in
2–3 rows below the mouth, subquadrate, irregularly short-rectangular, or
oblate, firm-walled below, cells in neck smaller than the median cells; stomata
superficial on neck; opercula obliquely long-rostrate, 1.5 mm long; annuli
rudimentary; peristome yellowish white, exostome teeth linear, 0.5–0.6
mm long, dorsal (outer) surface horizontally striate at base, smooth or finely
papillose above, trabeculae weakly developed on both sides, endostome nearly as
long as exostome, basal membrane very low, segments filamentous, narrowly
perforated, not keeled, smooth to papillose, cilia rudimentary or absent.
Spores round, lightly roughened, 17–26 μm long. Calyptrae 2 mm
long, cucullate, hairy.
Etymology.
The specific epithet calomicron combines the Greek Kalo- (rope)
with micro (small) and refers to its distinctly serriately ranked
leaves.
Distribution.
West-Central and East Tropical Africa, Western Indian
Ocean.
Illustrations.
Dusén (1895, p. 33 a & b; 3f 1).
Ecology.
On trees and shady rocks; 1200–1900 m.
Selected specimens examined
Săo Tomé and Príncipe. Săo
Tomé: Quintas (H).
CAMEROON. Bueam (H, NY, S)
TANZANIA. Arusha: Crosby & Crosby 9119 (BM, FH, H,
MO, NY, S, JE); Tanga: Pócs 8531I (MO).
MADAGASCAR. Antserana: Crosby & Crosby 7180 (H, MO,
NY, S); Antananarivo: Crosby & Crosby
9281 (MO); Fianarantsoa: Crosby & Crosby 6873 (BM, JE, MO,
NY); Toamasina: Magill et al. 9548 (B, FH, G, MO, NY).
REUNION. Arrondissement au Vent: Crosby & Crosby 8980 (B, BM, CAS,
DUKE, E, F, FH, G, H, JE, MEXU, MO, NICH, NY, PC, S, UC, US).
Orthostichella
calomicra is a small to medium-sized
species with closely spaced, panduriform stem and branch leaves. The primary
stems of O. calomicra are often branched close to the stolons, as a
result the plants often form compact mats. The stem and branch leaves are
appressed, clasping below and abruptly flexed and spreading above. The stem and
branch leaves are strongly auriculate at base and the basal margins are nearly
always weakly toothed. The costae of O. calomicra are consistently weak
and the alar cells are poorly developed.
Morphologically
O. calomicra is extremely close to O. roseana which differs in
having a looser habit, entire basal leaf margins, longer setae, and smaller
spores. In size and aspect it is nearly identical to O. hexasticha, and
both species have the same leaf stance. Orthostichella hexasticha
differs from O. calomicra in having stem leaves that are rounded to the
insertion rather than strongly auriculate Orthostichella versicolor has
forms that often have strongly auriculate stipe and, less often, stem leaves.
Furthermore, O. versicolor and O. calomicra are more less similar
in size. Orthostichella calomicra consistently differs from O.
versicolor in having branch leaves that are appressed and clasping at base.
2.
Orthostichella
hexasticha (Schwägr.) Buck,
Bryologist 97: 435. 1994.
Hypnum hexastichum Schwägr., Sp. Musc. Frond. Suppl. 1(2): 210. 1816. Isothecium
hexastichum (Schwägr.) Brid., Bryol. Univ. 2: 380. 1827. Pilotrichum
hexasticum (Schwägr.) C. Müll., Bot. Zeitung (Berlin) 6: 767. 1848. Neckera hexasticha
(Schwägr.) C. Müll., Syn. Musc. Frond. 2: 126. 1850. Meteorium hexastichum
(Schwägr.) Mitt., J. Linn. Soc., Bot. 12: 432. 1869. Pilotrichella
hexasticha (Schwägr.) Jaeger, Ber. Thätigk. St. Gallischen Naturwiss. Ges.
1875–76: 257. 1877. Protologue: Dominican
Republic. In Domingo insula lectum
communicavere cl. Thouin et Hornemann. Holotype: Hypnum hexastichum
Domingo Thouin. Diund [?]. Hornemann. (G).
Pilotrichella tenella Jaeg., Ber. Thätigk. St. Gallischen Naturwiss. Ges.
1875–76: 261 1877, new name for Neckera tenella C. Müll., Bot. Zeitung (Berlin) 17: 238. 1859,
not Neckera tenella Schwägr., 1827., illegitimate homonym. Protologue:
Insula St. Domingo. A bryologo Tönder suecico Lund. Mohr accepti. Lectotype (designated
here): Leskea n. sp. tenella ex St domingoa Mohr &
Weber [Herb. Swartzii] (S).
Plants slender to medium-sized, dull, light green,
yellow-green, reddish yellow or brownish green, in loose mats often with
pendent strands. Primary stems red, creeping; leaves reduced, erect,
ovate-triangular, 0.4–1.0 mm long. Secondary stems from primary stems stipitate,
others horizontal, erect or pendent, frequently and irregularly branched,
branches at times with flagelliform tips, to 4–10(–25) cm long, yellowish
green, becoming dark red with age, in cross section with sclerodermis of 3–4
small, thick-walled, reddish orange cells, cortex cells enlarged, firm-walled,
pale yellow, red with age, central strand absent; paraphyllia absent,
pseudoparaphyllia absent, scale leaves present; axillary hairs 2–3 cells long,
basal cell quadrate to subquadrate, reddish brown, upper 1–2 cells broadly
cylindrical, hyaline (occasionally reddish); rhizoids on primary stems, at base
of stipitate secondary stems, or on flagelliform branch tips, red, not or
irregularly branched, from clusters of initials abaxial to the leaf insertion.
Secondary stem and branch apices turgid; stem and branch leaves differentiated
only in size, obscurely seriately ranked when dry, distinctly so when wet,
leaves panduriform, broadly oblong, appressed, more or less clasping below,
broadly obovate, abruptly flexed and spreading above, 0.5–1.2 mm long, at times
somewhat auriculate, apex acuminate, long-cuspidate; margins broadly incurved,
sharply serrulate above, plane or broadly incurved, serruate or entire below;
costae short double, short single, or absent; leaf cells long flexuose, smooth
to slightly porose, 16–37 x 3–4 μm, outer basal cells shorter and broader,
inner basal cells elongate, porose, alar cells subquadrate to short-rectangular
extending up the margins in 2–4 rows and across the insertion in 4–6 rows,
yellow across the insertion. Dioicous. Perigonia gemmate, lateral on secondary
stems and branches, 0.8–1.0 mm long, perigonial leaves ovate, acuminate, costa
absent; paraphyses present, antheridia few. Perichaetia lateral, 2.5 mm long,
paraphyses and archegonia numerous; perichaetial leaves lanceolate 1.8–2.2 mm
long, vaginula densely hairy. Setae short, wavy, 1.5–2.4 mm long, smooth,
yellow. Capsules exserted, erect to somewhat inclined, ovoid to
short-cylindrical, 1.0–1.5 mm long, neck moderately developed; exothecial cells
small, rounded in 2–3 rows below the mouth, subquadrate, irregularly
short-rectangular, or oblate, firm-walled below, cells in neck smaller than the
median cells; stomata superficial on neck; opercula obliquely long-rostrate,
1.0 mm long; annulus not seen; peristome yellowish white, exostome teeth
linear, to 0.5 mm long, dorsal (outer) surface lightly horizontally striate at
base, smooth or finely papillose above, trabeculae weakly developed on both
sides, endostome nearly as long as exostome, basal membrane low, segments filamentous,
narrowly perforated, smooth to papillose, cilia rudimentary or absent. Spores
round, lightly roughened, 14–20 μm long. Calyptrae narrowly cucullate,
with a few long hairs.
Etymology.
The specific epithet hexasticha combines the Greek hexa- (six)
and -sticha (in a row) in reference to its seriately ranked leaves.
Distribution.
Caribbean (Cuba, Haiti, Dominican Republic).
Illustrations. Duarte-Bello (1997, Pl. 203); Buck
(1998, Pl. 91 1–8).
 Orthostichella hexasticha A. Habit. B. Median leaf cells. C.
Leaf apex. D. branch leaf. E. basal leaf cells. F. Stem leaf.
Orthostichella hexasticha A. Habit. B. Median leaf cells. C.
Leaf apex. D. branch leaf. E. basal leaf cells. F. Stem leaf.
 Orthostichella hexasticha A. Calyptra. B. Sporophyte. C. Operculum. D.
Exothecial cells showing stomata. E. Exothecial Cells. F. Axillary hairs. G. Stem
cross-section.
Orthostichella hexasticha A. Calyptra. B. Sporophyte. C. Operculum. D.
Exothecial cells showing stomata. E. Exothecial Cells. F. Axillary hairs. G. Stem
cross-section.
Ecology.
On rocks (laterite), tree trunks, branches, twigs, shrubs, lianas, and rotting
logs in pine forests, cloud forests, and roadbanks; 700–2000.
Selected
specimens examined. CUBA.
Guantánamo: Gradstein, Bryophyta Neotropica 370 (MO); Gramma: Taylor 528 (NY); Sancti Spíritus: Buck
23441 (NY); Santiago de Cuba: Leon 11228 (FH, NY);
Villa Clara: Pócs & Borhidi 9011/AH (MO). HAITI. Ouest: Imshaug 23071
(MO); Sud: Duncan
37 (MO); Sud’est: Buck 9395 (H, NY). DOMINICAN REPUBLIC. Azua: Mejía
& Zanoni 8270 (MO, NY); Barahona: Abbott 1789 (B, BM, FH, NY); Independencia: Bolay
41 (MO); La Estrelleta: Reese 15410 (NY); La Vega: Zanoni et al.
39270 (NY); Pedernales: Steere 22830 (NY); San Rafael: Norris et al. 6804 (NY).
The
most distinctive feature of O. hexasticha is its leaf shape. The
strongly panduriform leaves are erect, clasping at base, then abruptly reflexed
outward, and finally curved inward toward the stem. The upper portion of the
leaves is strongly concave with broadly incurved, sharply serrulate upper
margins that nearly meet below apex. The basal leaf margins are rounded to the
insertion or somewhat auriculate, often broadly incurved, entire or serrulate
at base. This leaf shape gives the plants an overall aspect that is helpful in
recognizing the species. Unfortunately there are numerous collections
(including the type of O. hexasticha) that consist of small plants that
have this apect weakly developed (see e.g., Norris B6612 MO, NY).
Furthermore, these collections have leaves that are distinctly ranked when dry.
They must be examined with a compound microscope before they can be positively
identified. In these collections of O. hexasticha the presence of
panduriform leaves that are not or moderately auriculate at base will separate
them from all other Neotropical members of the genus. The branches and stems of
O. hexasticha frequently end in flagelliform tips.
Squamidium
nigricans (Hook.) Broth. is similar
in size and aspect to O. hexasticha. Plants of S. nigricans
usually have some parts with an intense blackish color and this feature can be
used to separate it from O. hexasticha which never has this color. In
addition, S. nigricans never has flagelliform branch and stem tips.
Microscopically, the presence in the leaves of a long, faint single costa, and
greatly enlarged, strongly differentiated alar cells distinguished S.
nigricans from O. hexasticha.
Orthostichella
hexasticha is found in Cuba, Hispaniola, and Puerto
Rico. The Puerto Rican collections of O. hexasticha
represent the small form noted above. Buck (1998) considered these collections
to be O. versicolor.
3.
Orthostichella longinervis (Ren. & Card.)
Allen & Magill, comb. nov.
Pilotrichella loninervis Ren. & Card., Bull. Soc. Roy. Bot. Beligique
32(2): 23. 1893. Orthostichopsis longinervis (Ren. & Card.) Broth.,
Nat. Pflanzenfam. 1(3): 805. 1906. Protologue: Madagascar: ad truncos; Diego
Suarez (Chenagon); in silvis ditionis Antsianaka (fratres Perrot);
inter Vinanintelo et Ikongo (Dr Besson).
Plants
slender to medium-sized, dull, light green, yellow-green, reddish yellow or
brownish green, in loose mats at times with pendent strands. Stolons yellow to
red, creeping; leaves widely spaced, imbricate, narrowly ovate, abruptly
cuspidate, to 1.5 mm long; costa single, long, extending into the leaf apex;
alar cells moderately, dark-red, quadrate,. Primary stems at times strongly
stipitate or foliose to the stolons, horizontal, erect or pendent, 2–12 cm
long, frequently and irregularly branched, branches often with flagelliform
tips, yellowish green, becoming dark red with age, in cross section with
sclerodermis of 2–5 small, thick-walled, reddish orange cells, cortex cells
enlarged, firm-walled, pale yellow, red with age, central strand absent;
paraphyllia absent, pseudoparaphyllia absent, scale leaves present; axillary
hairs 3–4 cells long, basal 1–2 cells quadrate to subquadrate, reddish brown,
upper 2–3 cells broadly cylindrical, hyaline or reddish; rhizoids consistently
on stolons, also at base of stipitate secondary stems, red, not or irregularly
branched, from clusters of initials abaxial to the leaf insertions. Stipe
leaves broadly ovate, abruptly cuspidate, rounded at base; costae long, single,
reaching into the apex; alar cells moderately developed. Secondary stem and
branch apices turgid; stem and branch leaves differentiated only in size, stem
leaves not or obscurely seriately ranked when dry, branch leaves often ranked,
dry or wet, leaves obovate, oblong-obovate, to panduriform, erect to
erect-spreading, variously clasping below, 0.8–1.3 mm long, rounded to the
insertion, apex shorlty cuspidate; margins broadly incurved and subentire to
serrulate above, plane or broadly incurved, serrulate or entire below; costae
consistenly long, single, usually subpercurrent, at times wavy, often with
short lateral spurs; leaf cells long flexuose, smooth, 20–45 x 3–5, outer basal
cells shorter and broader, inner basal cells near insertion elongate, often
porose, alar cells red to red-yellow, subquadrate to short-rectangular
extending up the margins in 2–4 rows and across the insertion in 3–6 rows.
Dioicous. Perigonia gemmate, lateral on secondary stems and branches, 1 mm
long, perigonial leaves ovate, acuminate, costa absent; paraphyses present,
antheridia numerous. Perichaetia lateral, 1.2-1.8 mm long, paraphyses and
archegonia numerous; perichaetial leaves lanceolate, vaginula lightly hairy.
Setae short, wavy, 2–3 mm long, smooth below, roughened-papillose above,
yellow, becoming reddish with age. Capsules exserted, erect to somewhat
inclined, ovoid to short-cylindrical, 1.5 mm long, neck moderately developed;
exothecial cells small, rounded in 2–3 rows below the mouth, subquadrate,
irregularly short-rectangular, or oblate, firm-walled below, cells in neck
smaller than the median cells; stomata superficial on neck; operculum and
annulus not seen; peristome yellowish red, exostome teeth linear, 0.2–0.4 mm
long, dorsal (outer) surface horizontally papillose-striate at base, papillose
above, trabeculae weakly developed on both sides, endostome nearly as long as
exostome, basal membrane very low, segments filamentous, narrowly perforated,
not keeled, smooth to papillose, cilia rudimentary or absent. Spores round,
lightly roughened, 14–18 μm long. Calyptra not seen.
Etymology.
The specific epithet longinervis combines the Latin longus- (long)
with nervus (nerve) and refers to the long costa found in the leaf of
the species.
Distribution.
Western Indian Ocean.
Illustrations.
Renauld and Cardot (1895–1905, Pl. 83 1).
Ecology.
On tree branches; 800–1050 m.
Specimens
examined.
MADAGASCAR.
Antananarivo: Crosby & Crosby 5190 (MO); Antseranana: Magill et
al. 9947 (MO); Toamasina: Magill et al. 9664 (BM, FH, H, MO, NY, S),
Crosby & Crosby 9319 (MO).
Orthostichella
longinervis is a medium-sized species
that consistently has extremely well developed stipitate stems. There are some
collections of O. panduraefolia with strongly stipitate stems,
and while the feature occurs here and there among the other Orthostichella species,
it is never as prominent as found in O. longinervis. The critical
feature of O. longinervis, however, is its costal structure. The costa
is consistently expressed, long, typically subpercurrent, strongly developed,
and also often has short lateral spurs. This feature makes the species unique
in the genus, which typically has an extremely variable costa: double, single,
or absent on leaves from a single plant. Indeed, these two features of O.
longinervis are so odd within the context of Orthostichella that
they raise questions as to whether the species is properly placed there.
Athough
the gametophytes of Orthostichella can be extremely variable the same
can not be said for its sporophytes. Indeed, within all of its species, the
features of the Orthostichella sporophyte are remarkable stable. In this
regard it seems important that the O. longinervis sporophyte is also
identical in form to that of the other species. Furthermore, in terms of stem
morphology, leaf shape, margin stance and serration, areolation, and alar cell
development there is nothing unusual in gametophytes of O. longinervis
to indicate it is misplaced in Orthostichella.
Renauld
and Cardot (1883) considered O. longinervis to closely approach the
aspect of Porotrichum, but to differ, among other ways, in having
concave, subcocheariform, imbricate branch leaves in distinct seriate rows.
They also regarded the presence of a long, single costa in O. longinervis
as evidence that it might only be a form of Pilotrichella subimbricata or
P. chrysoneura, both of which are now placed in Orthostichopsis.
This latter view of Orthostichella longinervis was adopted by Brotherus
(1906) when he transferred the species into Orthostichopis. Although the
costa form in the leaves of Orthostichella longinerivis is identical to
that found in Orthostichopsis, the taxon is clearly misplaced within Orthostichopsis
because it has foliose rather than filamentous pseudoparaphyllia. The
presence of filamentous pseudoparaphyllia is a sine qua non feature not
only of Orthostichopsis but also the Pterobryaceae.
Although
Orthostichella longinervis differs significantly from Porotrichum
in having concave, spirally ranked leaves, there are other genera in the
Neckeraceae that lack complanate leaves. In particular Porotrichodendron
has one one species (P. lindigii (Hampe) Buck) with tumid, evenly
foliate leaves. Evidence against the placement of O. longinervis in Porotrichodendron
can be found in its lack of a stem central strand, as well as the very close
similarity of its sporophyte to those of other Orthostichella species.
On balance, this species appears best placed in Orthostichella where it
occupies a critical position in supporting the placement of the genus within
the Neckeraceae.
4.
Orthostichella
pandurifolia (C. Müll.) Buck,
Bryologist 97: 435. 1994.
Neckera
panduraefoliaC, Müll., Bot. Zeitung (Berlin) 13: 767. 1855.
Pilotrichella pandurifolia (C.Müller) Jaeger, Ber. Thätigk. St. Gallischen
Naturwiss. Ges. 1875–76: 255. 1877. Protologue: South Africa. Promontor. bonae
spei: Zeyher copiose sterilem arboream anno 1823 legit. Type: South
Africa. Prom. b. spei: Zeyher 1823 (isotype NY).
Neckera
decolorans Hampe
in C. Müll., Linnaea 40: 265. 1876, invalid name listed in synonymy. Pilotrichella
decolorans C.Müller in Kindberg, Enum. Bryin. Exot., suppl. 2 102
1891, invalid name, lacks a description. Based on: Insula Johana, Comoros, leg. Hildebrandt [Herb.
Hampe] (BM).
Pilotrichella kuntzei
C. Müll., Hedwigia 38: 127. 1889. Protologue: Habitatio. Prom. bonae spei,
Kingwilliamstown, “im Perie-Walde” ad Polypodium Eckloni: Dr. Otto
Kuntze le. et. mis. 1894, synomized by Magill & Rooy (1998).
Orthostichella
capillicaulis C. Müll., Flora 73: 491. 1890, invalid name, genus
invalid at time of publication. Pilotrichella capillicaulis C. Müll ex
Kindb. Enum. Bryin. Exot., suppl. 2:102 1891, new name for Orthostichella
capillicaulis C. Müll. Protologue: Tanzania. Africa or. trop., Leikipia in
occidente montis Kenia, ad pedem der Aberdare-Kette: L. Höhnel Novembri
1887 in Exped. Telekiana. Type: Tanzania. Afr. or. Kenia, leg. L. Höhnel
(H, isotype).
Orthostichella
curvifrons
C. Müll., Flora 73: 491. 1890, invalid name, genus invalid at time of publication.
Pilotrichella curvifrons C. Müll ex Kindb., Enum. Bryin. Exot.,
suppl. 2: 102 1891. new name for Orthostichella curvifrons C. Müll.,
Protologue: Tanzania. Africa or. trop., Leikipia in regione meridionali montis
Kenia, ad pedem der Aberdare-Kette: L. Höhnel Novembri 1887 in Exped.
Telekiana. Type: Tanzania. Afr. or. Leikipia, leg. L. Höhnel (H,
isotype).
Orthostichella
tenellula C.
Müll., Flora 73: 492. 1890, invalid name, genus invalid at time of publication.
Pilotrichella tenellula C. Müll. ex Kindb., Enum. Bryin. Exot.,
Suppl. 2: 102. 1891, new new for Orthostichella tenellula C. Müll.
Protologue: Patria. Africa or. trop., in sylva primaeva montis Kilima-Ndscharo:
L. Höhnel in Exped. Telekiana 1887. Type: Tanzania. Afr. or.,
Kilima-Ndscharo, leg. Höhnel (H, isotype).
Pilotrichella cuspidata Broth., Bot. Jahrb. Syst. 24: 255. 1897. Protologue:
Pondoland: in feuchtem Laubgebüsch an dem Unterholz an allen Zweigen und an den
Bäumchen rankend und wie Bartmoos herabhängend nur in einem Gebüsch des
Egorawaldes gefunden, daselbst aber eine grosse Strecke des Gebüsches damit
behangen (Beyrich n. 38, Bachmann n. 6). Types: Pondoland, Bachmann no
6 (BM, lectotype, designated here); Pondoland, Beyrich 38 (BM,
syntype).
Pilotrichella incurva Broth., Bot. Jahrb. Syst. 24: 256. 1897. Protologue:
Angola: Huilla (Antunes). Type. Afr. occ., Huilla, leg. Antunes
comm. Brotherus (S, isotype).
Pilotrichella stuhlmanni Broth., Bot. Jahrb. Syst. 24: 256. 1897. Protologue:
Seengebiet: Bukoba (Stuhlmann), Mau, an Bäumen c 2100 m (Scott Elliot
n. 73). Types: Bukoba, Stuhlmann (S, syntype); Bukoba, Stuhlmann 1890,
(S, syntype).
Pilotrichella
conferta
Ren. & Card., Bull. Soc. Roy. Bot. Beligique 38(1): 24. 1899 [1900].
Protologue: Lesotho. Hab. Africa australis: Lessouto (Vernet, herb.
Boissier). Type. Africa australis: Lessouto, Leg. Vernet, 1879 (PC,
holotype; BM, S, isotypes).
Orthostichella
leptopteridea C. Müll. in Par., Index Bryol. Suppl. 271. 1900,
invalid name, lacks a description. Pilotrichella leptopteridea Par.,
Index Bryol. Suppl. 271. 1900, invalid name, lacks a description, Based on:
Africa trop. Schoah, in viciniis urbis Ankober legit et in ventre Horpensis Museo. zool. Florentino misit 1887 (Stopfmaterial), cl. D.
Traversi, Hb. E. Levier (S)
Pilotrichella
percordata Broth
in P. Vard., Rev. Bryol. 47: 53. 1920, invalid name, lacks a
description. Based on: Kikouyou, British East Africa, 1910. A. G. Allan 315,
ex herb. Rev. D. Lillie (H); . Kikouyou, British East Africa, 1910. A. G.
Allan, ex herb. Rev. D. Lillie, [Herb. Potier de la Varde], (PC).
Pilotrichella
cordata
Brotherus in Potier de la Varde, Bull. Soc. Bot. France 71: 1057. 1924.
Protologue: [Tanganyka ?] Kiswani (leg. R.P. Wetzler) Invalid name,
lacks a description. Based on Kiswani, (Mambra) Tanganyka Territory, Juni 1924,
coll: R. P. Wetzler, comm. R. P. Soul (PC); Usumbara, Kiswani, leg. R.
P. Soul (FH, H).
Plants
slender to medium-sized, dull, light green, yellow-green, reddish yellow or
brownish green, in loose mats often with pendent strands. Stolons yellow to
red, creeping; leaves widely spaced, reduced, erect, ovate-triangular, abruptly
acuminate, to 1.6 mm long; ecostate or costa short and double; alar cells well
developed, dark-red, quadrate,. Primary stems at times stipitate or foliose to
the stolons, horizontal, erect or pendent, frequently and irregularly branched,
branches often with flagelliform tips, 2–15 cm long, yellowish green, becoming
dark red with age, in cross section with sclerodermis of 2–5 small,
thick-walled, reddish orange cells, cortex cells enlarged, firm-walled, pale
yellow, red with age, central strand absent; paraphyllia absent,
pseudoparaphyllia absent, scale leaves present; axillary hairs 3–4 cells long,
basal 1–2 cells quadrate to subquadrate, reddish brown, upper 2–3 cells broadly
cylindrical, hyaline or reddish; rhizoids consistently on stolons, also at base
of stipitate secondary stems, red, not or irregularly branched, from clusters
of initials abaxial to the leaf insertions. Stipe leaves ovate below,
piliferous above, auriculate to strongly rounded at base; costae short and
double; alar cells well developed. Secondary stem and branch apices turgid;
stem and branch leaves differentiated only in size, stem leaves not or
obscurely seriately ranked when dry, branch leaves often distinctly seriately
ranked, dry or wet, leaves obovate to oblong-obovate, erect to erect-spreading,
variously clasping below, 1.0–2.0 mm long, at times somewhat auriculate, apex
long cuspidate, branch leaf cuspid 140-200 µm long; margins broadly incurved and
subentire to serrulate above, plane or broadly incurved, serrulate or entire
below; costae usually short double or short single, occasionally absent; leaf
cells long flexuose, smooth, upper cells 20–46 x 4–6, median cells 30–60 x 3–4
μm, outer basal cells shorter and broader, inner basal cells near
insertion elongate, often porose, alar cells red to red-yellow, subquadrate to
short-rectangular extending up the margins in 2–4 rows and across the insertion
in 4–6 rows, yellow across the insertion. Asexual reproduction by deciduous
leaves. Dioicous. Perigonia gemmate, lateral on secondary stems and branches,
0.8–1.2 mm long, perigonial leaves ovate, acuminate, costa absent; paraphyses
present, antheridia numerous. Perichaetia lateral, 1.4-2.0 mm long, paraphyses
and archegonia numerous; perichaetial leaves lanceolate 1.4–2.0 mm long,
vaginula densely hairy. Setae short, wavy, 1.5–3.5 mm long, smooth below,
roughened-papillose above, yellow, becoming yellowish red with age. Capsules
exserted, erect to somewhat inclined, ovoid to short-cylindrical, 1.0–2.0 mm
long, neck moderately developed; exothecial cells small, rounded in 2–3 rows
below the mouth, subquadrate, irregularly short-rectangular, or oblate,
firm-walled below, cells in neck smaller than the median cells; stomata
superficial on neck; opercula obliquely long-rostrate, 1.0–1.5 mm long; annuli
rudimentary, consisting of 2–3 rows of thin-walled enlarged cells; peristome
yellowish white, exostome teeth linear, 0.3–0.4 mm long, dorsal (outer) surface
horizontally striate at base, smooth or finely papillose above, trabeculae
weakly developed on both sides, endostome nearly as long as exostome, basal
membrane very low, segments filamentous, narrowly perforated, not keeled,
smooth to papillose, cilia rudimentary or absent. Spores round, lightly
roughened, 14–20 μm long. Calyptrae, 2–3 mm long,narrowly cucullate,
lightly hairy.
Etymology.
The specific epithet panduraefolia combines the Latin panduratus
(fiddle-shaped) with folium (leaf).
Distribution.
Northeast, East, and South Tropical Africa, Southern Africa, Western Indian
Ocean.
Illustrations.
Magill and Rooy (1998, Fig. 160 1–11).
Ecology.
On tree trunks and branches, also fairly common on rocks; 100–1800 m.
Selected
specimens examined.
DJIBOUTI.
leg. unknown, 1906 (NY, S).
KENYA.
Central: Magill 13892 (BM, H, MO, NY, S).
TANZANIA.
Arusha: Pocs & Pocs 6213 (B).
ANGOLA: Huilla Antunes (S).
LESOTHO Vernet
(S).
SWAZILAND.
Hhohho: Magill 3497 (MO).
SOUTH
AFRICA. Eastern Cape: Perold 4142 (MO); Kwazula-Natal: Magill 5173
(H, MO); Limpopo: Magill 6543 (H, MO); Mpumalanga: Koekemoer 2219
(MO).
COMORO
ISLANDS. Johana Island: Hildebrandt (BM).
Orthostichella
pandurifolia has creeping primary
stems from which the secondary stems can be erect or pendent. The secondary
stems can be distinctly long-stipitate or evenly foliate throughout, with both
types of foliate secondary stems sometimes found on the same primary stem. Its
branches commonly end in slender attenuations. The branch leaves, and sometimes
the secondary stems, of O. pandurifolia are typically occur in distinct
spiral ranks while those on the stolons and primary stems are unranked. The
leaves are long-cuspidate, have incurved, serrulate upper leaf margins,
elongate, smooth, firm-walled leaf cells, and weakly developed, reddish yellow,
firm-walled alar cells. Most leaves in O. pandurifolia have a single or
double costae that sometimes reache well above midleaf, however, here and there
ecostate leaves can be found. This costal variation is opposite that of O.
versicolor in which most leaves are ecostate, and only occasionally do the
leaves have short double, short single, or long single costae. Sporophytes are
not often found in O. pandurifolia. The setae are short (1.5–3.5 mm
long), flexuose, and roughened above. The operculum is long, obliquely
rostrate, and the peristome is pale-yellow and greatly reduced. Distinctive
features of the peristome include its narrow, mostly papillose exostome teeth
that strongly cross-striate at the very base, and narrow endostomial segments
that are perforated but not keeled. The calyptrae are cucullate and sparsely
hairy and the vaginula are densely hairy.
There
are two major forms of O. pandurifolia. At the southern end of its
range, the species has very strongly ranked branch leaves, and plants with
deciduous leaves are unknown. In this form of the species sporophytes are
occasionally found. The second form of the species (nomenclaturally centered on
Pilotrichella stuhlmanni Broth.) is found in the middle and northern
parts of its range. The plants in this region have weakly ranked branch leaves,
and are nearly always found with deciduous leaves. None of the plants from this
region have been found with sporophytes. Orthostichella pandurifolia is
primarily an epiphytic species. There are however many collections that have
been collected on boulders, and unlike O. versicolor, the
saxicolous condition appears to be a primary rather than a secondary habitat
for O. pandurifolia.
Orthostichella
pandurifolia has sometimes been
confused with Squamidium brasiliense (Hornsch.) Broth because they both
are pendent mosses with with weaklyn developed costae. Squamidium
brasiliense differs from Orthostichella pandurifolia in having
julaceous stems, consistently long, single costae, and exceptionally well-developed
alar cells. Orthostichopsis species differ from Orthostichella
pandurifolia in having filamentous pseudoparaphyllia, long, strongly
developed, single costae, and cylindrical capsules.
Orthostichidium
pentastichum (Brid.) Allen and O.
involutifolium (Mitt.) Broth. are similar to Orthostichella pandurifolia
in having ecostate leaves with broadly incurved upper leaf margins and weakly
developed alar cells. Plants of O. pentasticum are more slender and less
branched than those of Orthostichella pandurifolia, while those of O.
involutifolium tend to be a little bit larger than those of Orthostichella
pandurifolia, and its leaves are more stiffly erect as well as more
strongly ranked and they have filamentous pseudoparaphyllia. Furthermore, both Orthostichidium
species have filamentous pseudoparaphyllia, and O. involutifolium often
has clusters of uniseriate gemmae around their branch primordia. Orthostichella
pandurifolia never has asexual gemmae, and the branch primordia are
surrounded by scale leaves.
5.
Orthostichella
roseana Allen & Magill, species
nova.
Type:
Ecuador. Vicinity of Huigra, mostly on the Hacienda de Licay, October 1918, J.
N. Rose 2400 (holotype NY; isotypes B, FH, MO, NY, S).
Species
haec a O. hexasticha inter alia foliis caulium auriculatis setaeque
longioribus differt.
Plants slender, dull, light green, yellow-green,
reddish yellow or brownish green, in loose mats often with pendent strands.
Stolons red, creeping; leaves widely spaced, reduced, erect, broadly ovate,
abruptly pointed, ecostate, 0.6-0-8 mm long. Primary stems stipitate,
horizontal, erect or pendent, frequently and irregularly branched, branches
frequently and irregularly branched, at times with flagelliform tips, to 10 cm
long, yellowish green, becoming dark red with age, in cross section with
sclerodermis of 3–5 small, thick-walled, reddish orange cells, cortex cells
enlarged, firm-walled, pale yellow, red with age, central strand absent;
paraphyllia absent, pseudoparaphyllia absent, scale leaves present; axillary
hairs 3–4 cells long, basal 1–2 cells quadrate to subquadrate, reddish brown,
upper 2–3 cells broadly cylindrical, hyaline or reddish; rhizoids only on
stolons and at the base of stipitate stems, red, not or irregularly branched,
from clusters of initials abaxial to the leaf insertions. Secondary stem and
branch apices turgid; stem and branch leaves differentiated only in size,
obscurely seriately ranked when dry, distinctly so when wet, leaves panduriform,
broadly oblong, appressed and clasping below, broadly obovate, abruptly flexed
and spreading above, stem leaves 1.0–1.2 mm long, strongly auriculate, apex
acuminate, long-cuspidate; margins broadly incurved, sharply serrulate above,
plane or broadly incurved, serrulate or entire below; costae short double,
short single, or absent; leaf cells long flexuose, smooth to slightly porose,
18–50 x 3–6 μm, outer basal cells shorter and broader, inner basal cells
elongate, porose, alar cells subquadrate to short-rectangular extending up the
margins in 2–4 rows and across the insertion in 2–4 rows, yellow across the
insertion. Dioicous. Perigonia gemmate, lateral on secondary stems 1.0 mm long,
perigonial leaves ovate, acuminate, costa absent; paraphyses present,
antheridia numerous. Perichaetia lateral, 2.0–2.5 mm long, paraphyses and
archegonia numerous; perichaetial leaves lanceolate 1.2–2.0 mm long, vaginula
hairy. Setae short, wavy, 3.5–4.0 mm long, smooth below, roughened above,
yellow. Capsules exserted, erect to somewhat inclined, ovoid to
short-cylindrical, 1.0–2.0 mm long, neck moderately developed; exothecial cells
small, rounded in 2–3 rows below the mouth, subquadrate, irregularly
short-rectangular, or oblate, firm-walled below, cells in neck smaller than the
median cells; stomata superficial on neck; opercula obliquely long-rostrate,
1.0–1.5 mm long; annulus not seen; peristome yellowish white, exostome teeth
linear, to 0.5 mm long, dorsal (outer) surface horizontally striate at base,
smooth or finely papillose above, trabeculae weakly developed on both sides,
endostome nearly as long as exostome, basal membrane very low, segments
filamentous, narrowly perforated, not keeled, smooth to papillose, cilia
rudimentary or absent. Spores round, lightly roughened, 10–14 μm long.
Immature calyptrae, 2 mm long, cucullate, lightly hairy.
Etymology.
The specific epithet roseana honors the collector of the type specimen
Joseph Nelson Rose (1862–1928) curator at the U.S. National Herbarium. Rose
collected plants in the Central and Southeastern United States, Mexico, and
South America. Stafleu & Cowen (1983).
Distribution.
Western South America.
Ecology.
Pendent from trees; 850–2500 m.
 Orthostechella roseana A. Habit. B. Branch leaf apex.
C. Stipe leaf. D. Leaf margin. E. Leaf apex. F. Stem cross-section. G-I. Branch
leaves. J. Basal leaf cells. K-O. leaves.
Orthostechella roseana A. Habit. B. Branch leaf apex.
C. Stipe leaf. D. Leaf margin. E. Leaf apex. F. Stem cross-section. G-I. Branch
leaves. J. Basal leaf cells. K-O. leaves.
Specimens
examined. ECUADOR. Azuay: Steyermark 52753 (FH, L, MO, NY); Cańar: Steere
& Balslev 25952 (NY); Chimborazo: Rose 24000 (B, FH, MO, NY, S),
Pallatanga, Spruce 1218 (NY); Loja: Laegaard 18475B (MO);
Pichincha: sin. coll. (FH, S). PERU. Cajamarca: Soukup 3908 (FH).
Orthostichella
roseana is a small species with well
spaced, panduriform stem and branch leaves. The stem and branch leaves are also
appressed, clasping below and abruptly flexed and spreading above. The stem
leaves are strongly auriculate at base. Stolons are difficult to find in the
few known collections of O. roseana, however, the species does appear to
be stipitate.
Morphologically
O. roseana occupies an intermediate position between O. hexasticha and
O. versicolor. In size and aspect it is nearly identical to O.
hexasticha, and both species have the same leaf stance. Orthostichella
hexasticha differs from O. roseana in having stem leaves that are
rounded to the insertion rather than strongly auriculate, and shorter (1.5–2.4
mm) setae. The sporophytes of O. roseana are identical to those of O.
versicolor. Furthermore, O. versicolor has two forms (the rigida-form
and pachygaster-form) that often have strongly auriculate stipe and,
less often, stem leaves. Most collections of O. versicolor are larger in
size than O. roseana, but there are several forms of O. versicolor that
are as small or smaller than O. roseana. Orthostichella roseana consistently
differs from O. versicolor in having well spaced, panduriform branch
leaves that are appressed and clasping at base.
Orthostichella
roseana is presently found in the
Adean regions of Ecuador and northern Peru. There is an especially confusing
form of O. versicolor from Bolivia that has the aspect of O. roseana.
It differs, however, in being somewhat larger in size and in having branch
leaves that are more or less spreading from the insertions.
6.
Orthostichella versicolor (C. Müll.) Allen &
Buck, Mem. New York Bot. Gard. 76(3): 140. 2003.
Neckera
versicolor C. Müll., Syn. Musc.
Frond. 2(1): 127. 1850. Orthostichella versicolor (C. Müll.) Hampe,
Vidensk. Meddel. Dansk Naturhist. Foren. Kjřbenhavn. ser. 3, 2: 277. 1870,
combination invalid. Pilotrichella versicolor (C. Müll.) Jaeg., Ber.
Thätigk. St. Gallischen Naturwiss. Ges. 1875–76: 258.1877. Protologue: Brasil.
Brasilia, Rio Janeiro: Beyrich, Beske. Lectotype (designated here):
Brasilia, Serra d’Estrella, Beyrich (BM); isolectotype: Serra d’Estrella
et Nov. Friburg, ad radices arborum, Beyrich (BM).
Neckera viridis
C. Müll., Syn. Musc. Frond. 2(1): 125. 1850. Pilotrichella viridis (C.
Müll.) Jaeg., Ber. Thätigk. St. Gallischen Naturwiss. Ges. 1875–76: 258. 1877. Orthostichella
viridis (C. Müll.) C. Müll., Linnaea 42: 493. 1879, combination invalid.
Protologue: Venezuela. Venezuela, Caracas: Moritz. Coll. I.
Thorreyana Hamburgensis No. 38. Lectotype (designated here): Carcaras, leg.
Moritz No. 38 (BM); isolectotypes: Caracas, Moritz 38 (L);
Venezuela, Caracas, Moritz, Müller in Hb. (NY).
Neckera rigida
C. Müll., Syn. Musc. Frond. 2(1): 126. 1850. Pilotrichella rigida (C.
Müll.) Besch., Mém. Soc. Sci. Nat. Cherbourg 16: 222. 1872. Protologue: Mexico.
Mexico: Miquel, Deppe.Lectotype (designated here): Mexiko. Deppe
(S); isolectotype: Mexico, Miquel (BM).
Neckera pachygaster C. Müll., Syn. Musc. Frond. 2(1): 126. 1850. Pilotrichella
pachygaster (C. Müll.) Jaeg., Ber. Thätigk. St. Gallischen Naturwiss. Ges.
1875–76: 256. 1877, syn nov. Protologue: Venezuela. Venezuela, in
montibus nivosis prov. Meridae: Moritz. Lectotype (designated here):
Caracas. leg. Moritz 171. Neckera pachygaster CM.,
Columbien (Merida 1844/45) Moritz (BM); isolectotypes: Columbien,
Merida, 1844/45, leg. Moritz 171 (H), Merida, Moritz 171 (L).
Neckera pachygaster C. Müll.var. gracilis C. Müll., Bot. Zeitung (Berlin) 15: 582
1857. Pilotrichella pachygaster var. gracilis (C.Müller) Paris, Index
Bryol. 948 1897. Protologue: Colombia. Nova Granada, S. Martha, Mina, 4000 ped.
alta: Funck et Schlim in Coll. Linden No. 1034. Lectotype: Nova Granada,
S. Martha, Mina, 4000 ped., leg. Funck & Schlim (BM). Probably
isolectotype: Voyage de L. Schlim. Nlle Grenade, prov. de
Santa Marta. Hauteur 4000 fts. Minca, 1846 ŕ 1852 fl. en Mai, No
1043 (L).
Pilotrichella mexicana Schimp. ex Besch., Mém. Soc. Sci. Nat.
Cherbourg 16: 223. 1872. Protologue: Mexico. Mejico (F. Müller, Schimper
comm.). Holotype: Herbarium Mexicanum, A°. 1853. Fred.
Müller. Comm. H. Schlumber. [Herb. Bescherelle] (BM); isotype: Orizaba, lg.
F. Müller (BM).
Pilotrichella pulchella W. P. Schimper ex Bescherelle, Mém. Soc. Sci.
Nat. Cherbourg 16: 222 1872. Protologue: Mexico. Orizaba (F. Müller,
Lorentz mihi comm.). Holotype: Mexico. Orizaba, 1853, legit F. Müller
[Herb. Bescherelle] (BM); isotype: Mexico, Orizaba, leg. Müller 1853
(H).
Neckera ampullacea Hampe ex C. Müll., Linnaea 40: 264. 1876. Pilotrichella
ampullacea (C. Müll.) Jaeger, Ber. Thätigk. St. Gallischen Naturwiss. Ges.
1875–76: 256. 1877. Protologue: Comoro Islands. Comoro – insula Johanna, ubi
primum legit Prof. Peters Berol. 1843; inter alios muscos misit J. M.
Hildebrandt 1875. Lecotype (designated here): Africa: Comoro – insula
Johańa. leg. Hildebrandt, 1875 (H).
Pilotrichella caldensis Ĺngstr., Ofvers. Forh. Kongl. Svenska Vetensk.-Akad.
33(4): 34 1876. Protologue: Brazil. Regnell n. 3 a misit. Type.
Brasilia, Rio de Janeiro, leg. A. F. Regnell 3a (Holotype S, isotypes H,
S).
Pilotrichella pachygastrella C.Müller ex Ĺngstr., Ofvers. Forh. Kongl. Svenska
Vetensk.-Akad. 33(4): 33. 1876. Protologue: Brazil Regnell No 3b et Henschen.
Lectotype (designated here): Brasilia, A. F. Regnell no 3b
(S); isolectotype: Caldas, Regnell no 3b (H,
S). Syntype: Caldas Brasiliae, S. Henschen (BM, S).
Neckera cyathipoma C. Müll., Linnaea 42: 414. 1879. Pilotrichella cyathipoma (C.
Müll.) Kindb., Enum. Bryin. Exot. 28. 1888. Protologue: Argentina. Argentinia
subtropica, Rio seco prope Sn. Andrés, cum N. Avellandae saepius
commixta, fertilis 19 Septbr. 1873. — Cuesta de Buyuyu, 15 Majo 1873, sterilis;
in alia cuesta Boliviae sterilis; Jujui; Cuesta de Sn. Diego, 12. Junio 1873
fertilis; Cuesta de Sn. Rosa; ubique rarius fructificans. Quoque in Cuesta
colorada inter Sn. Luis et Amareta Boliviae,14 Junio 1873. Types: Argentinia
subtropica, Rio seco prope St. Andrés, cum N. Avellandae saepius
commixta, 19 Septmb. 1873, P. G. Lorentz (lectotype designated here JE);
Argentinia subtropica, Rio seco prope San. Andrés, cum N. Avellandae
saepius commixta, 19 Sept. 1873, leg. P. G. Lorentz (isolectotype H);
Argentinia subtropica. Cuesta de Buyuyu. P. G. Lorentz (syntype FH, H).
Pilotrichella inflatifolia C. Müll., Flora 69: 282. 1886. Protologue: Săo Tomé. Bom Successo, 1050–1100 met. alta, ad arbores, [Adolf
Moller]. Types: V. afrika, S. Thomé: Moller, ded. C. Müller 1889 (S,
lectotype designated here); Ins. St. Thome (1100 m. alt). Legit A.
Moller (S, isotypes).
Pilotrichella
leptoclada
C.Müll., Flora 69: 282 1886. Protologue: Săo Tomé.
Encostas do Pico de S. Thomé, alt. 1500–2100 met., ad arbores, [Adolf Moller].
Types: Africa, S. Thomé: Moller, ded. C. Müller 1889, (S, lectotype
designated here); Vestafrica, S. Thomé, Moller 12, com. N. C. Kindberg,
ex hb. C. M. 1889, (H, isolectotype); Afr. occ. ins. S. Thomé, 1885, leg. A.
Moller (NY, isolectotype).
Pilotrichella guineensis Ĺngstr. ex C. Müll., Flora 69: 282. 1886.
Protologue: ex Guinea. Type: Guinea, ad Afzelius (S, lectotype
designated here).
Neckera moenkemeyeri C. Müll., Flora 69: 516. 1886. Pilotrichella moenkemeyeri (C.
Müll.) Kindb. Enum. Bryin. Exot. 102. 1891. Pilotrichella maenkemeyeri Kindb.
ex Par., Index Bryol. 952. 1897, orthographic variant. Protologue.
Nigeria. Africa occid. tropica, territorium fluminis Niger, Old-Calabar, ad
arborum truncos, 10 Novbr. 1883: W. Mönkemeyer. Types : Nigeria.
Niger-gebiet, Alt-Calabar ad arborum truncos. Fl. Afr. trop. occ. 11/10 1884,
leg. W. Mönkemeyer (JE, lectotype designated here, H, S, isolectotypes);
Nigeria. Niger fl. Alt Calabar, ad arbor. truncos, 1884, W. Mönkemeyer
(JE, isolectotype), syn. nov.
Pilotrichella obovata Kiaer in Wright, J. Bot. 26: 266. 1888, invalid
name lacks a description. Based on: Madagascar, Mt. Ankaratra (Borgen 28).
Musci Madagascarienses, Herb. Kiaer. In montibus Ankaratra 1877–1879 legit M.
Borgen No. 28 (L).
Orthostichella profusicaulis C. Müll., Flora 73: 493. 1890., invalid name. genus
invalid. Pilotrichella profusicaulis (C. Müll.) Par., Index Bryol. 948.
1897, combination invalid. Protologue: Patria Africa or. trop. Ugueno, in
regione meridionali montis Kilima-Ndscharo, 1500 m altitudinis: Dr. Hans Meyer
1889. Based on: Africa, Ugueno, Kilimandscharo, 1500 m, 1889, Hans Meyer,
com. C. Müller 1890 (S).
Pilotrichella grimaldii Ren. & Card., Bull. Soc. Roy. Belgique 30(2): 192.
1891 [1892]. Protologue: Madagascar: Diego Suarez (Chenagon).
Types: Madagascar, circa Diego Suarz, leg. Chenagon, Musci
Mascareno-Madagascarienses No 39, Herb. F. Renauld, (S,
isotype); Diego Suarez – Madagascar, leg. Cap. Chenagon (H, isotype);
Madagascar: Diego Suarez, leg. Chenagon 1890, Herb. F. Renauld, (S,
isotype).
Pilotrichella subpachygastrella Broth., Bih. Kongl. Svenska Vetensk.-Akad. Handl. 21
Afd. 3(3): 46. 1895. Protologue. Brazil. Prov. S. Paulo, ad arbores montis
Serra de Caracol (Mosén n. 428). E. Brasilia sine loco designato sub n.
24 misit amicissimus J. Cardot. Lecotype (designated here): Brasilia, prov. S.
Paulo, ad arbores montis “Serra de Caracol” 15/8/1873, Hj. Mosén (H);
isolectotype: Brazil. Prov. S. Paulo ad arbores montis Serra de Caracol. 1815/873.
H. Mosén 428 (S).
Pilotrichella communis C. Müll. ex Dusén, Kongl. Svenska Vetensk.
Acad. Handl. 28(2): 28. 1895. Orthostichella communis (Dusén) C. Müll. in
Paris, Index Bryol. 943. 1897, invalid name cited in synonymy. Protologue:
Cameroon. Habitat in Camerunia, ubi ad Isangille (Oran) pagum m. Martio a. 1892
c. fr. parum evolutis legi [Dusén]. Type: Africa occ. Camerunia.
Isangille pagum in ramis arboreus. Martii 1892, P. Dusén 651 (S,
holotype). Musci Africani in Camerunia a P. Dusén collecti 651.
Ad Isangille pagum in ramulis arborum die 23 m. Martii a 1892 (S, NY,
isotypes), syn. nov.
Pilotrichella gracilicaulis C. Müll. ex Dusén, Kongl.
Svenska Vetensk. Acad. Handl. 28(2): 30. 1895. Orthostichella gracilicaulis (Dusén)
C. Müll. in Paris, Index Bryol. 943. 1897, invalid name cited in
synonymy. Protologue: Cameroon. Habitat in Camerunia in ramulis arborum, ubi ad
Bomanam pagum c. 670 metra supra mare m. Julio a. 1892 c.fr. et inter Bibundi
pagum et Bomanam pagum c. 300 metra supra mare m. Decembri a. 1890 c.fr. et
Januario a 1892 c.fr. legi [Dusén]. Types. Musci Africani in Camerunia a
P. Dusén collect. In montibus Camerunensibus prope Bomanam pagum c. 600
[sic] metra supra mare in truncis arborum die 19 m. Julio a. 1892, Dusén 164
(S, lectotype designated here). Musci Africani in Camerunia a P. Dusén
collecti. Ad Bomanam pagum in ramulis. 19 July 1892 (S, syntype). Musci
Africani in Camerunia a P. Dusén collect. In montibus Camerunensibus
prope Bomanam pagum c. 620 [sic] metra supra mare in ramulis arborum m. Julio
a. 1892, Dusén 164 (S, syntype). Musci Africani in Camerunia a P.
Dusén collecti, Bibundi 5 January 1892 (S, syntype).
Pilotrichella turgidellacea C. Müll. ex Dusén, Kongl.
Svenska Vetensk. Acad. Handl. 28(2): 31. 1895. Orthostichella turgidellacea (C. Müll. ex Dusén) Buck, Bryologist 97: 435.
1994.
Protologue: Cameroon. Habitat in Camerunia in ramis ramulisque arborum , ubi ad
Bomanam pagum c. 670 metra supra mare m. Decembri a 1890 et supra Etome pagum
c. 800 metra supra mare m. Januario a. 1892 legi [Dusén]. Types. Musci Africani in Camerunia a P. Dusén
collecti. In montibus Camerunensibus ad Bomanam pagum c. 670 metra supra mare
in truncis arobrum die 18 m. Decembris a. 1890, [Dusén] 131, P
Duséns mossherbarium (S, lectotype designated here, S isolectotype); V. Afrika,
Kamrum, Bomana, 18/12/90, P. Dusén (S, isolectotype); Africa occ.,
Camerunia prope Etome pagum in ramulis, Januario a. 1892, P. Dusén (S,
syntype).
Orthostichella filamentosula C. Müll., Bull. Herb. Boissier 5: 204. 1897. Pilotrichella
filamentosula (C. Müll.) Par., Index Bryol. Suppl. 271. 1900. Protologue:
Guatemala. Sine loco speciali quam linteum involucris serviens (Packmaterial)
in Museo zoologico Florentino 1892: Hb. Levier. Types: e viciniis urbis
Guatemala. rudem accepit Mus. Zoolog. Florent. quam involucrum animalium, 1892,
legit indigenus (lectotype designated here BM); e viciniis urbis
Guatemala. rudem accepit cum pellibus animalium Mus. Zool. Florentinum, 1892, collector
ignotus (isolectotype NY, H).
Pilotrichella subimbricata (Hampe) Jaeg., var. flageyi Ren. & Card. In
Ren., Prodr. Fl. Bryol. Madagascar 196. 1898. Orthostichopsis subimbricata (Hampe)
Broth. var. flageyi (Ren. & Card.) Card., Hist.Phys. Madagascar,
Mousses 345. 1915. Protologue. Madagascar: Diego Suarez, Chenagon, 1890;
Andrangoloaka, Sikora, 1892. Type: Madagascar, Diego Suarez, leg. Chenangon
(S, lectotype, designated here), syn. nov.
Pilotrichella subpanduraefolia Par., Index Bryol. Suppl. 272. 1900, new name for Pilotrichella
panduraefolia C. Müll. ex Dusén, Kongl. Svenska Vetensk. Acad.
Handl. 28(2): 32. 1895, illegitimate homonym not Pilotrichella panduraefolia
(C. Müll.) Jaeg, 1877. Protologue: Habitat in Camerunia in truncis
ramulisque arborum, ubi in montibus Rumpi ad Tokko pagum c. 900 metra supra
mare m. Aprili a. 1892 et in montibus Camerunensibus c. 300 metra supra mare m.
Januario a. 1892 legi [Dusén]. Types. Musci Africani in Camerunia a P.
Dusén collecti. In montibus “Rumpi” ad Tokko pagum c. 900 metra supra mare in
truncis arborum die 16 m. Aprilis a. 1892 [Dusén] 859 (S,
lectotype; L, S isolectotypes); In montibus Camerunensibus c. 300 metra supra
Bibundi pagum, in ramulis arborum m. Januario a. 1892, P. Dusén (S,
syntypes).
Orthostichella microcarpa C. Müll., Hedwigia 40: 86. 1901. Pilotrichella
microcarpa (C.Müll.) Broth., Nat. Pflanzenfam. I(3): 811 1906. Protologue:
Brazil. Brasilia, Sa. Catharina, Serra Geral, in declivibus Serrae ad ramos
arborum., Febr. 1890: E. Ule, Coll. No. 866. Type: Brasilia, S.
Catharina, Serra Geral, auf Baum zweigen am abhang der Serra do Oratorio,
2/1890, leg. E. Ule 866 (lectotype designated here H).
Orthostichella mucronatula C. Müll., Hedwigia 40: 87. 1901. Pilotrichella
mucronatula (C. Müll.) Broth., Nat. Pflanzenfam. I(3): 811 1906. Protologue: Brazil. Brasilia,
Sa. Catharina, Tubarăo, in sylva ad ramulos arborum prope Conconhaz, Sept.
1889; E. Ule, Coll. No. 778; ad flumen Laranjeiras superius prope
Orleans in Sierra Geral, Sept. 1889; idem, Coll. No. 779; Nova Venezia,
ad truncos arborum sylvestrium, Julio 1891: idem, Coll 1169. Lectotype
designated here: Brasilia, prov. S. Catharina, Tubarăo, auf Baumzweigen in
Wäldehen bei Conconhaz, Sept. 1889, leg E. Ule 778 (H); syntypes: An
Bäumen am oberen Larenjeiras bei Orleans, Estado de Sta. Catharina: September
1889, E. Ule 779 (JE); Sta Catharina, in arboribus ad flumen
Laranjeiras prope Oreans in Serra Geral, leg. E. Ule 779 (JE); Brasilia,
prov. S. Catarina, Orleans auf Baumen am oberen Laranjeiras, Sept. 1889, leg E.
Ule 779 (H); An Baumstämmen im Walde bei Nova Venezia, Estado de Sta.
Catharina: Juli 1891, E. Ule 1169 (BM, S); Brasilia, prov. S. Catarina,
Nova Venezia, am Baumstämmen im Walde. Juli 1891, leg E. Ule 1169 (H).
Pilotrichella subpachygastrella var. minor Broth. in Bauer, Verh. Zool.-Bot. Ges. Wien 55:
579. 1905, invalid name, lacks a description. Based on: Brazil. Canóas, auf
Baumrinde in Wädern [Brazil, Porto Alegre], Ed. M. Reineck & Jos.
Czermak (B, BM, FH, JE, S).
Pilotrichella
attenuata
Broth., Denkschr. Kaiserl. Akad. Wiss., Math.- Naturwiss. Kl. 88: 739 1913.
Protologue: Tanzania. Deutsch-Osafrika: West -Usambara, Wald unterhalb
Mazumbei, an Bäumen, zirka 1200 m ü. M., 18./VIII. 1909, [Brunnthaler].
Holotype: West-Usambara, Wald unter Mazumbi, an Bäumen, 1200 m, 18/8 1909, leg.
J. Brunnthaler.
Pilotrichella
angustifolia Herz., Biblioth. Bot.
87: 112. 1916, syn nov.
Protologue:
Bolivia. Ad Baumästen in der Talschlucht bei Tablas, ca. 1800 m, [Herzog]
4666. Type: An Baumästen in der Talschlucht bei Tablas, ca. 1800 m., Mai
1911, leg. Th. Herzog 4666 (holotype JE; isotypes H, L, S).
Pilotrichella
cyathipoma var. laxiretis Herz.,
Biblioth. Bot. 87: 112. 1916, syn. nov.
Protologue:
An Bäumen im Wald des Randgebirges bei Yacuiba, ca. 500 m., [Herzog] No.
2626. Type: Bolivia. Im Wald des Randgebirges bei Yacuiba, ca. 500 m.,
Oktober 1910, Th. Herzog No. 2626 (holotype JE, isotypes H, L,
S).
Pilotrichella
allionii Broth., Rev. Bryol. 47: 14.
1920 [1921], syn nov.
Protologue:
Ecuador. Prov. de l'Oriente. Gualaquiza; in arboribus ad ripam fl. Bomboiza
prope ejus confluentiam cum fl. Gualaquiza; 800–900 m [M. Allioni].
Gualaquiza; in silva El Salado; 950 m [M. Allioni]. Types: Ecuador -
Oriente - V. Bomboiza. Ad arborum ramos in silva “El Salado” fregnens sed
permisita Gualaquiza m 950, 25 Aug. 1910, M. Allioni 497 (H, lectotype
designated here). Ecuador., prov. Azuay. Gualaquiza: ad ripam flum. Bomboiza
prope ejus confluentiam cum fl. Gualaquiza, in arboribus, Jul. 1909, legit Rev.
M. Allioni (H, syntype); V. Boimbizo, Ad truncos in sylva El .Salado, 950
m, 25 Viii 1910, M. Allioni (PC, isolectotype).
Pilotrichella rigens Broth., Denkschr. Kaiserl. Akad. Wiss., Math.- Naturwiss. Kl. 83: 309
1926, illegitimate homonym. Not Pilotrichella rigens Card., 1910.
Protologue: Brazil. Săo Paulo. Prope Raiz da Serra; ad arbores; 20–50 m s. m.
([Schiffner] 695 p.p.). Prope Salto Grande do Rio Paranapanema;
ad arbores; ca. 500 m s. m. ([Schiffner] 1716, 1718). In ripa
sinistra fluminis Paranapanema ad cataractas “Salto Grande”; ad arbores ([Schiffner]
1795). Prope “Fazenda Bella Vista” in districtu urbis S.Cruz ad flumen
Rio Pardo; ad ramulos; ca. 500 m s. m. ([Schiffner] 1937). In
itinere Cerqueira Cesar–Frazenda “Bella Vista”; ad arbores ad flumen Rio Turvo;
500 m s. m. ([Schiffner] 1183). Apud cataractas “Salto dos Treis
Ranjos” prope urbem Cerqueira Cesar; ad arbores; ca. 500 m s. m. ([Schiffner]
493). In silvaticis inter Faxina et Apiahy prope Lagoas ([Schiffner]
97). Apiahy, Catos Altas da Ribeira (Puiggari 1986). Ad viam
inter Apiahy et Yporanga (Puiggari 48). Syntypes: Prov. Paraná: In ripa
sinistra fluminis Paranapanema ad cataractas “Salto Grande”, ad arbores,
27.VII.1901, lgt. V. Schiffner 1795 (BM, H); Săo Paulo. Prope
“Fazenda Bella Vista” in districtu urbis S.Cruz ad flumen Rio Pardo, ca. 500 m
s. m., ad ramulos, 19.VII.1901, lgt. V. Schiffner 1937 (BM, H,
S); Săo Paulo. Apud cataractas “Salto dos Treis Ranjos” prope urbem
Cerqueira–Cesar. ca. 500 m s. m., ad arbores, 21.VII.1901, lgt. V. Schiffner
493 (BM, H, S); Săo Paulo. Prope Salto Grande do Rio Paranapanema. ca
500 m.s.m., ad arbores, 25.VII.1901, lgt. V. Schiffner 1716 (BM, H); Săo
Paulo. Prope Salto Grande do Rio Paranapanema; ad arbores; ca. 500 m s. m. 25.
VII. 1901, lgt. Schiffner 1718 (BM, H, S); Săo Paulo. In itinere
Cerqueira Cesar–Frazenda “Bella Vista”. Ad flumen Rio Turvo, ad arbores, 500 m
s. m., 22 VI 1901, lgt. V. Schiffner 1183, (BM, H, S); Săo Paulo,
Apiahy, chemin d’ Yporanga, 7/1880, leg. J. J. Puiggari 48 (H);
Brasilis, Săo Paulo, Apiahy, Catos Altas da Ribeira, 1/9/1881, leg. J. J.
Puiggari 1986 (H); Săo Paulo. In silvaticis inter Faxina et Apiahy. ca. 800
m.s.m. pr. Lagoas, 23.VIII.1901, lgt.. V. Schiffner 97 (H);
Brazilia. Prov. Săo Paulo. Prope Raiz da Serra; 20–50 m.s.m., ad arbores;
4.VI.1901, lgt. V. Schiffner 695 (BM, H).
Nomenclatural notes
1.
The protologue of Neckera moenkemeyeri C. Müll gives the year of the
type collection as 1883 while the types in H, JE, and S give the year of
collection as 1884. According to Vegter (1976), Mönkemeyer collected in
southern Nigeria and Fernando Po between 1884–1885.
2.
There is a specimen of in L that appears to be an isolectotype of Neckera
pachygaster var. gracilis C. Müll. The collection bears the orginal label
from the “Voyage de L. Schlim”, but differs from the protologue in two
respects: its collection number is No. 1043 rather than No. 1034,
and the town is given as Minca rather than Mina. The L collection is
considerably larger than the BM collection, and the plants in both collections
are identical.
3.
None of the potential type collections of P. gracilicaulis in S exactly
fit the protologue information for that species. There is one collection,
marked “TYPUS” that contains a label in Dusén’s handwriting which is also
marked in red “sp. nov!”, but this collection was made on 21 January 1891 at
700 meters. The only specimen clearly marked as from the Dusén herbarium
is chosen here as the lectotype.
Plants
slender to medium-sized, dull, light green, yellow-green, reddish yellow or
brownish green, in loose mats often with pendent strands. Stolons yellow to
red, creeping; leaves widely spaced, reduced, erect, ovate-triangular, abruptly
acuminate, ecostate, 1.0–1.4 mm long. Primary stems at times stipitate or
foliose to the stolons, horizontal, erect or pendent, frequently and
irregularly branched, branches at times with flagelliform tips, to 4–10(–40) cm
long, yellowish green, becoming dark red with age, in cross section with
sclerodermis of 3–5 small, thick-walled, reddish orange cells, cortex cells
enlarged, firm-walled, pale yellow, red with age, central strand absent;
paraphyllia absent, pseudoparaphyllia absent, scale leaves present; axillary
hairs 3–4 cells long, basal 1–2 cells quadrate to subquadrate, reddish brown,
upper 2–3 cells broadly cylindrical, hyaline or reddish; rhizoids consistently
on stolons, also at base of stipitate secondary stems, or on flagelliform
branch tips, red, not or irregularly branched, from clusters of initials
abaxial to the leaf insertions. Stipe leaves ovate, cordate, rounded at base or
straight to the insertions. Secondary stem and branch apices turgid; stem and
branch leaves differentiated only in size, stem leaves not or obscurely
seriately ranked when dry, branch leaves often distinctly seriately ranked, dry
or wet, leaves obovate to oblong-obovate, erect to erect-spreading, variously
clasping below, 1.0–1.6 mm long, at times somewhat auriculate, apex cuspidate;
margins broadly incurved and sharply serrulate above, plane or broadly
incurved, serrulate or entire below; costae mostly absent, occasionally short
double or short single; leaf cells long flexuose, smooth, upper cells 20–30 x
4–6, median cells 30–60 x 3–4 μm, outer basal cells shorter and broader,
inner basal cells near insertion elongate, often porose, alar cells red to
red-yellow, subquadrate to short-rectangular extending up the margins in 2–4
rows and across the insertion in 4–6 rows, yellow across the insertion. Asexual
reproduction by deciduous leaves common. Dioicous. Perigonia gemmate, lateral
on secondary stems and branches, 0.8–1.2 mm long, perigonial leaves ovate,
acuminate, costa absent; paraphyses present, antheridia numerous. Perichaetia
lateral, 1.4-2.0 mm long, paraphyses and archegonia numerous; perichaetial
leaves lanceolate 1.4–2.0 mm long, vaginula moderately hairy. Setae short,
wavy, 2.5–7.0 mm long, smooth below, roughened above, yellow, becoming
yellowish red with age. Capsules exserted, erect to somewhat inclined, ovoid to
short-cylindrical, 1.0–2.0 mm long, neck moderately developed; exothecial cells
small, rounded in 2–3 rows below the mouth, subquadrate, irregularly
short-rectangular, or oblate, firm-walled below, cells in neck smaller than the
median cells; stomata superficial on neck; opercula obliquely long-rostrate,
1.0–1.5 mm long; annuli rudimentary, consisting of 2–3 rows of thin-walled
enlarged cells; peristome yellowish white, exostome teeth linear, to 0.5 mm
long, dorsal (outer) surface lightly horizontally striate at base, smooth or
finely papillose above, trabeculae weakly developed on both sides, endostome
nearly as long as exostome, basal membrane very low, segments filamentous,
narrowly perforated, not keeled, smooth to papillose, cilia rudimentary or
absent. Spores round, lightly roughened, 14–20 μm long. Calyptrae, 2–3 mm
long,narrowly cucullate, smooth to lightly hairy.
Etymology.
The specific epithet vesicolor is a Latin adjective meaning “variously
colored.” It probably refers to the fact that plants of O. veriscolor
occur in various combinations of green, yellow, red, and brown.
Distribution.
Mexico; Central America; Caribbean; Western, Northern, and Southern South
America, Brazil; West and East Tropical Africa; Western Indian Ocean.
Illustrations.
Herzog (1916, Fig. 41 a–b); Bartram (1949, Fig. 118 A–C); Florschütz (1964,
Fig. 100); Churchill (1994, Fig. 27 C); Sharp et al.(1994, Fig. 537);
Churchill (1995, Fig. 124 a–g); Buck (1998, Pl. 91 9–17); Gradstein et
al. (2001, Fig. 156 A–I); Buck (2003, Fig. 126).
 Orthostichella versicolor A. Branch with sporpohyte. B. Stem leaf. C. Branch
leaf apex. D-F & K-L. Branch leaves. G. Median leaf cells. H. Peristome
teeth and segment. I. Sporophyte with operculum. J. Basal leaf cells. M. Stem
cross-section.
Orthostichella versicolor A. Branch with sporpohyte. B. Stem leaf. C. Branch
leaf apex. D-F & K-L. Branch leaves. G. Median leaf cells. H. Peristome
teeth and segment. I. Sporophyte with operculum. J. Basal leaf cells. M. Stem
cross-section.
Ecology.
On the trunks, branches, and twigs of trees, vines, shrubs, logs, leaves, and
tree fern fronds, occasionally on moist rocks along rivers and boulders along
roadsides and dry slopes; 50–2600 m.
Selected
specimens examined.
MEXICO.
Baja California Sur: Brandegee (NY); Chiapas: Laughlin 540 (MO);
Colima [Socorro Island]: Felger 15853 (MO); Hidalgo: Magill 2317
(MO); Jalisco: Mexia 1338a (FH, MO, NY); Morelos: Pringle 15670
(FH, US); Oaxaco: Richards et al. 3801b (MO, NY, US); Puebla: Sharp
1000 (MO, NY); San Luis Potosí: Sharp 5888 (MO, NY); Sinaloa: H.
S. Gentry 7283 (FH, NY); Tabasco: Ruiz 1459a (H); Tamaulipas: Manuel
333 (MO); Veracruz: Pringle 15147 (FH, MO, NY, US).
BELIZE.
Cayo: Allen 18428 (BRM, MO); Toledo: Allen 19057 (BRM, MO).
GUATEMALA. Alta Verapaz: Standley 91732 (FH, NY, S); Chimaltenango: Wenzel
& Mitchell s. n. (MO); Chiquimula: Steyermark 31558 (FH, NY);
Guatemala: Standley 80668 (FH, NY); Huehuetenango: Steyermark 49439 (FH);
Izabal: Steyermark 39991 (FH, MO); Jalapa: Steyermark 32533 (FH,
S); Petén: Lundell 2830 (NY, S); Quezaltenango: Standley 86702 (FH,
NY, US, S); Quiché: Aguilar 915 (FH, S); Sacatepéquez: Standley 88951
(FH, NY, S); San Marcos: Croat 40907 (MO, NY); Suchitepéquez: Steyermark
46666 (FH, MO, US). EL SALVADOR. Ahuachapán: Padilla s.n. (NY).
HONDURAS. Atlántida: Allen 17314 (MO, TEFH); Copán: Allen 17781
(MO); Cortés: Allen 14354 (MO, NY); Franciso Morazán: Allen 12389B
(MO); Lempira: Allen 12148 (MO); Olancho: Allen 12777B (MO);
Santa Bárbara: Crosby 2793 (MO); Yoro: Allen 13664 (MO).
NICARAGUA. Estelí: Moreno 19249 (H, MO, NY, US, S); Granada: Almedo
1459B (MO); Jinotega: Stevens 16695 (MO, NIC); Madriz: Stevens
16137 (MO, NIC); Matagalpa: Stevens 22193 (MO); Zelaya: Schramm (FH,
NY, US). COSTA RICA. Alajuela: Crosby & Crosby 5754 (BM, CR, G, H, NY,
US); Cartago: Liesner & Judziewicz 14486 (MO, NY); Guanacaste: Crosby
3851 (MO, S); Hererdia: Crosby & Crosby 5780 (BM, CR, F, FH, G,
H, JE, NY, S, US); Limón: Steere CR-57 (NY); Puntarenas: Lyon 511
(CR, MO); San José: Crosby & Crosby 5746 (BM, CR, F, FH, G, H, JE,
MICH, NY, PC, S, US). PANAMA. Bocas Del Toro: Allen 5844 (MO); Canal
Area: Bailey & Bailey 511 (NY); Chiriquí: Croat 15925 (MO);
Coclé: Witherspoon 8771 (MO); Colón: Crosby 10354 (MO); Darién: Allen
8718 (MO); Panamá: Crosby 4489 (MO); Veraguas: Knapp & Sytsma
2543 (MO).
CUBA.
Gramma: Ekman 5390 (FH); Pinar del Río: León 12731 (NY);
Sancti Spíritus: Clement 11 (NY); Santiago de Cuba: Hamilton (NY).
PUERTO RICO. Aguadilla: Stevens 1781 (NY); Arecibo: Britton &
Marble 1177 (MO, NY, S); Ponce: Britton & Marble 974 (NY).
DOMINICA. Elliot 639 (BM, FH).
COLOMBIA.
Antioquia: Callejas, Roldán & Científico 10182 (NY); Boyacá: Amórtegui
357 (FH); Caquetá: Churchill & Betancur 16896 (H, MO, NY);
Cauca: Churchill & Franco 16583 (H, MO, NY); Cundinamarca: Apollinaire
(B, BM, FH, JE, L, MO, NY, S); Magdalena: Mägdefrau 1159 (B, MO);
Narińo: Churchill & Rengifo 17599 (H, MO, NY, S); Risaralda: Betancur
et al. 3110 (NY); Santander: Churchill, Franco & Parra 18760 (NY);
Valle: Hartman 217 (MO); Vaupes: Schultes 11740 (FH). ECUADOR.
Carchi: Steere 8871 (NY); Imbabura: Steere 9262 (NY);
Morona-Santiago: Holm-Nielsen et al. 4632 (MO, S); Napo: Holm-Nielsen
& Jeppesen 760 (MO); Pastaza: Holm-Nielsen & Jeppesen 406
(MO); Pichincha: Fransén 87 (NY, S); Tungurahua: Spruce 1197 (BM,
NY); Zamora-Chinchipe: Ortega 544a (MO). PERU. Ayacucho: Cook &
Gilbert 919 (NY); Cuzco: Cook & Gilbert 919 (BM); Junin:
Gradstein, Bryophyta Neotropica 67 (B, H, L, MO, NY, S); San Martin: Frahm
et al. 1362 (B, H, NY); Tumbes: Simpson BR-137 (MO). BOLIVIA.
Chuquisaca: (Prov. Sud Cinti) Serrano 1318 (MO), (Prov. Calvo) Churchill
20693 (MO); Cochabamba: (Prov. Chapare) Churchill et al. 19857 (MO);
La Paz (Prov. Inquisivi) Lewis 89-1018 (MO); Santa Cruz: (Prov.
Caballero) Churchill et al. 21914 (MO), (Prov. Chiquitos) Lewis
85-1298 (US), (Prov. Florida) Churchill & Vásquez 21777 (MO),
(Prov. Ibanez) Fuentes s.n. (MO), (Prov. Ichilo) Nee 38863 (MO,
NY), (Prov. Vallegrande) Vargas 690 (MO); Tarija: (Prov. Arce) Lewis
84-2192 (US), (Prov. O’Connor) Lewis 79-532 (MO).
PARAGUAY.
Alto Paraná: Zardini & Guerrero 39949 (MO); Caazapá: Zardini
7850B (MO, NY); Canendiyú: Zardini &bFranco 55007A (H, JE, MO,
NY, US); Paraguarí: Hahn 746 (H, MO, NY, US); San Pedro: Zardini
& Guerrero 42771 (MO). ARGENTINA. Jujuy: Hosseus 27/VI/33 (JE);
Misiones: Hamilton s.n. (B, BM); Salta: Price et al. 1580 (MO);
Tucumán: Buck 25968 (NY). URUGUAY. Montevideo: sin. coll. (NY).
VENEZUELA.
Anzoátequi: Steyermark 61131 (FH); Aragua: Nee & Whalen 16883 (B,
NY); Barinas: Nee & Whalen 17084 (NY); Falcon: Griffin &
Wingfield PV-1648 (MO); Lara: Griffin & López 336 (B, MO);
Mérida: Fendler 114 (NY); Monagas: Pursell 8960 (MO); Portuguesa:
Steyermark et al. 126699 (MO); Táchira: Liesner & González 10923A
(MO); Trujillo: Liesner et al 13029 (MO) SURINAME. Commewijne: Schallert
(NY); Sipaliwini: Allen 25508 (B, BM, FH, H, JE, MO, NY, U, US). FRENCH
GUIANA. St. Laurent-du-Maroni: Buck 25580 (NY).
BRAZIL.
Federal District: Heringer 8303A (NY); Minas Gerais: Mexia 5358a
(FH, MO); Paraná: Dusen 8495 (NY); Rio de Janeiro: Gradstein Bryophya
Neotropica 218 (B, H, MO, NY, S); Rio Grande do Sul: Wasum et al. 2160
(MO, NY); Santa Catarina: Frahm 1652 (MO); Săo Paulo: Visnadi &
Vital 515 (MO).
NIGERIA.
Cross River: Mönkemeyer (JE).
GHANA.
Capt. Benton (BM).
KENYA.
Rift Valley: Bishop Harmington (BM); Kaibibich, Cherangani Hills,
1°12’N, 35°17 E, Friis 2772 (MO).
MALAWAI:
Southern: Chapman & Chapman 9471 (MO).
TANZANIA.
Arusha: Rojkowski 369 (B).
ZAIRE:
Kivu: Hendrickx 3891 (MO).
ZIMBABWE.
Inyanga: Müller 2646 (MO); Nyanga: Clarke 90 (MO).
COMORO
ISLANDS. Anjouan Island: Magill & Pócs 11241 (B, BM, F, G, H, MO,
NY, PC S, US); Grande Comore Island: Magill & Pócs 10924 (BM, L, FH,
G, H, JE, MO, NY, S, US); Mayotte Island: Magill & Pócs 11620 (JE,
MO, NY); Mohéli Island: Magill & Pócs 11729 (MO).
MADAGASCAR.
Antseranana: Magill et al. 9961 (H, MO, NY); Antananarivo: Dorr &
Rakotozafy 2806 (MO); Fianarantsoa: Crosby & Crosby 8642 (MO);
Toamasina: Dorr 3141 (BM, FH, JE, MO, S).
Orthostichella
veriscolor has a complex morphology
with creeping primary stems and erect or pendent (some collections reported to
hang 1.0–1.5 meters long) secondary stems. The secondary stems can be strongly
stipitate or evenly foliate throughout. Remarkably, stipitate and evenly foliate
secondary stems can sometimes be found on the same primary stem. Its branches
commonly end in filiform attenuations or stolons. These different structures
can be morphologically distinct, but more often they intergrade one into
another or they can be abruptly transformed from one structure to another. As a
rule the leaves on the branches, and sometimes the secondary stems, of O.
versicolor are variously spirally ranked while those on the stolons and
primary stems are unranked. But, this rule is often violated. Most leaves are
cuspidate, have distinctive incurved, serrulate upper leaf margins, elongate,
smooth, firm-walled leaf cells, and weakly developed, reddish yellow,
firm-walled alar cells. The costa in O. versicolor is one of its oddest
features. Most leaves are ecostate, however, here and there leaves can be found
with short double, short single, or long single costae. Sporophytes are rarely
encountered in this dioicous moss. The setae are 2–6 mm long, flexuose, and
roughened above. The operculum is long, obliquely rostrate, and the peristome
is pale-yellow and greatly reduced. Distinctive features of the peristome
include its narrow, mostly papillose exostome teeth that weakly cross-striate
at the very base, and narrow endostomial segments that are perforated but not
keeled. The calyptrae are cucullate and either naked or sparsely hairy.
There
are three major forms of O. veriscolor in the Neotropics. The common
form of the species in the northern part of its range is nomenclaturally
centered on Neckera rigida C. Müll. (Mexico) and Neckera pachgaster C.
Müll. (Venezuela). In this form the plants are often turgid, distinctly
stipitate, and the leaves are weakly ranked. The secondary stem leaves are
strongly auriculate and deciduous branch leaves are not generally encountered.
Throughout the southern part of its range the common form of the species has
more slender plants that may or may not be stipitate. The secondary stem leaves
are not or weakly auriculate, the branch leaves are distinctly ranked, and deciduous
branch leaves are frequently encountered. The non-stipitate plants of this
southern form are nomenclaturally centered on P. veriscolor C. Müll.
(Brazil), while the stipitate plants are nomenclaturally centered on P.
rigens Broth. (Brazil). Throughout the range of the species there are even
more slender expressions, that lack stipitate secondary stems, have more or
less unranked, non-auriculate stem leaves, and non-deciduous branch leaves. The
oldest name at the species level for this expression is Pilotrichella
pulchella Schimper ex Bescherelle (Mexico), but it had earlier been
described as Neckera pachygaster var. gracilis C. Müll.
(Venezuela).
Orthostichella
versicolor is primarily an epiphytic
species. It is occasionally collected on boulders, but field observations on
plants in this habitat point to epiphytic plants that have been able to
maintain growth after falling from the canopy. Collections taken from twigs
tend to have plants small with weakly ranked leave, while those from tree
trunks often consist of big plants with strongly ranked leaves. In some large
collections that have been carefully collected and so show all parts of the
population every one of the morphological expressions of the species can be
found.
This
species has generally been known as Pilotrichella pentasticha (Brid.)
Wijk & Marg. or Orthostichella pentasticha (Brid.) Buck every since
Robinson (1967) chose to use the oldest specific epithet (Hypnum
pentastichum Brid.) for this species complex. Hypnum pentastichum
was based on a specimen of unknown origin sent to Bridel by Philibert Commerson
(1726–1773). This specimen is not in B, however, Marshall Crosby recently
located the specimen at PC. The PC collection was made by Commerson on
Mauritius in 1769 or 1770, and unfortunately it is not an Orthostichella but
rather belongs to the genus Orthostichidium (Orthostichium
pentastichum (Brid.) Allen, comb. nov. Basionym: Hypnum
pentastichum Brid., Muscol. Recent. 2(2): 100. 1801). The second oldest
name for this species complex is Hypnum imbricatum (P. Beauv.) which is
based on a M. Bory-St. Vincent collection from Réunion. I have not been able to
locate the type of this name. It is possible that it too is an Orthostichidium
or even the same as O. pentastichum.
Orthostichella
versicolor can be confused in the
field with Squamidium nigricans (Hook.) Broth., Orthostichopsis
tenuis (Jaeger) Broth., Orthostichopsis praetermissa Buck) or
Orthostichidium quadrangulare (Schwaegr.) Allen because they all are
pendent mosses with distinctly ranked leaves. Squamidium nigricans,
Orthostichopsis tenuis, and O. praetermissa differ from Orthostichella
versicolor in having a long single costae. Squamidium nigricans also
has well-developed alar cells and often a black coloration, while the two Orthostichopsis
species have filamentous pseudoparaphyllia, densely hairy calyptrae and
vaginula, and cylindrical capsules. Orthostichopsis praetermissa further
differs from Orthostichella versicolor in having hair-pointed leaves. Orthostichella
versicolor can often be distinguished from these three species with a hand
lens beause of its branches that frequently end in filiform attenuations and
have deciduous leaves.
Orthostichidium
quadrangulare is especially close to Orthostichella
versicolor since both species have ecostate leaves with broadly incurved
upper leaf margins and weakly developed alar cells. Plants of O.
quadrangulare tend to be a little bit larger than those of Orthostichella
versicolor, and its leaves are more stiffly erect as well as more strongly
ranked. However, the only certain way to separate Orthostichidium
quadrangulare from Orthostichella versicolor is to examine their
branch primordia. In Orthostichidium quadrangulare the primordia often
give rise to clusters of uniseriate gemmae and they are surrounded by
filamentous pseudoparaphyllia. Orthostichella versicolor never has
asexual gemmae, and the branch primordia are surrounded by scale leaves.
7.
Orthostichella welwitschii (Duby) Allen & Magill, comb. nov.
Neckera welwitschii Duby, Mém. Soc. Phys. Genčve 21: 428. 1872. Meteorium welwitschii (Duby)
Jaeg., Ber. Thätigk. St. Gallischen Naturwiss. Ges. 1875–76: 243. 1877. Pilotrichella
welwitschii (Duby) Gepp in Hiern., Cat. Afr. Pl. 2(2): 292. 1901.
Protologue: Angola. Frequenter in ramulis extremis variarum arborum pendulam in
sylvis primitivis 2300–2400 ped. alt. montis Campolungo prov. Golungo-alto
regni Angolensis reperiit cel. Welwitsch. Type. Welwitsch Iter
Angolense, Dist. Golungo; 2400 feet, (S, isotype); Musci Angolensis N° 145.
Distr. Gol°-alto, ad 2800 ped., ad truncos vituntos Coffee arab. indigense, in
monte Campolungo freq. sine fr. Febr. 1855, leg. Welw. (H, isotype);
Angola: distr. Golungo-Alto. Leg. Welwitsch, 1855. n° 145 (PC, isotype).
Pilotrichella sordidoviridis C. Müll. ex Dusén,
Kongl. Svenska Vetensk. Acad. Handl. 28(2): 27. 1895., Protologue: Habitat in
Camerunia in ramulis arborum, ubi ad Bibundi emporium m. Julio a. 1892 c. fr.
legi [Dusén]. Types: Africa occ., Camerunia ad Bibundi emporium in
ramulis, Julio 1892, P. Dusén [P. Duséns mosssherbarium] (S, lectotype);
Musci Africani in Camerunia. Ad Bibundi emporium in ramulis, 21/7 1892, P.
Dusén (FH, S, isotypes); Musci Africani in Camerunia. Ad Bubundi pagum in
truncis arborum m. Julio a. 1892, P. Dusén 829 (S, isotype).
Pilotrichella
dewrevrei
Ren. & Card., Bull. Soc. Roy. Bot. Beligique 38(2): 72. 1899. Pilotrichella
sordido-veridis Dusén var. dewrevrei (Ren. & Card.) Ren. &
Card., Bull. Soc. Roy. Bot. Beligique 38(1): 25. 1900. Protologue: Congo sine
loco (Alfr. Dewčvre). Type: Congo belge. Leg. DeWčvre [Herb. J.
Cardot] (PC, holotype).
Pilotrichella
pilifolia
Dixon, Smithsonian Misc. Collect. 69(8): 8. 1918.
Prtologue. Uganda. Epiphytic; pendent in forest, Mabira, near Mubango, alt.
4,000 ft, July, 1916, Dümmer 2961; c. fr. Type: On tree trunks, pendent.
Mabira Forest, Mubango, 4000 ft, Mardh 20, Drummer 2961 (FH, isotype).
Plants
slender to medium-sized, dull, light green, yellow-green, reddish yellow or
brownish green, in loose mats often with pendent strands. Stolons creeping,
leaves well spaced. Primary stems foliose, horizontal or pendent, frequently
and irregularly branched, branches often with flagelliform tips, 2–15 cm long,
yellowish green, becoming dark red with age, in cross section with sclerodermis
of 2–5 small, thick-walled, reddish orange cells, cortex cells enlarged,
firm-walled, pale yellow, red with age, central strand absent; paraphyllia
absent, pseudoparaphyllia absent, scale leaves present; axillary hairs 3 cells
long, basal 1 cell quadrate to subquadrate, upper 2 cells broadly cylindrical,
all cells reddish; rhizoids on stolons and on primary stems, red, not or
irregularly branched, from clusters of initials abaxial to the leaf insertions.
Primary stem and branch apices turgid; stem leaves not seriately ranked, ovate,
erect, variously clasping below, 1.4–2.0 mm long, rounded to the base, abruptly
narrowly acuminate, to hair-pointed; margins broadly incurved and subentire to
serrulate above, plane or broadly incurved, serrulate or entire below; costae
absent or very shortly single or double or short single; leaf cells long
flexuose, smooth, upper cells 30–44 x 4–5, median cells 40–60 x 3–4 μm,
outer basal cells shorter and broader, inner basal cells near insertion
elongate, often porose, alar cells, subquadrate to short-rectangular extending
up the margins in 2–4 rows and across the insertion in 4–6 rows, yellow across
the insertion. Branch leaves often distinctly seriately ranked, dry or wet,
leaves obovate to oblong-obovate, often panduriform, erect to erect-spreading,
variously clasping below, 0.9–1.5 mm long, rounded to the insertion, apex
apiculate to short cuspidate, cuspid 40-80 µm long; margins broadly incurved
and subentire to serrulate above, plane or broadly incurved, serrulate or
entire below; costae very short, double, single, occasionally absent; leaf
cells long flexuose, smooth, upper cells 20–40 x 4–5, median cells 30–60 x 3–4
μm, outer basal cells shorter and broader, inner basal cells near
insertion elongate, often porose, alar cells red to red-yellow, subquadrate to
short-rectangular extending up the margins in 2–4 rows and across the insertion
in 4–6 rows, yellow across the insertion. Dioicous. Perigonia not seen.
Perichaetia lateral, unfertilized perichaetia 1.0–1.5 mm long, paraphyses and archegonia
numerous; fertilized perichaetial 2.0–2.5 mm long, leaves lanceolate 2–3 mm
long, vaginula hairy. Setae wavy, 3–5 mm long, smooth below,
roughened-papillose above, yellow, becoming yellowish red with age. Capsules
exserted, erect to somewhat inclined, ovoid to short-cylindrical, 1.0–1.5 mm
long, neck moderately developed; exothecial cells small, rounded in 2–3
rows below the mouth, subquadrate, irregularly short-rectangular, or oblate,
firm-walled below, cells in neck smaller than the median cells; stomata
superficial on neck; opercula obliquely long-rostrate, 1.0 mm long;
annuli rudimentary, consisting of 2–3 rows of thin-walled enlarged cells; peristome
yellowish white, exostome teeth linear, 0.3–0.4 mm long, dorsal (outer) surface
lightly horizontally striate at base, smooth or finely papillose above,
trabeculae weakly developed on both sides, endostome nearly as long as
exostome, basal membrane very low, segments filamentous, narrowly perforated,
not keeled, smooth to papillose, cilia rudimentary or absent. Spores round,
lightly roughened, 18–24 μm long. Calyptra not seen.
Etymology.
The specific epithet welwitschii is given in honor of its collector,
Friedrich Martin Josef Welwitsch (25 Feb 1806–20 Oct 1872), an Austrian
botanist. Welwitsch collected plants in Angola (1853–1861) for the Portuguese
government, and during that time also collected in other parts of Southwest
Africa (Stafleu & Cowan 1988, Frahm & Eggers 2001, includes portrait).
Illustrations:
Dusén (1895, .p.
27 a--c, Pl. 2, fig. 19, Pl. 4 fig. 3)
Habitat:
On tree trunks and branches; 852–1219 m.
Distribution.
West, West Central, East, and South Tropical Africa.
Selected
specimens examined.
ANGOLA.
Dist. Golungo: Welwitsch (S).
CAMEROON.
Bubundi,
Dusén 829 (S).
IVORY COAST. In silva Akrčdiou,
inter Thiassale et Dabou, A. Jolly (B).
TANZANIA. Stromgebiet des oberen
Ruhudje, Landschaft [District] Lupembe, nordlich des Flusses, Schlieben 726
(B).
ZAIRE. Dewčvre (PC).
Orthostichella
welwitschii is a small to
medium-sized species with extremely long hair-pointed stem leaves. The species
appears to lack stipitate primary stems, but this may be an artifact due to the
few collections from which the species is known. The stem leaves of P.
welwitschii are more or less similar in form to those of O.
pandurifolia, however, they are not as consistently costate and its branch
leaves are identical to those of O. versicolor. In terms of overall
morphology O. welwitschii appears most similar to O. versicolor.
Unfortunately the pendent stems of O. versicolor often have hair-pointed
leaves and this is likely cause considerable difficulty in separating the two
species. The stem leaves of O. welwitschii, however, consistently
have extremely long hair-points and often the hair-points are kinked. This
feature is especially pronounced in the stem buds. In most collections of O.
versicolor short-pointed stem leaves can be found. There are
included in this treatment of O. welwitwschii a number of small slender
collections that consistent entirely of pendent stems and branches. These
collections have hair-pointed stem leaves, but they may actually represent
pendent forms of O. versicolor.
Excluded taxa.
Neckera trichophoroides Hampe, Linnaea 38: 668. 1874. Pilotrichella
trichophoroides (Hampe) Jaeg., Ber. Thätigk. St. Gallischen Naturwiss. Ges.
1875–76: 259. 1877. Meteorium trichophoroides (Hampe) Mitt., Trans. Roy.
Soc. Victoria 19: 82. 1882. Barbella trichophoroides (Hampe) Broth.,
Nat. Pflanzenfam. 1(3): 284. 1906. Protologue: Lord Howe’s Island, Summit of
Mount Gower. Type: Lord How’e Island, Summmit Mount Gower (S, isotype).
Fleischer
(1908) made this a synonym of Barbella enervis (Mitt.) Fleisch. This
opinion was confirmed by Streimann (1991) who further considered B. enervis a
synonym of Barbella cubensis (Mitt.) Broth. Barbella cubensis is
now a synonym of Barbellopsis trichophora (Mont.) Buck (Streimann 1993,
Buck 1998). Wijk et al. (1964) attribute the name Neckera
trichophoroides to Hampe ex C. Müll., however there is no indication
in the protologue that Carl Müller had anything to do with the species.
Pilotrichella biformis Hampe var. congesta Ren. & Card., Bull.
Soc. Roy. Bot. Beligique 41(1): 76. 1905. Squamidium biforme (Hampe)
Broth. var. congesta (Ren. & Card.) Een, Lindbergia 3: 217. 1976.
Protologue: La Réunion: St Philippe (rev. Rodriguez);
Madagascar: Ampériféry (rev. Campenon). Types: Rodriguez (PC
lectotype); Campenon (H, PC syntypes).
Allen
and Crosby (1986) synonmized this taxon with Squamidium brasiliense (Hornsch.)
Broth.
Pilotrichella capilliramea C. Müll. in Par., Index Bryol. Suppl. 270. 1900, invalid
name, lacks a description.
No
material of this taxon was examined. It is restricted to the Philippines and is
not likely to be an Orthostichella. The last checklist of Philippine
mosses (Tan & Iwatsuki 1991) makes no mention of this taxon.
Pilotrichella chlorothrix C. Müll., Flora 73: 490. 1890. Squamidium
chlorothrix (C. Müll.) Broth., Nat. Pflanzenfam. 1(3): 809. 1906.
Protologue: Africa or. trop. in sylva primaeva montis Kilima-Ndscharo,
1500–3000 m: Dr. Hans Meyer 1889. Type: Africa or. trop. in sylva
primaeva montis Kilima-Ndscharo, 1500–3000 m: Dr. Hans Meyer 1889 (H).
Allen
and Crosby (1986) synonmized this taxon with Squamidium brasiliense (Hornsch.)
Broth.
Orthostichopsis chrysoneura (Hampe ex C. Müll.) Broth. Nat. Pflanzenfam.
1(3): 805. 1906. Neckera chrysoneura Hampe ex C. Müll., Linnaea
40: 263. 1876. Pilotrichella chrysoneura (Hampe ex C. Müll.)
Jaeg., Ber. Thätigk. St. Gallischen Naturwiss. Ges. 1875–76: 256. 1877. Protologue:
Comoro Islands. Patria. Comoro-insula Johanna, unde ad filices arborescentes
involvendas misit J. M. Hildebrandt 1875. Types. Africa orient:
Comor-insula Johanna, ad arbores, leg. Hildebrandt 1875 (S, isotype);
Archipelagus Comorensis, insula Johanna: J. M. Hilderandt, 1875 (S,
isotype).
Renauld
and Cardot (1915) considered Orthostichopsis chrysoneura a local race of
the Madagascan O. subimbricata (Hampe) Broth). Argent (1973b)
also placed O. chrysoneura into the synonymy of Orthostichopsis
subimbricata, although he did not see the type of O. subimbricata. Orthostichopsis
chrysoneura is gametophytically extremely close to the southern South
American O. tenuis (Jaeg.) Broth. The two differ only in seta length: O.
tenuis capsules exserted; O. subimbricata capsules immersed.
Pilotrichella densiramea Broth., Bot. Jahrb. Syst. 20: 197. 1894. Squamidium
densirameum (Broth.) Broth., Nat. Pflanzenfam. 1(3): 809. 1906. Squamidiuim
biforme (Hampe) Broth., var. densirameum (Broth.) Bizot, Acta Bot.
Acad. Sci. Hung. 28: 33. 1982. Protologue: Tanzania. Usambara: Handeď,
trockener Hochwald, an Baumrinde (Holst n. 9056) und Lugulua Wald, auf
alten Bäumen (n. 2629a). Types: Usambara: Handeď, trockener Hochwald, an
Baumrinde Holst n. 9056 (S, syntype); Lugulua Wald, auf alten Bäumen Holst
2629a, H, lectotype).
Allen
and Crosby (1986) synonomized this taxon with Squamidium brasiliense
(Hornsch.) Broth.
Pilotrichella
dubia C. Müll. in Paris, Index Bryol. 944. 1897, invalid name
lacks a description. Protologue: Pac.: Nov. Guinea. (M. Yule).
This
taxon is excluded from Orthostichella based on its occurrence in New
Guinea.
Pilotrichella eustichia Schimp. in Kindb., Enum. Bryin. Exot. Suppl.
2: 102. 1891, invalid name, lacks a description. No specimen was cited along
with this name.
There
is a collection in H from the Bescherelle herbarium named Pilotrichella
eustichia Sch. The specimen, collected in Mauritius (Danty n. 14),
is Orthostichidium pentastichum.
Pilotrichella kraussei Lor., Moosstudien 164. 1864. Protologue: Chile. Prope
Valdiviam leg.Dr. Krausse. Isotype: Chile. Ad arborum ramos in
silvis densis udis. 0–2000’ prope Correl, puerto de Valdivia, leg. Dr. H.
Krausse (NY).
This
taxon is synonymous with Weymouthia cochlearifolia (Schwägr.) Dix.
Pilotrichella lepto-erioclada C. Müll. in Par., Index Bryol. 946. 1896, invalid name,
lacks a description. Based on: Musci Africani in Camerunia a P. Dusén collecti.
Ad N’dian emporium in ramulis arborum die 26 m. Martii a. 1892, Dusén 800
(S).
The
plants in this collection belong to the Sematophyllaceae.
Pilotrichella percapillaris C. Müll. in Par., Index Bryol. 948. 1897,
invalid name, lacks a description. Based on V. Afr., Kamerun, Jumbo 1080 m,
17/4/92, P. Dusén (S).
The
plants in this collection are Rhacopilopsis trinitensis (C. Müll.)
Britt. ex Dix.
Hildebrantiella
phleoides (Desvaux
ex Brid) Allen & Magill, comb. nov. Hypnum phleoides Desvaux ex
Brid., Bryol. Univ. 2: 431. 1827. Pilotrichella phleoides (Desvaux ex
Bridel) Jaeger, Ber. Thäigk. St. Gallischen Naturwiss. Ges. 1875–76: 261. 1877.
Protologue: In insula Bourbonis [Reunion] in arborum cortice habitat. Type: Neckera
phleoides mihi (Hypnum Desv.) Ins. Bourbon, hb. Brid. (JE, isotype).
Hildebrantiella pachyclada Besch.,
Ann. Sci. Nat., Bot. sér. 6, 10: 264. 1880, syn. nov.
Pilotrichella
recurvula C.
Müll., Hedwigia 41: 129. 1902. Protologue: Australia. Habitatio. Queensland,
Kev Range, Ballendea, 1889; F. M. Bailey. Hb. Brotheri 1890.
Streimann
(1991) examined the type of this species and determined it was identical to Calyptothecium
recurvulum (Broth.) Broth..
Pilotrichella rehmanni C. Müll. in Geh., Rev. Bryol. 5: 70. 1878,
invalid name, lacks a description.
This
taxon is synonymous with Squamidium brasiliense (Hornsch.) Broth.
Pilotrichella serricola C.Müll. in Par., Index Bryol. 949 1897,
invalid name, lacks a description. Based on E. Ule, Bryotheca brasiliensis n.
67 (BM).
This
taxon is synonymous with Squamidium brasiliense (Hornsch.) Broth.
Pilotrichella subbiformis Ren. & Card., Bull. Soc. Roy. Bot. Beligique
39(2): 110. 1900. Erythrodontium subbiformis (Ren. & Card.) Broth.,
Nat. Pflanzenfam. 1(3): 888. 1907. Protologue: Congo. Rég. V: Kisantu (J.
Gillet, 1900, no 712). Type. Congo belge: Kisantu, leg. Gillet,
Herb. F. Renauld (S, isotype).
The
very small isotype of this taxon at S belongs to Erythrodontium.
Pilotrichella subsemitorta Broth. in Naveau, Rev. Bryol., n.s. 1: 38.
1928, invalid name, lacks a description.
According
to Theriót (1929) this is an error for Papillaria subsemitorta Broth. ex
Ther.
Nomina nuda
not seen.
Pilotrichella
ankasinae
C. Müller in Kindberg, Enum. Bryin. Exot., Suppl. 2 102 1891.
Protologue: nothing. Invalid name, lacks a description.
Pilotrichella
chrysoneura var. viridis Renauld & Cardot, Rev. Bot. Bull. Mens. 9: 221
1891. Protologue: [Madagascar]. Invalid name, lacks a description
Pilotrichella
cuspidata Broth.,
var. gracilescens Tosco & Piovano, Allionia 4: 124. 1956.
Protologue: Kenya (Central Prov.) – Massailand, foresta (Leg. G. Balbo,
6 genn. 1908); Tuthu, Monte Gasongori (Leg. G. Balbo, 7 lug. 1908).
Invalid name, holotype not indicate.
Pilotrichella
platyclada Par.,
Index Bryol. 948. 1897, invalid name lacks a description. Orthostichella
platyclada C. Müll. in Par., Index Bryol. 948. 1897, invalid name
lacks a description. Based on: Musci
Africani in Camerunia a P. Dusén collecti. 846.
Pilotrichella
porotrichoides C. Müller in Paris, Index Bryol. Suppl. 271 1900,
invalid name, lacks a description. Based on: Sikora Pl. Madag. n. 14234
Pilotrichella
stracheyana C.
Müll. in Kern, Rev. Bryol. 19: 41. 1892, invalid name lacks a
description. Protologue: As stuffing in a Satyr pheasant (Ceriornis satyra).
Types not seen.
Pilotrichella
debilinervis Ren. & Card., Bull. Soc. Roy. Bot. Beligique 34(2):
70. Protologue: Hab. Bourbon: Salazie, in sylva “de Belouze” dicta (Chauvet;
hb. de Poli).
Pilotrichella
delicatula
Brotherus in Mildbraed, Wiss. Erg. Deut. Zentr.-Afr. Exped., Bot. 2: 160 1910.
Protologue: Vulkan-Gebiet [Central Africa]: Bambusmischwald von Bugoie, ca.
2100 m ü. M[ildbraed]. (steril im Sept. 1907–n. 2034).
Illegitimate homonym, not Pilotrichella delicatula (Besch.) Schimp.,
(1897).
Hypnum
imbricatum Palisot
de Beauvois, Prodr. Aethéogam. 65. 1805. Pilotrichella imbricata
(Palisot de Beauvois) Bescherelle, Mem. Soc. Sci. Nat. Cherbourg 16: 222 1872.
Protologue: l'Ile de Bourbon [Reunion] Bory-St.-Vincent
Pilotrichella
imbricatula
C. Müller ex Geheeh, Abh. Naturwiss. Vereine Bremen 7: 209 1881.
Protologue: [Madagascar] Wald von Ambatondrazaka, [Brogen or Borchgrewink]
6 Decbr. 1877. [= P. isleana Besch fid. Card. in Grand., Hist
Madag. 39: 350, 1915.]
Pilotrichella
imbricatula var. exserta Thériot, Recueil Publ. Soc. Havraise Etudes Diverses
1929: 116 1930. Protologue: [Madagascar] Massif de l'Andringitra. (leg. H.
Humbert).
Pilotrichella
imbricatula var. nervosa Renauld & Cardot, Bull. Soc. Roy. Bot. Belgique
33(2): 126 1895. Pilotrichella isleana var. nervosa (Renauld &
Cardot) Cardot in Grandidier Hist. Phys. Madagascar, Mousses 39: 351 1915.
Protologue: Madagascar: inter Savondronina et Ranomafana (Dr Besson).
— (Renauld, Musci Mascareno-Madagascarienses Exsiccati, n°.228).
Pilotrichella
latiramea
C.Müller ex Dusén, Kongl. Svenska Vetensk. Acad. Handl. 28(2): 29. 1895.
Protologue: Habitat in Camerunia in ramulis arborum, ubi ad Ekundu Ndene
emporium m. Julio a. 1892 legi [Dusén]
Neckera
pseudo-imbricata C. Müll., Bot. Zeitung (Berlin) 20: 381. 1862. Pilotrichella
pseudoimbricata (C. Müller) Jaeger, Ber. Thätigk. St. Gallischen Naturwiss.
Ges. 1875–76: 255 1877. Protologue: Comoro Islands. Insula Anjoana ad canal. de
Mozambique Dr. Peters 1843.
Neckera pseudo-imbricata
C. Müll. var. pallescens C. Müll., Linnaea 40: 265 1876. Pilotrichella
pseudoimbricata var. pallescens (C. Müller) Paris, Index Bryol. 948 1897.
No specimen cited in protologue.
Neckera pseudo-imbricata
C. Müll. var. rufescens C. Müll., Linnaea 40: 265 1876. Pilotrichella
pseudoimbricata var. rufescens (C. Müller) Paris, Index Bryol. 948
1897. No specimen cited in protologue.
Pilotrichella
ragazzii
Brizi, Annuario Reale Ist. Bot. Roma 5: 79. 1893. Prtologue: [Ethiopia].
Comunissima sul tronco degli alberi nella foresta di Fekerič-Ghemb. 20 Maggio
1885. — Let-Marefiŕ. 21 Marzo 1885. [V. Ragazzi].
Pilotrichella
sericea C.
Müll. ex Kindb., Enum. Bryin. Exot., Suppl. 2: 102. 1891, new new for Orthostichella
sericea C. Müll., Flora 73: 492. 1890, invalid name due to an invalid
generic name. Protologue: Patria. Africa or. trop., Leikipia in regione
occidentalis montis Kilima-Ndscharo, ad pedem der Aberdare-Kette: L. Höhnel in
Exped. Telekiana 1887.
Literature cited.
Bartram, E. B. 1933.
Manual of Hawaiian mosses. Bernice P. Bishop Mus. Bull. 101: 1–275.
Bescherelle, E.
1872. Prodromus bryologiae mexicanae ou énumération des mousses du Mexique avec
description des espčces nouvelles. Mém. Soc. Sci. Nat. Cherbourg 16: 143–256.
Brotherus, V. F.
1906. Pilotrichella. In: A. Engler & K. Prantl (eds.), Nat.
Pflanzenfam. 1 (3): 809–811.
_____.
1925. Musci. In: A. Engler (ed.), Nat. Pflanzenfam. (ed. 2) 11: I–iv,
1–542.
Buck, W. R. 1994. A
new attempt at understanding the Meteoriaceae. J. Hattori Bot. Lab. 75: 51–72.
_____a. The resurrection of Orthostichella.
Bryologist 97: 434–435.
_____. 1998. Pleurocarpous mosses of the West Indies.
Mem. New York Bot. Gard. 82: 1–400.
_____ & B. Goffinet. 2000. Morphology and
classification of mosses, pp. 71–123.
In: A. J. Shaw & B. Goffinet (eds.), Bryophyte Biology. Cambridge
University Press, Cambridge.
Churchill, S. P.
& E. Linares C. 1995. Prodromus Bryologiae Novo-Granatensis. Bibliot. José
Jerónimo Triana 12: [I–x] 1–924.
Duarte‑Bello,
P. P. 1997. Musgos de Cuba. Fontqueria 47: xxii + 717 pp.
Fleischer, M. 1908. Die Musci der Flora von
Buitenzorg. Volume 3: i–xxiv, 645–1103 pp. E. J. Brill, Leiden.
Florschütz, P. A.
1964. Musci: In: J. Lanjouw (ed.), Flora of Suriname 6(1): 1–271. E. J. Brill,
Leiden.
Frahm, J.-P. & J. Eggers. 2001. Lexikon
Deutschsprachiger Bryologen. 672 pp. Books on Demand, Norderstedt, Germany.
Goffinet, B. & W. R. Buck. 2004. Systematics of
Bryophyta (mosses): from molecules to a revised classification. Monogr. Syst.
Bot. Missouri Bot. Gard. 98: 205–239.
Gradstein, S. R., S.
P. Churchill & N. Salazar-Allen. 2001. Guide to the Bryophytes of Tropical
America. Mem. New York Bot. Gard. 86: viii, 1–577.
Greuter, W. et
al. 2000. International Code of Botanical Nomenclature. Regnum Veg. 138:
I–xviii + 474 pp.
Jaeger, A. & F.
Sauerbeck. 1877. Genera et species muscorum systematice disposita seu
adumbratio florae muscorum totius orbis terrarum. Ber. Thätigk. St. Gallischen
Naturwiss. Ges. 1875-76: 201–371.
Lorentz,
P. G. 1864. Moosstudien. Leipzig.
Magill, R. E. &
J. van Rooy. 1998. Flora of Southern Africa. Bryophyta. Part 1. Musci. Fascicle
3. Erpodiaceae–Hookeriaceae. I–vii + 445–622 pp. Government Printer, Pretoria.
Müller, C. 1850. Neckera.
In: Synopsis Muscorum Frondosorum 2: 40–146. Berlin.
_____. 1879. Musci
Fendleriani Venezuelenses. Linnaea 42: 461–502.
Renauld, F. & J.
Cardot. 1893. Musci exotici novi vel minus cogniti. Bull. Soc. Roy. Bot. Beligique 32(2): 8–40.
Stafleu, F. A. & R. S. Cowan. 1983. Taxonomic
Literature. Volume IV: P–Sak. Regnum Vegetabile 110: 1–1214.
_____. 1988. Taxonomic Literature. Volume VII: W–Z.
Regnum Vegetabile 116: LVI + 1–653.
Schwägrichen, F. 1816. Species Muscorum Frondosorum
Supplementum 1(2): 1–373, Tab. 50–100. J. A. Barth, Lipsiae.
Striemann, H. 1991. Taxonomic studies on Australian
Meteoriaceae (Musci). 2: The genera Aerobryopsis, Barbella, Floribundaria,
Meteoriopsis, Meteorium and Weymouthia. J. Hattori Bot. Lab. 69:
277–312.
_____. 1993. Barbella trichophora, and older
name for B. cubensis (Musci: Meteoriaceae). Bryologist 96: 223–225.
Tan, B. C. & Z. Iwarsuki. 1991. A new annotated
Philippine moss checklist. Harvard Pap. Bot. 3: 1–64.
Vitt, D. H. 1984.
Classification of the Bryopsida. In: R. M. Schuster (ed.), New Manual of
Bryology 2(13): 696–759. Hattori
Botanical Laboratory, Nichinan.
Walther, K. 1983.
Syllabus der Pflanzenfamilien. Vol. 2. Bryophytina, Laubmoose. 108 pp. G.
Borntraeger. Berlin.
Wijk, van der R., W. D. Margadant & P. A.
Florschütz. 1964. Index Muscorum. Vol. III (Hypnum–O). Regnum Veg. 33:
1–529.
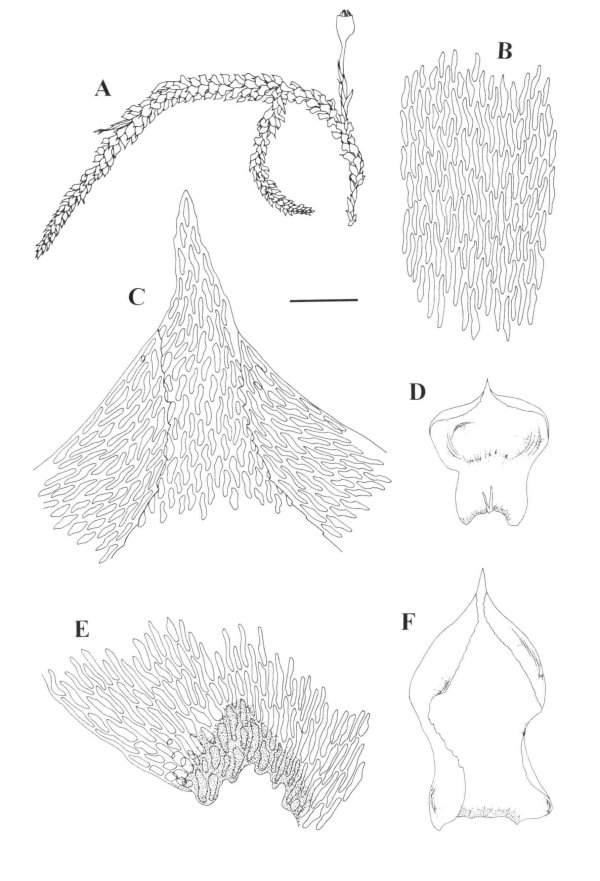 Orthostichella hexasticha
Orthostichella hexasticha Orthostichella hexasticha
Orthostichella hexasticha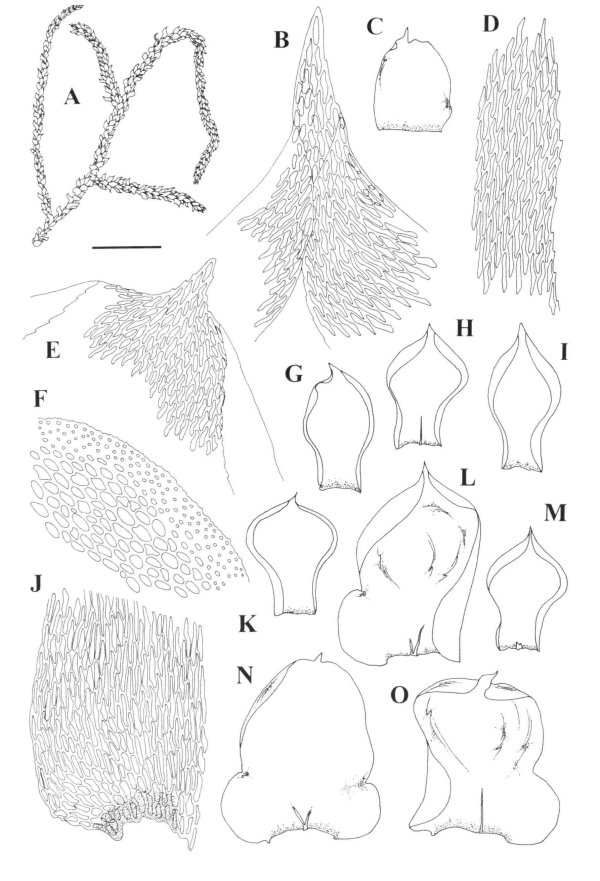 Orthostechella roseana
Orthostechella roseana 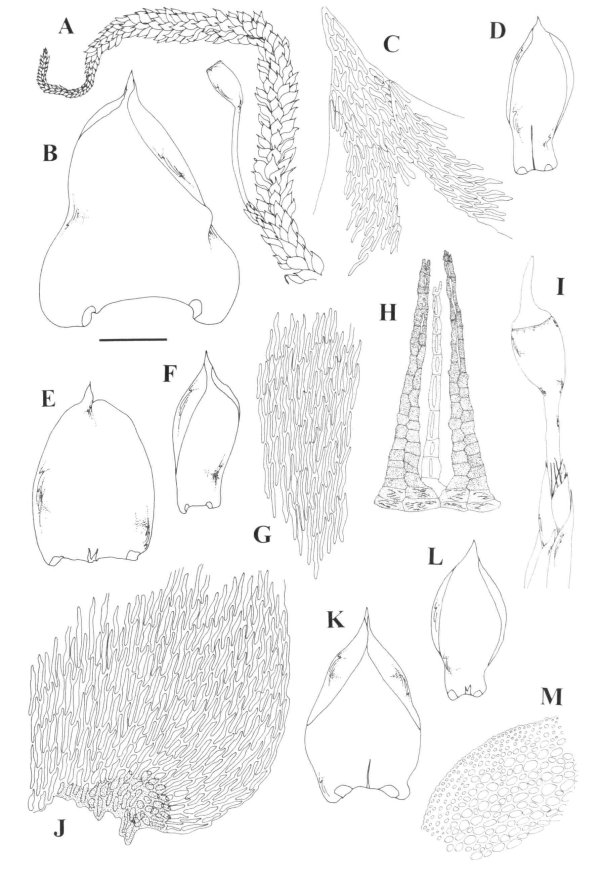 Orthostichella versicolor
Orthostichella versicolor