QUICK SEARCH
MO PROJECTS:
Africa
Asia/Pacific
Mesoamerica
North America
South America
General Taxonomy
Photo Essays
Training in Latin
America
MO RESEARCH:
Wm. L. Brown Center
Bryology
GIS
Graduate Studies
Research Experiences
for Undergraduates
Imaging Lab
Library
MBG Press
Publications
Climate Change
Catalog Fossil Plants
MO DATABASES:
W³MOST
Image Index
Rare Books
Angiosperm
Phylogeny
Res Botanica
All Databases
INFORMATION:
What's New?
People at MO
Visitor's Guide
Herbarium
Jobs & Fellowships
Symposium
Research Links
Site Map
Search
THE CLASSIFICATION OF NATURAL AND |
|
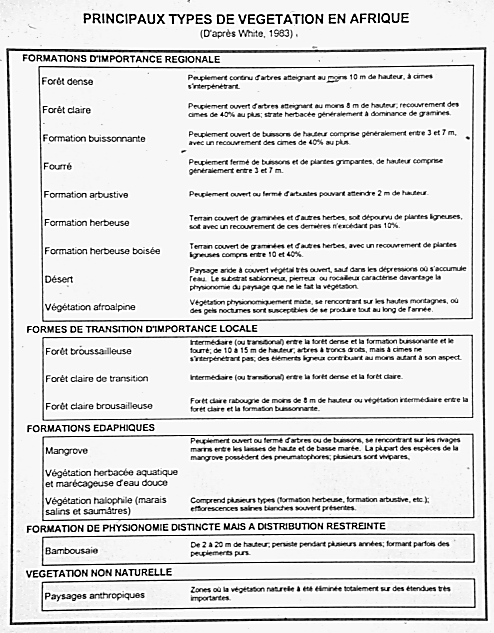
A useful example of a more structured and objective approach is that of Frank White and his collaborators. White (1983, 1993) published a vegetation classification and map for continental Africa, based on the concept that physiognomic data, and in particular the cover and height of the vegetation, are fundamentally different from chorological data on patterns of species distribution. His approach was to study these two aspects independently, starting with physiognomy.
White developed an intentionally simple system in which 17 major vegetation types are recognized in 5 categories. For example, forests are defined as all vegetation comprising a continuous stand of trees at least 10m tall with interlocking crowns.
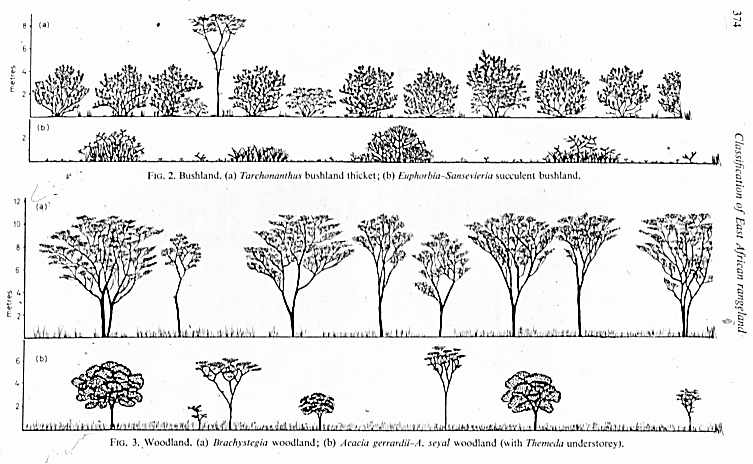
Bushland (formation buissonnante) includes open stands of bushes 3-7 m tall, with a canopy cover of 40% or more, and woodland (forÊt claire) is defined as open stands of trees at least 8 m tall, with a canopy cover of 40% or more, and with a ground layer usually dominated by grasses and other herbs, as can be seen in illustrative forest profiles prepared by Pratt et al. (1966).
|
 |
It is important to note that grasslands (formations herbeuses)
were defined by White (1983) specifically as land covered with grasses and other herbs, with woody plants covering no more than 10% of the ground. Several authors have argued strongly against the use of imported terms such as prairie, savanna, steppe and pseudosteppe in Africa (see Chapman and White, 1970; Descoings, 1973, 1978; White, 1983). For example, savanna has been applied in so many ways that it is highly ambiguous (see also Eiten, 1986, 1992); steppe should be restricted to the characteristic grasslands of eastern Europe (cf. Eiten, 1992); and prairie
refers to a type of vegetation that was once widespread in
central North America. White's (1983) classification accommodates these and other forms of vegetation without having to use such problematical terms (but see Lawesson, 1994, for some critical comments), and we feel that it would be appropriate to abandon them definitively when discussing the vegetation of Madagascar.
 12 species of Diospyros Ebenaceae) fruiting simultaneously at Nosy Mangabe Special Reserve |
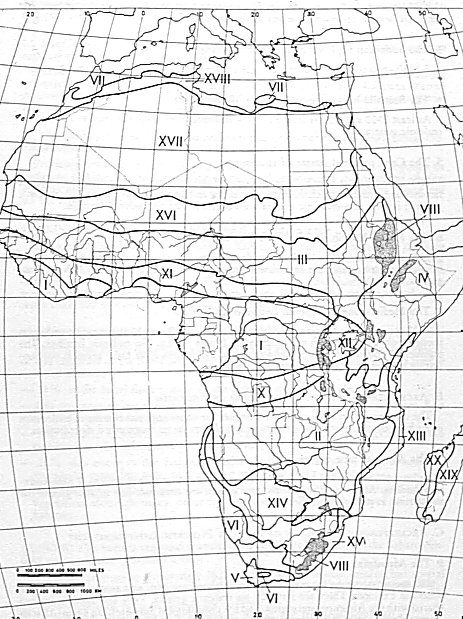
The geographic distribution of plant species is not entirely random. Clear patterns exist, and they can be detected by careful analysis of the distribution patterns of a large numbers of species. White (1976, 1979, 1983, 1993) used this kind of chorological approach in Africa. For example, the Guineo-Congolean regional center of endemism (Zone I on White's map) comprises about 8,000 spp, of which about 80% are endemic to the region; there are also 9 endemic plant families and 1/4 of the genera are endemic (White, 1983, 1993). It is important to note that all of these regions, or phytochoria, are based exclusively on chorological data.
|
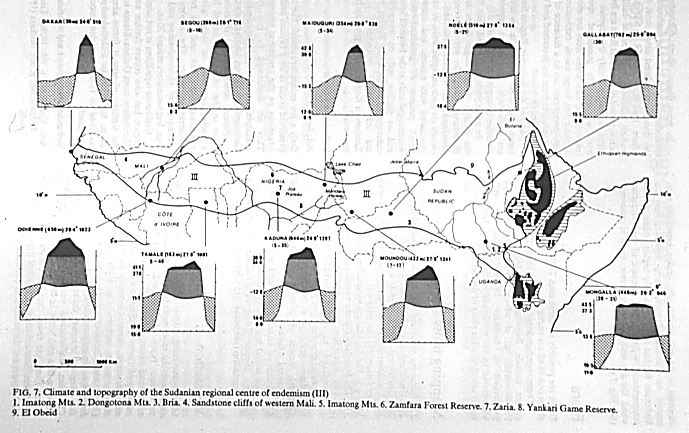
Once defined, however, these regions almost always comprise areas with very similar bioclimatological characteristics, as illustrated by clima-diagrams from various sites within the Sudanian regional center of endemism, which spans from Senegal to Ethiopia (White, 1983). White mapped his basic physiognomic categories (forest, bushland, thicket, etc.) onto the chorological zones, which serve to subdivide the vegetation types into cohesive regions on the basis of chorology.
|
© 1995-2025 Missouri Botanical Garden, All Rights Reserved
4344 Shaw Blvd.
St. Louis, MO 63110
(314) 577-5100
Technical Support