EMBRYOPSIDA Pirani & Prado
FISHGametophyte dominant, independent, multicellular, initially ±globular, not motile, branched; showing gravitropism; glycolate oxidase +, glycolate metabolism in leaf peroxisomes [glyoxysomes], acquisition of phenylalanine lysase* [PAL], flavonoid synthesis*, microbial terpene synthase-like genes +, triterpenoids produced by CYP716 enzymes, CYP73 and phenylpropanoid metabolism [development of phenolic network], xyloglucans in primary cell wall, side chains charged; plant poikilohydrous [protoplasm dessication tolerant], ectohydrous [free water outside plant physiologically important]; thalloid, leafy, with single-celled apical meristem, tissues little differentiated, rhizoids +, unicellular; chloroplasts several per cell, pyrenoids 0; centrioles/centrosomes in vegetative cells 0, microtubules with γ-tubulin along their lengths [?here], interphase microtubules form hoop-like system; metaphase spindle anastral, predictive preprophase band + [with microtubules and F-actin; where new cell wall will form], phragmoplast + [cell wall deposition centrifugal, from around the anaphase spindle], plasmodesmata +; antheridia and archegonia +, jacketed*, surficial; blepharoplast +, centrioles develop de novo, bicentriole pair coaxial, separate at midpoint, centrioles rotate, associated with basal bodies of cilia, multilayered structure + [4 layers: L1, L4, tubules; L2, L3, short vertical lamellae] (0), spline + [tubules from L1 encircling spermatid], basal body 200-250 nm long, associated with amorphous electron-dense material, microtubules in basal end lacking symmetry, stellate array of filaments in transition zone extended, axonemal cap 0 [microtubules disorganized at apex of cilium]; male gametes [spermatozoids] with a left-handed coil, cilia 2, lateral, asymmetrical; oogamy; sporophyte +*, multicellular, growth 3-dimensional*, cuticle +*, plane of first cell division transverse [with respect to long axis of archegonium/embryo sac], sporangium and upper part of seta developing from epibasal cell [towards the archegonial neck, exoscopic], with at least transient apical cell [?level], initially surrounded by and dependent on gametophyte, placental transfer cells +, in both sporophyte and gametophyte, wall ingrowths develop early; suspensor/foot +, cells at foot tip somewhat haustorial; sporangium +, single, terminal, dehiscence longitudinal; meiosis sporic, monoplastidic, MTOC [= MicroTubule Organizing Centre] associated with plastid, sporocytes 4-lobed, cytokinesis simultaneous, preceding nuclear division, quadripolar microtubule system +; wall development both centripetal and centrifugal, 1000 spores/sporangium, sporopollenin in the spore wall* laid down in association with trilamellar layers [white-line centred lamellae; tripartite lamellae]; plastid transmission maternal; nuclear genome [1 C] <1.4 pg, main telomere sequence motif TTTAGGG, KNOX1 and KNOX2 [duplication] and LEAFY genes present, ethylene involved in cell elongation; chloroplast genome with close association between trnLUAA and trnFGAA genes [precursors for starch synthesis], tufA, minD, minE genes moved to nucleus; mitochondrial trnS(gcu) and trnN(guu) genes +.
Many of the bolded characters in the characterization above are apomorphies of more or less inclusive clades of streptophytes along the lineage leading to the embryophytes, not apomorphies of crown-group embryophytes per se.
All groups below are crown groups, nearly all are extant. Characters mentioned are those of the immediate common ancestor of the group, [] contains explanatory material, () features common in clade, exact status unclear.
POLYSPORANGIOPHYTA†
Sporophyte well developed, branched, branching dichotomous, potentially indeterminate; hydroids +; stomata on stem; sporangia several, terminal; spore walls not multilamellate [?here].
II. TRACHEOPHYTA / VASCULAR PLANTS
Sporophyte long lived, cells polyplastidic, photosynthetic red light response, stomata open in response to blue light; plant homoiohydrous [water content of protoplasm relatively stable]; control of leaf hydration passive; plant endohydrous [physiologically important free water inside plant]; PIN[auxin efflux facilitators]-mediated polar auxin transport; (condensed or nonhydrolyzable tannins/proanthocyanidins +); borate cross-linked rhamnogalactan II, xyloglucans with side chains uncharged [?level], in secondary walls of vascular and mechanical tissue; lignins +; roots +, often ≤1 mm across, root hairs and root cap +; stem apex multicellular [several apical initials, no tunica], with cytohistochemical zonation, plasmodesmata formation based on cell lineage; vascular development acropetal, tracheids +, in both protoxylem and metaxylem, G- and S-types; sieve cells + [nucleus degenerating]; endodermis +; stomata numerous, involved in gas exchange; leaves +, vascularized, spirally arranged, blades with mean venation density ca 1.8 mm/mm2 [to 5 mm/mm2], all epidermal cells with chloroplasts; sporangia in strobili, sporangia adaxial, columella 0; tapetum glandular; sporophyte-gametophyte junction lacking dead gametophytic cells, mucilage, ?position of transfer cells; MTOCs not associated with plastids, basal body 350-550 nm long, stellate array in transition region initially joining microtubule triplets; archegonia embedded/sunken [only neck protruding]; embryo suspensor +, shoot apex developing away from micropyle/archegonial neck [from hypobasal cell, endoscopic], root lateral with respect to the longitudinal axis of the embryo [plant homorhizic].
[MONILOPHYTA + LIGNOPHYTA]Sporophyte growth ± monopodial, branching spiral; roots endomycorrhizal [with Glomeromycota], lateral roots +, endogenous; G-type tracheids +, with scalariform-bordered pits; leaves with apical/marginal growth, venation development basipetal, growth determinate; sporangium dehiscence by a single longitudinal slit; cells polyplastidic, MTOCs diffuse, perinuclear, migratory; blepharoplasts +, paired, with electron-dense material, centrioles on periphery, male gametes multiciliate; nuclear genome [1 C] 7.6-10 pg [mode]; chloroplast long single copy ca 30kb inversion [from psbM to ycf2]; mitochondrion with loss of 4 genes, absence of numerous group II introns; LITTLE ZIPPER proteins.
LIGNOPHYTA†
Sporophyte woody; stem branching axillary, buds exogenous; lateral root origin from the pericycle; cork cambium + [producing cork abaxially], vascular cambium bifacial [producing phloem abaxially and xylem adaxially].
SEED PLANTS† / SPERMATOPHYTA†
Growth of plant bipolar [plumule/stem and radicle/root independent, roots positively geotropic]; plants heterosporous; megasporangium surrounded by cupule [i.e. = unitegmic ovule, cupule = integument]; pollen lands on ovule; megaspore germination endosporic, female gametophyte initially retained on the plant, free-nuclear/syncytial to start with, walls then coming to surround the individual nuclei, process proceeding centripetally.
EXTANT SEED PLANTS
Plant evergreen; nicotinic acid metabolised to trigonelline, (cyanogenesis via tyrosine pathway); microbial terpene synthase-like genes 0; primary cell walls rich in xyloglucans and/or glucomannans, 25-30% pectin [Type I walls]; lignin chains started by monolignol dimerization [resinols common], particularly with guaiacyl and p-hydroxyphenyl [G + H] units [sinapyl units uncommon, no Maüle reaction]; roots often ≥1 mm across, stele diarch to pentarch, xylem and phloem originating on alternating radii, cork cambium deep seated, gravitropism response fast; stem apical meristem complex [with quiescent centre, etc.], plasmodesma density in SAM 1.6-6.2[mean]/μm2 [interface-specific plasmodesmatal network]; eustele +, protoxylem endarch, endodermis 0; wood homoxylous, tracheids and rays alone, tracheid/tracheid pits circular, bordered; mature sieve tube/cell lacking functioning nucleus, sieve tube plastids with starch grains; phloem fibres +; cork cambium superficial; leaf nodes 1:1, a single trace leaving the vascular sympodium; leaf vascular bundles amphicribral; guard cells the only epidermal cells with chloroplasts, stomatal pore with active opening in response to leaf hydration, control by abscisic acid, metabolic regulation of water use efficiency, etc.; branching by axillary buds, exogenous; prophylls two, lateral; leaves with petiole and lamina, development basipetal, lamina simple; sporangia borne on sporophylls; spores not dormant; microsporophylls aggregated in indeterminate cones/strobili; grains monosulcate, aperture in ana- position [distal], primexine + [involved in exine pattern formation with deposition of sporopollenin from tapetum there], exine and intine homogeneous, exine alveolar/honeycomb; ovules with parietal tissue [= crassinucellate], megaspore tetrad linear, functional megaspore single, chalazal, sporopollenin 0; gametophyte ± wholly dependent on sporophyte, development initially endosporic [apical cell 0, rhizoids 0, etc.]; male gametophyte with tube developing from distal end of grain, male gametes two, developing after pollination, with cell walls; embryo cellular ab initio, suspensor short-minute, embryonic axis straight [shoot and root at opposite ends], primary root/radicle produces taproot [= allorhizic], cotyledons 2; embryo ± dormant; chloroplast ycf2 gene in inverted repeat, trans splicing of five mitochondrial group II introns, rpl6 gene absent; ??whole nuclear genome duplication [ζ/zeta duplication event], 2C genome size (0.71-)1.99(-5.49) pg, two copies of LEAFY gene, PHY gene duplications [three - [BP [A/N + C/O]] - copies], 5.8S and 5S rDNA in separate clusters.
IID. ANGIOSPERMAE / MAGNOLIOPHYTA
Lignans, O-methyl flavonols, dihydroflavonols, triterpenoid oleanane, apigenin and/or luteolin scattered, [cyanogenesis in ANA grade?], lignin also with syringyl units common [G + S lignin, positive Maüle reaction - syringyl:guaiacyl ratio more than 2-2.5:1], hemicelluloses as xyloglucans; root cap meristem closed (open); pith relatively inconspicuous, lateral roots initiated immediately to the side of [when diarch] or opposite xylem poles; epidermis probably originating from inner layer of root cap, trichoblasts [differentiated root hair-forming cells] 0, hypodermis suberised and with Casparian strip [= exodermis]; shoot apex with tunica-corpus construction, tunica 2-layered; starch grains simple; primary cell wall mostly with pectic polysaccharides, poor in mannans; tracheid:tracheid [end wall] plates with scalariform pitting, multiseriate rays +, wood parenchyma +; sieve tubes enucleate, sieve plates with pores (0.1-)0.5-10< µm across, cytoplasm with P-proteins, not occluding pores of plate, companion cell and sieve tube from same mother cell; ?phloem loading/sugar transport; nodes 1:?; dark reversal Pfr → Pr; protoplasm dessication tolerant [plant poikilohydric]; stomata randomly oriented, brachyparacytic [ends of subsidiary cells ± level with ends of guard cells], outer stomatal ledges producing vestibule, reduction in stomatal conductance with increasing CO2 concentration; lamina formed from the primordial leaf apex, margins toothed, development of venation acropetal, overall growth ± diffuse, secondary veins pinnate, fine venation hierarchical-reticulate, (1.7-)4.1(-5.7) mm/mm2, vein endings free; flowers perfect, pedicellate, ± haplomorphic, protogynous; parts free, numbers variable, development centripetal; P = T, petal-like, each with a single trace, outer members not sharply differentiated from the others, not enclosing the floral bud; A many, filament not sharply distinguished from anther, stout, broad, with a single trace, anther introrse, tetrasporangiate, sporangia in two groups of two [dithecal], each theca dehiscing longitudinally by a common slit, ± embedded in the filament, walls with at least outer secondary parietal cells dividing, endothecium +, cells elongated at right angles to long axis of anther; tapetal cells binucleate; microspore mother cells in a block, microsporogenesis successive, walls developing by centripetal furrowing; pollen subspherical, tectum continuous or microperforate, ektexine columellate, endexine restricted to the apertural regions, thin, compact, intine in apertural areas thick, orbicules +, pollenkitt +; nectary 0; carpels present, superior, free, several, spiral, ascidiate [postgenital occlusion by secretion], stylulus at most short [shorter than ovary], hollow, cavity not lined by distinct epidermal layer, stigma ± decurrent, carinal, dry; suprastylar extragynoecial compitum +; ovules few [?1]/carpel, marginal, anatropous, bitegmic, micropyle endostomal, outer integument 2-3 cells across, often largely subdermal in origin, inner integument 2-3 cells across, often dermal in origin, parietal tissue 1-3 cells across, nucellar cap?; megasporocyte single, hypodermal, functional megaspore lacking cuticle; female gametophyte lacking chlorophyll, four-celled [one module, egg and polar nuclei sisters]; ovule not increasing in size between pollination and fertilization; pollen grains bicellular at dispersal, germinating in less than 3 hours, siphonogamy, pollen tube unbranched, growing towards the ovule, between cells, growth rate (ca 10-)80-20,000 µm h-1, tube apex of pectins, wall with callose, lumen with callose plugs, penetration of ovules via micropyle [porogamous], whole process takes ca 18 hours, distance to first ovule 1.1-2.1 mm; male gametophytes tricellular, gametes 2, lacking cell walls, ciliae 0, double fertilization +, ovules aborting unless fertilized; fruit indehiscent, P deciduous; mature seed much larger than fertilized ovule, small [<5 mm long], dry [no sarcotesta], exotestal; endosperm +, ?diploid [one polar nucleus + male gamete], cellular, development heteropolar [first division oblique, micropylar end initially with a single large cell, divisions uniseriate, chalazal cell smaller, divisions in several planes], copious, oily and/or proteinaceous, embryo short [<¼ length of seed]; plastid and mitochondrial transmission maternal; Arabidopsis-type telomeres [(TTTAGGG)n]; nuclear genome [2C] (0.57-)1.45(-3.71) [1 pg = 109 base pairs], ??whole nuclear genome duplication [ε/epsilon event]; ndhB gene 21 codons enlarged at the 5' end, single copy of LEAFY and RPB2 gene, knox genes extensively duplicated [A1-A4], AP1/FUL gene, palaeo AP3 and PI genes [paralogous B-class genes] +, with "DEAER" motif, SEP3/LOFSEP and three copies of the PHY gene, [PHYB [PHYA + PHYC]]; chloroplast IR expansions, chlB, -L, -N, trnP-GGG genes 0.
[NYMPHAEALES [AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]]: wood fibres +; axial parenchyma diffuse or diffuse-in-aggregates; pollen monosulcate [anasulcate], tectum reticulate-perforate [here?]; ?genome duplication; "DEAER" motif in AP3 and PI genes lost, gaps in these genes.
[AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]: phloem loading passive, via symplast, plasmodesmata numerous; vessel elements with scalariform perforation plates in primary xylem; essential oils in specialized cells [lamina and P ± pellucid-punctate]; tension wood + [reaction wood: with gelatinous fibres, G-fibres, on adaxial side of branch/stem junction]; anther wall with outer secondary parietal cell layer dividing; tectum reticulate; nucellar cap + [character lost where in eudicots?]; 12BP [4 amino acids] deletion in P1 gene.
[MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]] / MESANGIOSPERMAE: benzylisoquinoline alkaloids +; sesquiterpene synthase subfamily a [TPS-a] [?level], polyacetate derived anthraquinones + [?level]; outer epidermal walls of root elongation zone with cellulose fibrils oriented transverse to root axis; P more or less whorled, 3-merous [?here]; pollen tube growth intra-gynoecial; extragynoecial compitum 0; carpels plicate [?here]; embryo sac monosporic [spore chalazal], 8-celled, bipolar [Polygonum type], antipodal cells persisting; endosperm triploid.
[CERATOPHYLLALES + EUDICOTS]: ethereal oils 0 [or next node up]; fruit dry [very labile].
EUDICOTS: (Myricetin +), asarone 0 [unknown in some groups, + in some asterids]; root epidermis derived from root cap [?Buxaceae, etc.]; (vessel elements with simple perforation plates in primary xylem); nodes 3:3; stomata anomocytic; flowers (dimerous), cyclic; protandry common; K/outer P members with three traces, ("C" +, with a single trace); A ?, filaments fairly slender, anthers basifixed; microsporogenesis simultaneous, pollen tricolpate, apertures in pairs at six points of the young tetrad [Fischer's rule], cleavage centripetal, wall with endexine; G with complete postgenital fusion, stylulus/style solid [?here], short [<2 x length of ovary]; seed coat?; palaeotetraploidy event.
[PROTEALES [TROCHODENDRALES [BUXALES + CORE EUDICOTS]]]: (axial/receptacular nectary +).
[TROCHODENDRALES [BUXALES + CORE EUDICOTS]]: benzylisoquinoline alkaloids 0; euAP3 + TM6 genes [duplication of paleoAP3 gene: B class], mitochondrial rps2 gene lost.
[BUXALES + CORE EUDICOTS]: mitochondrial rps11 gene lost.
CORE EUDICOTS / GUNNERIDAE: (ellagic and gallic acids +); leaf margins serrate; compitum + [one position]; micropyle?; γ genome duplication [allopolyploidy, 4x x 2x], x = 3 x 7 = 21, 2C genome size (0.79-)1.05(-1.41) pg, PI-dB motif +; small deletion in the 18S ribosomal DNA common.
[ROSIDS ET AL. + ASTERIDS ET AL.] / PENTAPETALAE: root apical meristem closed; (cyanogenesis also via [iso]leucine, valine and phenylalanine pathways); flowers rather stereotyped: 5-merous, parts whorled; P = K + C, K enclosing the flower in bud, with three or more traces, C with single trace; A = 2x K/C, in two whorls, internal/adaxial to C, alternating, (numerous, but then usually fasciculate and/or centrifugal); pollen tricolporate; G [(3, 4) 5], whorled, placentation axile, style +, stigma not decurrent, compitum + [another position]; endosperm nuclear/coenocytic; fruit dry, dehiscent, loculicidal [when a capsule]; floral nectaries with CRABSCLAW expression; RNase-based gametophytic incompatibility system present.
[SANTALALES, CARYOPHYLLALES, SAXIFRAGALES, DILLENIALES, VITALES, ROSIDAE, [BERBERIDOPSIDALES + ASTERIDAE]]: ?
Phylogeny. For further discussion of relationships at the base of asterids and rosids, etc., see the Pentapetalae node.
Classification. Prior to the seventh version of this site asterids were part of a major polytomy that included rosids, Berberidopsidales, Santalales, and Caryophyllales, However, it seemed that the order of branching below the asterids seemed to be stabilizing, perhaps with a clade [Berberidopsidales [Santalales [Caryophyllales + Asterids]]], so the hierarchy was modified accordingly. Nevertheless, recent work (see above) indeed suggests that a polytomy is currently the best way to visualize relationships around here.
ROSIDS / ROSIDAE: anthers ± dorsifixed, transition to filament narrow, connective thin.
[ROSID I + ROSID II]: (mucilage cells with thickened inner periclinal walls and distinct cytoplasm); if nectary +, usu. receptacular; embryo long; chloroplast infA gene defunct, mitochondrial coxII.i3 intron 0.
ROSID I / FABIDAE / [ZYGOPHYLLALES [the COM clade + the N-fixing clade]]: endosperm scanty.
[the COM clade + the N-fixing clade]: ?
[OXALIDALES [CELASTRALES + MALPIGHIALES]] / the COM clade: seed exotegmic, cells fibrous.
[CELASTRALES + MALPIGHIALES]: ?
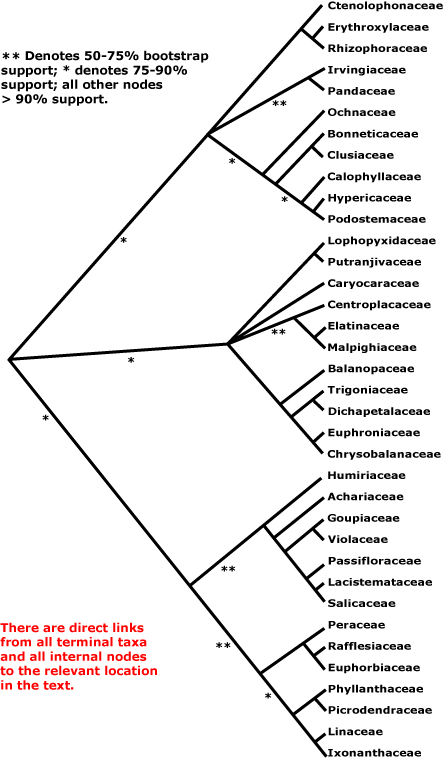
MALPIGHIALES Martius - Back to Main Tree.
Vessel element type?, (rays with multiseriate part no wider than uniseriate part); (sieve tubes with non-dispersive protein bodies); (stomata paracytic); (extra-floral nectaries +); lamina margin toothed; G often [3], stigma dry. - 36 families, 716 genera, 16,065 species.
Includes Achariaceae, Balanopaceae, Bonnetiaceae, Calophyllaceae, Caryocaraceae, Centroplacaceae, Chrysobalanaceae, Clusiaceae, Ctenolophonaceae, Dichapetalaceae, Elatinaceae, Erythroxylaceae, Euphorbiaceae, Euphroniaceae, Goupiaceae, Humiriaceae, Hypericaceae, Irvingiaceae, Ixonanthaceae, Lacistemataceae, Linaceae, Lophopyxidaceae, Malpighiaceae, Malesherbiaceae (= Passifloraceae-Malesherboideae), Medusagynaceae (= Ochnaceae-Medusagynoideae), Ochnaceae, Pandaceae, Passifloraceae, Phyllanthaceae, Picrodendraceae, Podostemaceae, Putranjivaceae, Quiinaceae (= Ochnaceae-Quiinoideae), Rafflesiaceae, Rhizophoraceae, Salicaceae, Trigoniaceae, Turneraceae (= Passifloraceae-Turneroideae), Violaceae.
Note: In all node characterizations, boldface denotes a possible apomorphy, (....) denotes a feature the exact status of which in the clade is uncertain, [....] includes explanatory material; other text lists features found pretty much throughout the clade. However, the precise node to which many characters, particularly the more cryptic ones, should be assigned is unclear. This is partly because homoplasy is very common, in addition, basic information for all too many characters is very incomplete, frequently coming from taxa well embedded in the clade of interest and so making the position of any putative apomorphy uncertain. Then there are the not-so-trivial issues of how character states are delimited and ancestral states are reconstructed (see above).
Age. Malpighiales may have begun to radiate some time in the Cretaceous-Late Aptian, some (119.4-)113.8(-110.7) or (105.9-)101.6(-101.1) Ma (Davis et al. 2005a; see also Xi et al. 2012b: table S7). Most other estimates are rather younger. The age of crown group Malpighiales was estimated as (93-)92, 90(-89) Ma, with Bayesian relaxed clock estimates slightly older, to 106 Ma (H. Wang et al. 2009), Wikström et al. (2001) suggested an age of (84-)81, 77(-74) Ma, Magallón and Castillo (2009) an age of ca 89.3 Ma, and Bell et al. (2010) an age of (97-)92, 89(-88) Ma. An age for [Salicaceae + Euphorbiaceae] of ca 82 Ma was suggested by t. Xue et al. (2020), the sister group of the malpigs was the N-fixing clade, the two diverging ca 105 Ma. (112.1-)111.1(-109.5) Ma is the crown group age in L. Cai et al. (2025b).
López-Martínez et al. (2023a: Table 3) noted that both Quadriplatanus georgianus (see Platanaceae) and Rariglanda jerseyensis (see Ericales) were strongly supported as being in Malpighiales in some analyses.
Evolution: Divergence & Distribution. The order contains ca 7.8% eudicot diversity (Magallón et al. 1999).
When the phylogeny of the group was considered to be rather like a starburst (see also below), the separation into the 15 or more clades that were then recognised, including individual families like Pandaceae, Caryocaraceae, Euphorbiaceae, Ochnaceae s.l., and Humiriaceae, was thought to have happened very rapidly in the late Aptian (Cretaceous), about 114-101 Ma (Davis et al. 2005a). Even with younger estimates for the age of the order, most families are thought to have diverged by the end of the Cretaceous (Wikström et al. 2001). Starbursts aside, current phylogenies still provide little support for the backbone of Malpighiales, which is akin to some sort of phylogenetic network (L. Cai et al. 2020), similarly, Cai et al. (2025b) suggested that some 7 clades had diverged within one million years of the origin of the order (ca 1/3 of the families they included), and more diverged soon afterwards. There may be a connection between diversification, whole genome duplications, and global climatic change around 56-54 Ma, i.e. the Palaeocene-Eocene Thermal Maximum 53.9 Ma, or (60.4-)56.8(-54.8) Ma (Cai et al. 2017/2018; Carretero-Paulet & van de Peer 2020).
Thus given the rather different relationships proposed by L. Cai et al. (2020, see also 2025b) and those suggested earlier, e.g. by Xi et al. (2012b), and the continued uncertainly over relationships here, further discussion on the evolution of Malpighiales is difficult, and the contents of the many of the Divergence & Distribution paragraphs on this page are particularly suspect.
Diversification rates in the order may have been moderately high (Magallón & Castillo 2009). Xi et al. (2012b: see different methods of analysis) examined diversification rates throughout the clade, and found about eight clades in which the rates of diversification decelerated and about five in which they accelerated; these are mentioned individually below.
Endress et al. (2013; see also Xi et al. 2012b in part) summarized floral variation in the order and found features potentially characterising most of the suprafamilial clades. Three-carpelate gynoecia occur in many families, articulated pedicels are also frequent, while paracytic stomata may characterise half of clade 2 below (Ochnaceae, etc.). Tokuoka and Tobe (2006) integrate testa anatomy and embryology with phylogeny. Furness (2011) looked at pollen development, focussing on the parietal-placentation clade; the massive amount of detail that she found is difficult to optimize on a tree, partly at least because of the sampling; Tao et al. (2018) discuss pollen evolution in the whole order. See also Kubitzki (2013a) for comments.
Ecology & Physiology. Malpighiales are particularly important in lowland tropical rainforests where they are a major component of the diversity, especially of the woody understory; they account for up to some 28% of the species and 38% of the total stems there (Davis et al. 2005a); members of Ericales (again, a clade with difficult relationships!), especially families like Sapotaceae and Lecythidaceae, are another major component of this vegetation. There are 10 families of Malpighiales with one or more species of the 227 species that make up half of all the trees with a d.b.h. 10 cm or more in Amazonian forests - all told, for a total of 43 species, i.e. about 20% (ter Steege et al. 2013).
Cretaceous diversification times for many of the clades in Malpighiales suggest that megathermal rainforest was developing then (see also Kubitzki 2013a), and obligately parasitic/mycoheterotrophic clades Rafflesiaceae s.l. (inc. Apodanthaceae) and Thismiaceae and even some Burmanniaceae may be of comparable ages. However, other evidence suggests that such forests may not have developed until early in the Caenozoic (Burnham & Johnson 2004, see Caenozoic Diversification), somewhat at odds with the dates just mentioned, however, Rafflesioideae are certainly and ancient clade (see below). Diversiification rate increases in the order are little nested - only 2/9 such events (Zuntini et al. 2024).
Plant-Animal Interactions. Caterpillars of outgroups to Nymphalidae-Nymphalinae and -Melitaeini, etc., are quite common on Malpighiales, especialy on families like Violaceae and relatives, Euphorbiaceae, etc. (Nylin & Wahlberg 2008; Nylin et al. 2013). The butterfly Cymothoë, with about 75 species, has hosts widely scattered in this order (Ackery 1988), although also found on Bignoniaceae (one species) and Rhamnaceae (sometimes another species). Phyllonorycter leaf-mining moths (Lepidoptera-Gracillariidae-Phyllocnistinae), with some 260 species and relatively common in more temperate regions, seem to have diversified here (and also especially Fagales) some time in the region of 50.8-27.3 Ma, well after the Malpighiales diversified, and after the genus itself evolved, some 76.3-50.3 Ma (Lopez-Vaamonde et al. 2006).
About a quarter of all records of extra-floral nectaries come from Malpighiales (Weber & Keeler 2013).
Genes & Genomes. Oginuma and Tobe (2010) provide the first chromosome counts for four families in the order. The intron in the atpF gene has been lost several times in Malpighiales, alone among angiosperms, however, this varies within Euphorbiaceae, Phyllanthaceae and Picrodendraceae (Daniell et al. 2008).
Chemistry, Morphology, etc.. Malpighiales may have closed root apical meristems (Clowes 2000), but the sampling is poor. Teeth with a single vein running into a congested and ± deciduous apex are common here - details of their distribution?
Endress et al. (2013) summarized the extensive and detailed morphological work that he and his collaborators have carried out on members of the order over the last twenty years and that of other workers; they emphasized that in 13 families (almost 1/3 of the families they recognized) ovules, etc., were largely unknown. Endress and Matthews (2006b) discuss petal appendages, etc., in the order. De Almeida et al. (2023b) draw attention to glands on the stamens in Malpighiales, although sampling is poor and the morphologies and products of these glands vary. Tobe and Raven (2011: see also supplement) provide an invaluable summary of embryological data for the whole order although, as they note, many families are poorly known; they plot the distribution of some characters of embryology and seed on a phylogenetic tree, much of which was, unfortunately, unresolved. For more on pollen morphology and development, see Furness (2012, 2013b) and Tao et al. (2018).
Phylogeny. Although Malpighiales are now strongly supported as being monophyletic (e.g. Davis et al. 2005a; Wurdack & Davis 2009; Xi et al. 2012b), relationships within them were initially poorly understood (e.g. Soltis et al. 2007a). Studies suggested relationships within particular clades of Malpighiales, e.g. Litt and Chase (1999), Schwarzbach and Ricklefs (2000), Chase et al. (2002), and Davis and Chase (2004), and these were in general agreement with relationships apparent in broader studies. The distinctive Lophopyxidaceae were placed close to Pandaceae (represented by Microdesmis) by Savolainen et al. (2000a; see also Chase et al. 2002); that relationship has held. Davis et al. (2005a) clarified some relationships in Malpighiales in a four-gene (all three compartments) analysis, in particular suggesting an association between the families with parietal placentation (and also Goupiaceae), and that Centroplacus (ex Euphorbiaceae s.l./Pandaceae) should be recognised as a separate family, perhaps sister to Ctenolophonaceae - support was weak (see also Korotkova et al. 2009 and Soltis et al. 2011 for relationships in Malpighiales). Indeed, in a molecular study by Wurdack et al. (2004) Centroplacus had been associated with Pandaceae, although with very little support. However, Ctenolophonaceae were linked with Erythroxylaceae and Rhizophoraceae, while Bhesa, ex Celastraceae, which L.-B. Zhang and Simmons (2006) found fell among the few Malpighiales they included in their analysis of Celastrales, linked with Centroplacaceae, etc. (Wurdack & Davis 2009). A [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Chrysobalanaceae + Euphroniaceae]]] clade had strong support, e.g. Davis et al. (2005a), Tokuoka and Tobe (2006) and Korotkova et al. (2009). Linaceae had been weakly associated with Picrodendraceae in Chase et al. (2002a), but it was linked with Irvingiaceae in Tokuoka and Tobe (2006). It had been suggested that Malpighiaceae were rather weakly associated with Peridiscaceae and were perhaps near Clusiaceae et al. (Chase et al. 2002); for the current position of Peridiscaceae, here in Saxifragales, see e.g. Davis and Chase (2004).
Some families have been particularly peripatetic. Irvingia was sister to Erythroxylum in a tree presented by Fernando et al. (1995), and the stipules of Irvingiaceae, Erythroxylaceae and Ixonanthaceae are indeed similar (Weberling et al. 1980). However, Irvingiaceae were weakly associated with Putranjivaceae in Chase et al (2002a) and with Linaceae (close) in Davis et al. (2005a). See Clade 2 below for current ideas of the relationships of Irvingia. Recent nuclear genome analyses (see below) suggest that yet other taxa are peripatetic, Linaceae themselves (or at least Linum) and in particular Podostemaceae, for example.
In 2011 there were nine clades composed of two or more families in Malpighiales along with seven separate families that together formed a very substantial polytomy (Davis et al. 2005a; Wurdack & Davis 2009; Xi et al. 2010; Soltis et al. 2011). M. Sun et al. (2016) and Z.-D. Chen et al. (2016) recovered little in the way of major groupings within Malpighiales. However, relationships in Xi et al. (2012b) were rather more resolved. The major analysis in this study used 78 protein-coding plastome genes and four ribosomal genes; families not included were Lophopyxidaceae, Malesherbiaceae and Rafflesiaceae (the last-named for obvious reasons); other analyses had included many more taxa but less complete sampling of genes (see Xi et al. 2012b for details). Malpighiales were divided into three main clades, the Salicaceae-Euphorbiaceae, Rhizophoraceae-Clusiaceae, and Malpighiaceae-Chrysobalanaceae clades (clades 1, 2 and 3 below), all with substantial molecular support (>80% ML bootstrap, 1.0 p.p.) and even with a modicum of morphological support. Although at the next level of the tree both clades 2 and 3 have polytomies and clade 1 an only weakly-supported dichotomy, overall the improvement of resolution in the tree seemed to be quite substantial (Xi et al. 2012b), and the relationships they suggested are largely followed below. H.-T. Li et al. (2019, 2021) recovered the three clades, although both they and groupings within them are for the most part poorly (or worse) supported, exceptions being relationships in the clusioid, violoid, rhizophoroid and chrysobalanoid clades in particular.
Clade 1. [[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]] [[Peraceae [Rafflesiaceae + Euphorbiaceae]] [[Phyllanthaceae + Picrodendraceae] [Linaceae + Ixonanthaceae]]]] - see below.
This is the one clade that was for the most part recovered by M. Sun et al. (2016). Although support for the [Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]] clade is not strong (Xi et al. 2012b), the part of this clade excluding Humiriaceae (= the parietal clade) has very strong support. Goupiaceae are certainly to be included here, although their association with Violaceae is only weakly supported, as is the position of the combined clade; major relationships in the rest of this clade have strong support (Xi et al. 2012b).
Molecular evidence that a group of families with parietal placentation and (often) three carpels was monophyletic had initially not been compelling (e.g. see Savolainen et al. 2000a; Chase et al. 2002), although part of the rpS 16 gene is absent from Passifloraceae-Passifloroideae and -Turneroideae, Violaceae, and Salicaceae s. str. (and also Linaceae and Malpighiaceae, so really a feature of Malpighiales?: see Downie & Palmer 1992). Salicaceae were weakly associated with Passifloraceae, and in turn with Humiriaceae and Pandaceae, and Violaceae were weakly associated with Achariaceae (and Goupiaceae, Lacistemataceae and Ctenolophonaceae) in Chase et al (2002). Tokuoka and Tobe (2006) found a weakly-supported relationship between the Passifloraceae group and Violaceae (see also Soltis et al. 2007a), and strongly supported relationships between Lacistemataceae and Salicaceae. However, Davis et al. (2005a) found a poorly/moderately supported association of these taxa with parietal placentation (59% bootstrap, 1.00 posterior probability), and also Goupiaceae, with axile placentation, and a similar grouping is also evident in e.g. Wikström et al. (2001), Wurdack and Davis (2009), Korotkova et al. (2009: 83% jacknife, 1.00 pp, Goupiaceae not included), Soltis et al. (2011: details of relationships unclear) and M. Sun et al. (2016). Ixonanthes was rather surprisingly embedded in Achariaceae in the Bayesian analysis of Soltis et al. (2007a), but that was due to misidentification of the material, which was a species of Hydnocarpus (K. Wurdack, pers. comm.).
Indeed, classical morphological studies had long suggested that there was a group that included Salicaceae, Achariaceae, Violaceae, Flacourtiaceae, and Passifloraceae and its segregates, Malesherbiaceae and Turneraceae, in part because of their common possession of parietal placentation, some sort of corona or scales in the flower, nectaries outside the stamens, etc. (e.g. Cronquist 1981). However, a number of other families now known to be quite unrelated, several now in Cucurbitales, were also included. Interestingly, species of the old Flacourtiaceae had one of two kinds of seed coat: the exotegmen was either more or less fibrous - taxa with this kind of exotegmen are now mostly in Salicaceae - or massive and non-fibrous - taxa with this exotegmen are now in Achariaceae (Corner 1976). It was also commonly recognized that Salicaceae were simply an extreme morphology reflecting the wind pollination common in that family, and that they could be linked with some of the old Flacourtiaceae. Distinctive cyclopentenoid cyanogenic glucosides and/or cyclopentenyl fatty acids, including gynocardin, also occur sporadically here (Webber & Miller 2008). The inclusion of Goupiaceae in this clade is the only real surprise since it is morphologically rather distinct.
The other weakly supported clade in Clade 1 is [[Peraceae [Rafflesiaceae + Euphorbiaceae]] [[Phyllanthaceae + Picrodendraceae] [Linaceae + Ixonanthaceae]]]], the euphorbioids. This is an unexpected clade in that the fruits of a rather broadly delimited Euphorbiaceae (inc. both Phyllanthaceae and Putranjivaceae) are very distinctive, with the walls falling away leaving the persistent columella, and that is one of the main characters that I use to recognize herbarium material of the extended family. It is hardly surprising that Merino Sutter and Endress (1995) argued for a broad circumscription of the family. Within Clade 1 the [[Phyllanthaceae + Picrodendraceae] [Linaceae + Ixonanthaceae]] and [Peraceae [Rafflesiaceae + Euphorbiaceae]] clades are strongly supported (Xi et al. 2012b). Note that M. Sun et al. (2016) recovered only part of this clade, within which Euphorbiaceae s. str. are paraphyletic while the placement of Irvingiaceae was unexpected. The strongly-supported relationships of the few members of this clade studied by L. Zhou et al. (2016) are incompatible with those below.
Determining the phylogenetic relationships of Rafflesiaceae is difficult. The floral morphologies of Rafflesiaceae, and also Cytinaceae, Apodanthaceae and Mitrastemonaceae, families that have often been associated with Rafflesiaceae, are distinctive, yet this morphology is often hard to interpret, furthermore, molecular analyses have been problematic in part because of the very long branches in some genes and the general problem of obtaining suitable sequences from holoparasites (e.g. see results from analysing sequences of the mitochondrial atp1 gene - Nickrent et al. 2004a). Indeed, when representatives of all four families were in the same analysis, an apparently monophyletic Rafflesiales could be recovered (Nickrent et al. 2004a). Nickrent (2002) had suggested that Rafflesiaceae themselves might be close to Malvales, however, in other analyses in which not all these erstwhile Rafflesiales were included their break-up seemed to be on the cards (see also Barkman et al. 2004; Davis & Wurdack 2004; Nickrent et al. 2004a; Davis et al. 2005a; Filipowicz & Renner 2010).
However, a Rafflesiaceae that includes Apodanthaceae does seem now to be firmly placed in Malpighiales - but where? Barkman et al. (2004) sequenced the mitochondrial gene, matR, of Rafflesia and found a strong association with Malpighiales (see below for previous placements of Rafflesiaceae). Although sampling within Malpighiales (only three taxa with parietal placentation were included) and other rosids was poor, Barkman et al. (2004) thought that the flowers of Rafflesia could be interpreted as having a number of features in common with those of Passifloraceae, including a corona (called a diaphragm by students of Rafflesiaceae), androgynophore, parietal placentation (although this is common in other echlorophyllous parasites), etc., but as Nickrent et al. (2004a) pointed out, the basic similarity of these structures needs careful examination. Davis and Wurdack (2004: two nuclear, one mitochondrial [matR] genes), with considerably more extensive sampling, confirmed the inclusion of Rafflesiaceae in Malpighiales, favouring a position closer to Ochnaceae, Clusiaceae and their relatives. Although tenuinucellate ovules are common there, it is quite common for holoparasitic taxa to lack parietal tissue in their ovules. A position of Rafflesiaceae in or near Malpighiales was common in the analyses described by Nickrent et al. (2004a). Davis et al. (2007), using largely mitochondrial genes, exemplars of all families of Malpighiales, three of the four genera of Peraceae, Chaetocarpus only excluded, and several Euphorbiaceae, including Cheilosioideae, placed Rafflesiaceae within Euphorbiaceae and with quite good support (see also Wurdack & Davis 2009). In a wrinkle on this association of Rafflesiaceae with Euphorbiaceae, M. Sun et al. (2016) found Rafflesiaceae to be sister to Neoscortechinia and Cheilosa, here sister to the rest of Euphorbiaceae s. str., all other members of the latter family being sister to the combined clade, while Z.-D. Chen et al. (2016: ?sampling, low support) obtained the relationships [Rafflesiaceae [Peraceae + Euphorbiaceae]], a grouping that was recognised here for some time. Of course, given the recent association of Rafflesiaceae with Apodanthaceae and the possible movement of this clade away from Euphorbiaceae (see below), Peraceae can now safely be made to disappear into Euphorbiaceae, whence they came.
Although the relationships of the holoparasitic Apodanthaceae had for some time been unclear (e.g. they were unplaced in A.P.G. 2009), a link with Rafflesiaceae does now seem to be on the cards. Nickrent et al. (2004) had suggested a position for Apodanthaceae either within Malvales (especially in the three-gene analyses and that of nuclear SSU rDNA), or in or near Cucurbitales (analysis of matR), but they inclined to the former position. Barkman et al. (2007: support weak, but rather comprehensive analysis) also suggested the latter position; the mitochondral genes cox1 and matR showed massive divergence, but not the atp1 gene (Barkman et al. 2007). Additional molecular analyses (D. Nickrent, pers. comm.; esp. Filipowicz & Renner 2010) supported the inclusion of Apodanthaceae in Cucurbitales. This is consistent with their dioecy, extrose anthers, more or less inferior ovary and parietal placentation, all features common in Cucurbitales (see also Filipowicz & Renner 2010), but all these features are common in parasitic plants in general (Renner & Ricklefs 1995). There were also a number of codon subsitutions in common between Apodanthaceae and Cucurbitales (Barkman et al. 2007; Filipowicz & Renner 2010). The exact position of the family in Cucurbitales was not certain, the relationships suggested with the morphologically rather different (but apomorphically so) Corynocarpaceae and Coriariaceae being only weakly supported, and Apodanthaceae were on a very long branch (Filipowicz & Renner 2010; see also M. Sun et al. 2016). The situation remained the same in Bellot and Renner (2014b), and in trees used when estimating substitution rates Apodanthaceae linked either with a clade [Anisophylleaceae + Corynocarpaceae] or a clade including the whole of the rest of the family, but with a rather different topology to that used here; other topologies were also obtained, although none with strong support. Schaefer and Renner (2011b) had found that Apodanthaceae associated with Oxalidales in nuclear analyses, but with Cucurbitales when mitochondrial data were used. F. González and Pabón-Mora (2017b: see Table 2) compared Apodanthaceae with other Cucurbitales, again noting characters like parietal placentation, inferior ovary and numerous ovules that they had in common with some of the rest of the order. Apodanthaceae were to be found within Cucurbitales in these pages for quite some time.
However, further changes are under way. Thus Apodanthaceae in the Kew Tree of Life (version ix.2024) were to be found by themselves, being between Caryophyllales and [Gunnerales + Dilleniales], but with very low support, although in Zuntini et al. (2024) they are placed near basal in Malpighiales, relationships there being [Ixonanthaceae [Apodanthaceae [all other Malpighiales...]]], if again with little support. As mentioned, Alzate et al. (2024: 55 species, 12 orders, 1011 single-copy nuclear genes) found that Apodanthaceae were well supported as being sister to Rafflesiaceae. This clade was sister to the other Malpighiales in their study (i.e. Euphorbiaceae, Hypericaceae, Salicaceae, Linaceae, etc.); other taxa in the analysis included members of the N-fixing clade, Brassicales, Malvales, Sapindales and Santalales. In a rDNA analysis by Alzate et al. (2024) [Apodanthaceae + Rafflesiaceae] were again included within Malpighiales, although several basal branches there were rather short and [A. + R.] are almost part of a polytomy. Similarly, L. Cai et al. (2025b) found [A. + R.] to be sister taca, but whether sister to [Euphorbiaceae + Peraceae], Putranjivaceae, Pandaceae, or even [Erythroxylaceae + Rhizophoraceae] was unclear. Cai et al. (2025b: 45 taxa, inc. 3 outgroups, 2135, 829 or 446 genes) had looked at relationships here and noted that 98.9% of the genes that they examined were uninformative when it came to placing A. and R., half of rhe genes that are normally conserved had been lost - this included housekeeping genes - there had been extensive HGT, there were high substitution rates and very A-T rich genomes; concatenation analyses tended to cause problems. Apodanthaceae and Rafflesiaceae inhabited the phylogenetic danger zone of excessive ILS and long branch attraction (Cai et al. 2025b). Rafflesiaceae s.l. are placed basally in Malpighiales below, although this position is clearly not fixed.
Clade 2. [[Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]], [Irvingiaceae + Pandaceae], [Ochnaceae [[Clusiaceae + Bonnetiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]]] - see below.
Weak support for an association of [Caryocaraceae [Linaceae + Irvingiaceae]] with [Rhizophoraceae + Erythroxylaceae] (Soltis et al. 2007a), has not been strengthened, although they have a number of features in common, such as a basally connate androecium, epitropous ovules with an endothelium, etc. (Matthews & Endress 2007). Although Ctenolophonaceae, etc., might also be associated, their floral similarities did not seem to be so great. However, Wurdack and Davis (2009) found support for the clade [Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]], but further relationships were unclear. Erythroxylaceae are commonly well supported as sister to Rhizophoraceae (e.g. Setoguchi et al. 1999; Schwarzbach & Ricklefs 2000; Chase et al. 2002; Korotkova et al. 2009). In the study by Xi et al. (2012b), the clade [Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]] (= rhizophoroids) had strong support (see also M. Sun et al. 2016), [Pandaceae + Irvingiaceae] (= pandoids) had rather weak support (64% ML bootstrap, 0.97 PP). Centroplacus was sister to Pandaceae, but with little support (Wurdack et al. 2004); for Centroplacus, see Clade 3 below.
Relationships within the clusioid clade, [Ochnaceae [[Clusiaceae + Bonnetiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]]], were initially unclear (see also Soltis et al. 1999b; Gustaffson et al. 2002; Davis et al. 2005b), and the clade had only weak support (70% ML bootstrap, 0.81 p.p.) in Xi et al. (2012b), but its composition is consistent with morphology; Ochnaceae and Clusiaceae et al. also have a generally similar flavonoid spectrum (Hegnauer 1990). Wurdack and Davis (2009), M. Sun et al. (2016) and particularly Ruhfel et al. (2011, 2013, see also Xi et al. 2012b) have confirmed the paraphyly of the old Clusiaceae, and support is generally quite strong, although that for the [Bonnetiaceae + Clusiaceae] clade is the weakest (Xi et al. 2012b; Ruhfel et al. 2013), while the branch leading to Podostemaceae is rather long. Meseguer et al. (2014a: nuclear markers) found that Podostemon, the only Podostemaceae they examined, was sister to Vismia, i.e. Hypericaceae were paraphyletic, although support was not strong, and in H.-T. Li et al. (2019) a clade [Erythroxylaceae + Rhizophoraceae] was embedded in the clusioid clade, but support was poor. In the 21-taxon plastome analysis of Trad et al. (2021), Podostemaceae found a place sister to Hypericaceae without much fuss, but the position of Bonnetiaceae was unclear - either sister to Clusiaceae, or sister to all other clusioids (Ochnaceae are not in their Fig. 5). In an analysis of two different data sets, both yielding the first topology just mentioned, very few gene trees supported it - or the clade [Calophyllaceae [Hypericaceae + Podostemaceae]], for that matter. Clearly, the position of Bonnetiaceae is uncertain (see also Li et al. 2021), and for more on the position of Podostemaceae, see at the end of this whole section.
Clade 3. [[Lophopyxidaceae + Putranjivaceae], Caryocaraceae, [Centroplacaceae [Elatinaceae + Malpighiaceae]], [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Chrysobalanaceae + Euphroniaceae]]]] - see below.
Although this clade has strong support in Xi et al. (2012b), relationships within it are still poorly understood. The [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Chrysobalanaceae + Euphroniaceae]]] and [Putranjivaceae + Lophopyxidaceae] clades (= chrysobalanoids and putranjivoids respectively) are well supported (see also M. Sun et al. 2016), but the [Centroplacaceae [Elatinaceae + Malpighiaceae]] clade (malpighioids) has poor support. The position of the distinctive Caryocaraceae is unclear, althougth there is little question that it belongs here (Xi et al. 2012b). There was some support for Picrodendraceae as sister to the chrysobalanoids in Soltis et al. (2007a: as Pseudanthaceae, Phyllanthaceae not included), but this relationship has not been confirmed.
However, these relationships are hardly written in stone. The topology of the tree in Cai et al. (2017/18) shows substantial differences, although details of relationships in the order were not their focus. The Ochnaceae-clusioid clade, taxa with parietal placentation, [Rhizophoraceae + Erythroxylaceae] and [Elatinaceae + Malpighiaceae] were the main groupings that they recovered that are consistent with those just discussed. The euphorbioid group in particular did not hold together, both Phyllanthaceae and Linaceae being separate from Euphorbiaceae, although perhaps perversely Drypetes (Putranjivaceae), once included in Euphorbiaceae but then widely separated, was sister to Euphorbiaceae (Cai et al. 2017/18). Along the same lines, de Santana Lopes et al. (2017) in an analysis of 63 plastid protein-coding genes from 38 taxa found that Linaceae were sister to Chrysobalanaceae, while in a 76 protein-coding plastome analysis of Nunes et al. (2020b), relationships were [Caryocaraceae [[Euphorbiaceae + Chrysobalanaceae] [[Malpighiaceae [Clusiaceae + Erythroxylaceae]] [[Putranjivaceae + Linaceae] [the parietal clade]]]] - families like Linaceae currently seem to have no fixed abode. Most recently the comprehesive angiosperm plastome analysis by H.-T. Li et al. (2021) not surprisingly largely recovers the relationships being discussed.
Cai et al. (2020) returned to the problem, noting that Malpighiales had included nine of the top ten most destabilizing clades in seed plants in an earlier study by S. A. Smith et al. (2013: Figs 2-4), and found that part of Malpighiales - Caryocar, Ctenolophon, Klainedoxa, also the Euphorbiaceae area, etc. - were in an utter mess. Cai et al. (2020) looked at 423 single-copy nuclear loci in 64 taxa (56 in Malpighiales) which belonged to 33 of the 36 families recognized in A.P.G. IV (2016), Rafflesiaceae, Podostemaceae and Irvingiaceae not being included. The result? "[R]ecalcitrant phylogenetic relationships like the backbone of Malpighiales may be better represented as phylogenetic networks." Indeed, the backbone of their tree has very little support, and Cai et al. (2020) noted ten separate groupings along it. Seven are mentioned below, the other three being made up of members of single families, Ochnaceae, Caryocaraceae and Ixonanthaceae. Proceeding up the tree, the six groups are [Linaceae [Hypericaceae [Calophyllaceae + Bonnetiaceae]]] (group 10 of Cai et al.), [Elatinaceae* + Malpighiaceae*] (group 8), [Erythroxylaceae + Rhizophoraceae] (group 7), [Phyllanthaceae + Picrodendraceae] (group 4), [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]] (group 3: the chrysobalanoids below), [Violaceae [Passifloraceae* [Goupiaceae [Achariaceae* [Lacistemataceae + Salicaceae*]]]]] (group 2: the parietal clade) and [[Lophopyxidaceae + Putranjivaceae] [[Peraceae* + Euphorbiaceae*] [Centroplacaceae* [Ctenolophonaceae [Humiriaceae* + Pandaceae*]]]]] (group 1) (Cai et al. 2020) - families in which more than one species was sampled have a star. Looking at simulations of gene tree variation, some 57.5% of this variation is made up of three elements, of which gene tree estimation error makes up 52%, gene flow/introgression as measured by a Reticulation Index, 32%, and independent lineage sorting, 15% (Cai et al. 2020: Fig. 5). Note that groups 1 and 2 perhaps could be combined, however, although the combined group had very strong support, it did have a very high reticulation index, and this is also high along the spine of Malpighiales (Cai et al. 2020). Of the three major clades mentioned by Xi et al. (2012b) above, Clade 1 is the only one that is even barely close to being recognized by Cai et al. (2020), nevertheless, relatives of [Peraceae + Euphorbiaceae], part of Clade 1, are scattered through the rest of their tree. Furthermore, details of relationships within groups of any size that are recognised both by Cai et al. (2020) and those in APWeb (as of ii.2021, which largely followed Xi et al. 2012b), differ substantially, except for those in the chrysobalanoids. However, support values for relationships in the chrysobalanoids are high in both studies, while those in Violaceae and relatives and Clusiaceae and relatives are definitely lower. Relationships along the spine of Malpighiales in the plastome analysis of She et al. (2019/2020) were rather poorly supported.
Podostems fitted without any obvious difficulty into the clusioid clade in the whole plastome analysis of Trad et al. (2021), although they sometimes wander outside Malpighiales. A whole genome analysis that focused on Cladopus chinensis yielded the relationships [Brassicales [Podostemales [[Malpighiales [Rosales + Cucurbitales]]]], although an analysis of 91 single-copy genes returned the more conventional relationships of [Podostemaceae [Salicaceae + Euphorbiaceae]] (few malpigs sampled: T. Xue et al. 2020; c.f. Fig. 4a, 4b). Xue et al. (2020) provided dates for the first set of relationships (thus the Cladopus clade diverged ca 106 Ma), but they preferred the second set. W. J. Baker et al. (2021a) also found Podostemaceae to be quite separate from other Malpighiales, being in an isolated clade much more basal in rosids. This position was more or less maintained in the Seed Plant Tree of Life i.2022 release, however, Tristichoideae were quite separate from Podostemoideae, the former being immediately basal to a clade containing Huaceae, Celastrales and Malpighiales, while Zehnderia microgyna (Podostemoideae), along with one or two other miscellaneous taxa, was immediately basal to Gentianales (Weddellina was not sampled).
No changes to relationships here had been made as of 5.2021, and the relationships recognized then were largely those of Xi et al. (2012b); only in ix.2024 were things reviewed... The old classification was Clade 1 of Xi et al. (2012b) = [[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]]] [[Peraceae [Rafflesiaceae + Euphorbiaceae]] [[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]]] Clade 2 of Xi et al. (2012b) = [[Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]], [Irvingiaceae + Pandaceae], [Ochnaceae [[Bonnetiaceae + Clusiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]] Possible Clade - note, includes Clade 1 = [[[Lophopyxidaceae + Putranjivaceae], Caryocaraceae, [Centroplacaceae [Elatinaceae + Malpighiaceae]], [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]]] [[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]]] [[Peraceae [Rafflesiaceae + Euphorbiaceae]] [[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]]]. Relationships within Malpighiales found by She et al. (2019) in an analysis using plastome data were only poorly supported; broader relationships were [Vitales [Saxifragales [[Geraniales + Myrtales] [Sapindales [Huerteales [Brassicales + Malvales]]]] [Zygophyllaes [N-fixing clade [Malpighiales [Oxalidales + Celstrales]]...
More extensive nuclear genome analyses have resulted in substantial movements of families/re-establishment of polytomies. Relationships in W. J. Baker et al. (2021a: see Seed Plant Tree) in an early Angiosperms 353analysis are interesting. Those within Ochnaceae, where sampling was good, are very much those in the treatment below, and the chrysobalanoids are also recovered, however, Linaceae (Linum only sampled) are sister to all other Malpighiales and Phyllanthaceae are polyphyletic, while group 10 (sort of) above is [Bonnetiaceae [Clusiaceae [Calophyllaceae + Hypericaceae]]], group 2 is there, but with different relationships, and so on. A recent nuclear analysis is that of Zuntini et al. (2024), although note that half the nodes along the backbone of the Malpghiales tree have rather low posterior probabilities (see asterisks), although relationships along the branches tend to be better supported. [Ixonanthaceae [[Ochnaceae [Bonnetiaceae [Clusiaceae [Calophyllaceae [Hypericaceae + Podostemaceae]]]]] *[*[Euphroniaceae + Rafflesiaceae] [[Elatinaceae + Malpighiaceae] *[*[Caryocaraceae [Picrodendraceae + Phyllanthaceae]] *[[Peraceae + Euphorbiaceae] *[*Irvingiaceae + Linaceae] *[*[[Erythroxylaceae + Rhizophoraceae] [Balanopaceae [Chrysobalanaceae [Dichapetalaceae + Trigoniaceae]]]] *[Centroplaceae *[[Lophopyxidaceae + Putranjivaceae] *[*[Ctenolophonaceae [Humiriaceae + Pandaceae]] [Achariaceae + Goupiaceae] [[Passifloraceae + Violaceae] *[Salicaceae + Lacistemaceae]]]]]]]]]]]]]]. Apodanthaceae (now included in Rafflesiaceae, see below) are weakly supported as being sister to all Malpighiales bar Ixonanthaceae.
On the other hand, relationships suggested by Y.-L. Qiu et al. (2024) are very different: [[Balanopaceae + Chrysobalanaceae] [[Salicaceae [Passifloraceae + Violaceae] [Ixonanthaceae + Achariaceae]]] [[Irvingiaceae + Ctenolophonaceae] [Centroplacaceae [Linaceae + Phyllanthaceae]]] [[Humiriaceae + Putranjivaceae] [Rhizophoraceae [[Malpighiaceae + Elatinaceae] [Euphorbiaceae [Hypericaceae [Clusiaceae + Bonnetiaceae]]]]]]]]]]. The sampling is slight, and although all nodes were dated, that means little given the spread in ages for many of the nodes below, the relationships Qiu et al. (2024) recovered, and the fact that their analysis was based on only five organellar and nuclear genes. In any event, there is no point in getting worked up about details of relationships here; Ericales and Malpighiales are perhaps the two largest orders in which relationships are still poorly understood.
Classification. See Kubitzki (2013a) for a summary. A.P.G. (1998) thought that it would be useful to adopt a narrow circumscription for families that used to be included in Flacourtiaceae and Euphorbiaceae s.l. (both in Clade 1 above) since the composition of the clades that were even then apparent were quite different from those in previous classifications. Indeed, the realignments caused by the break-up of the old Flacourtiaceae and the integration of the fragments with Salicaceae and Achariaceae correlate well with a number of morphological and anatomical characters (Wurdack & Davis 2009). These earlier decisions were compatible with the tree in Xi et al. (2012b) and are also compatible with that in Cai et al. (2020). However, given the relationships in Xi et al. (2012b), to restore Euphorbiaceae to close to its old circumscription would have required the inclusion of Peraceae, Picrodendraceae, Phyllanthaceae, Linaceae, Ixonanthaceae and Rafflesiaceae-Rafflesioideae, making a very heterogeneous and perplexing group. Moreover, given the recent and still developing ideas of relationships just mentioned, any Euphorbiaceae s.l. would be equally heterogeneous. On the other hand, the largely plastid-based relationships in the clusioid group necessitated the break-up of the old Clusiaceae/Guttiferae (see also A.P.G. 2003, 2009, 2016), but to restore it would have meant also including Podostemaceae and Bonnetiaceae, and here plastid- and nuclear-based relationships are similar. It will be interesting to see if nuclear analyses suggest any changes to the circumscriptions of families.
Previous Relationships. The history of the circumscription and putative relationships of the small family Ixonanthaceae, here sister to Linaceae (Clade 1, with strong support), is an example of problems taxonomists have faced in circumscribing major groups in this whole area, and in justifying relationships - yes, there are distinctive characters, but which reliably indicate relationships? Ixonanthaceae have previously been associated with several different families, although Van Hooren and Nooteboom (1988) noted that they had often been linked to Linaceae. Thus Robson and Airy Shaw (1962) thought that the "spiral convolution of the filaments and style" of Cyrillopsis (Ixonanthaceae) was a point of similarity between this genus and Irvingiaceae (Clade 2). Some species of Ochthocosmus (Ixonanthaceae) also have flowers very similar to those of Cyrillopsis, with the thin calyx reflexed after anthesis (Ixonanthes), while other species of Ochthocosmus have persistent, erect, almost scarious-looking sepals, as is common in Linaceae. Allantospermum has flowers very similar to those of Cyrillopsis, and its relationships have presented particular problems, the genus seeming to be intermediate between the Ixonanthes and Irvingia groups. Forman (1965) placed it with the former group, but both groups, he thought, were subfamilies in Ixonanthaceae, while Nooteboom (1967) placed it in the latter group, but here the two groups were subfamilies of Simaroubaceae. Pollen suggested to Oltmann (1971) that Allantospermum was in Ixonanthaceae, Irvingiaceae were not related. Takhtajan (1997) included Allantospermum in Irvingiaceae, close to Simaroubaceae, while Ixonanthaceae were in Rutales. Cronquist (1981) included Irvingia in Simaroubaceae-Sapindales while Ixonanthaceae were in Linales. Bove (1997: morphological phylogenetic analysis), on the other hand, suggested that Ixonanthaceae and Humiriaceae (also Clade 1, but not immediately related) were sister taxa, both having ellagic acid, a "free" annular nectary encircling the ovary, and an entire stigma. In the context of Linales (also including Linaceae, Hugoniaceae, Erythroxylaceae [also Clade 2, not immediately related to Irvingiaceae]: see Cronquist 1981), Ixonanthaceae were rather different in their free stamens, semi-inferior ovaries and pollen grains with supratectal spines (Bove 1997). Byng et al. (2016) link Cyrillopsis with Irvingiaceae, a position with which morphology and chemistry are in general agreement.
Synonymy: Linineae Shipunov, Rhabdodendrineae Shipunov, Rhizophorineae Shipunov - Balanopales Engler, Chailletiales Link, Chrysobalanales Link, Elatinales Martius, Erythroxylales Link, Euphorbiales Berchtold & J. Presl, Flacourtiales Martius, Garciniales Martius, Homaliales Martius, Hypericales Berchtold & J. Presl, Irvingiales Doweld, Lacistematales Martius, Linales Berchtold & J. Presl, Malesherbiales Martius, Marathrales Dumortier, Medusagynales Reveal & Doweld, Ochnales Berchtold & J. Presl, Pandales Engler & Gilg, Passiflorales Berchtold & J. Presl, Phyllanthales Doweld, Podostemales Lindley, Rafflesiales Martius, Rhizophorales Berchtold & J. Presl, Salicales Lindley, Samydales Berchtold & J. Presl, Sauvagesiales Martius, Scyphostegiales Croizat, Stilaginales Martius, Turnerales Link, Violales Berchtold & J. Presl - Euphorbianae Reveal, Ochnanae Doweld, Podostemanae Reveal, Rafflesianae Reveal, Rhizophoranae Reveal & Doweld, Violanae Reveal - Malpighiopsida Bartling, Passifloropsida Brongniart, Podostemopsida G. Cusset & C. Cusset, Salicopsida Bartling, Violopsida Brongniart
RAFFLESIACEAE Dumortier, nom. cons. - Back to Main Tree
Holoparasitic, plant endophytic, gravitropism 0; plant mycelium-like, typical histological organization 0 [e.g. root and shoot apical meristems 0, roots, stems and leaves 0]; haustoria 0, few-celled tapered extensors [= sinkers] directly connect the endophyte with host vasculature; vessel elements +; cells with very large nuclei, vacuoles small/0, cytoplasm dense; chloroplasts/photosynthesis 0; flowers unisexual, single, developing directly from endophyte in host parenchyma, reproductive apex forming via schizogeny, cap-like structure surrounds floral primordium; P imbricate, with stomata, innermost members valvate; nectary +, at base of synandrium / style/gynostemium / base of ovary wall, papillate [more obviously in staminate flowers]; staminate flowers: synandrium +, 1-4-whorled, A extrorse, athecal, filaments 0, pollen sacs globose to ovoid, adnate to central column [= pistillode, i.e. gynostemium], dehiscence extrorse, transverse, fibrous endothecium 0; carpelate flowers: central column with large vesicular papillae; G ± inferior, placentation parietal, stigma large, ± hemispherical/disc-shaped; ovules bitegmic, no vascular tissue in funicle, parietal tissue 0; fruits baccate; seeds many, small; embryo small [25> cells], undifferentiated; plastid trnE 0.
5/57 [list]. Tropical, rather scattered, not Madagascar, India-China, Papuasia, etc..
Age. The crown-group age of Rafflesiaceae is (108.7-)106.5(-102.8) or ca 98.8 Ma (L. Cai et al. 2025b).
1. Rafflesioideae Walpers
Plant on stems or roots; cuticle wax crystalloids 0; leaves scale-like; "bracts" below flowers; flowers medium-sized to huge [to 120 cm across], (perfect); P/T 5/10/16-lobed, ± biseriate, (inner T/C connate, forming large dome [floral chamber, diaphragm], corona +, very small Raff.] / outer P only, true corona +, large - Sapria) / outer P only, corona 0 - Rhiz.), (nectary on distal part of P - Rhiz.); staminate flowers: A 12-40, (polysporangiate); microsporogenesis successive, pollen inaperturate, atectate; pistillode +; carpelate flowers: staminodes +; G ?number, carpel margins closed by postgenital fusion and secretion, loculi irregular, schizogenous, gynostemium short, stigma on outer margin or underside of disc; outer integument (reduced, 0), ca 1 cell across, inner integument ca 2 cells across, micropyle (exo-)endostomal, nucellar epidermis persists, nucellar cap +, obturator 0; antipodal cells ephemeral or not; fruit also splitting irregularly; seeds in two parts [constricted in the middle, the testa not fully enveloping the embryo], coat lignified; exotegmic cells cuboidal, wall thickening U-shaped; endosperm initially nuclear [Raff.], 30-40 cells, oily, embryo 6-22 cells, surrounded by a translucent membrane; n = 11, 12.
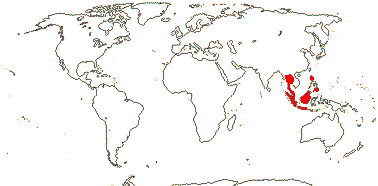
3/47: Rafflesia (42). S. China, Assam, Bhutan, Thailand, W. Malesia. Map: from Meijer (1997). [Photo - Flower.]
Age. The age of crown-group Rafflesioideae has been estimated to be (95.9-)81.7(-69.5) Ma (Bendiksby et al. 2010; see also Barkman et al. 2008; Pelser et al. 2019) or a mere (28.1-)15.1(-9.2) or ca 19.0 Ma (L. Cai et al. 2025b).
2. Apodanthoideae Walpers —— Synonymy: Apodanthaceae Takhtajan
Stem parasite; stomata anomocytic; flowers fairly small [7> mm across]; plant dioecious [Pi.]; "bracts" 2-3, an articulation, then 2-3 series of scales, secretory/nectariferous tissue in bract axils and abaxially/at the apices of the scales [Ap.], whorl of "bracts" [Pi], P +, 2-3(-4)-seriate, bi-, tri- or tetramerous [e.g. 2 + 4 + 4 or 3 + 6 + 6], (shortly clawed), (imbricate), inner whorl with adaxial tufts of hairs; staminate flowers: gynostemium +; A to 72 pollen sacs, locelli in 2 whorls [Ap.], no vascular bundles evident, vesicular hairs above A; pollen tricolpate, (apertures 0 - some Pi.), smooth; pistillode +/0, vesicular hairs on margin (all over); carpelate flowers: staminodes 0; G [4 (5)], (semi-inferior), opposite inner P, style at most short, very stout, hollow, chamber between style and stigma [Ap.], stigma ± hemispherical, stigmatic papillae peripheral, pollen tube transmitting tissue 0; ovules many/carpel, funicle with schizogenous cavity, long [Pi.], micropyle bi/endostomal, (endostome with secretory tissue - Ap.), or nucellus apex exposed, outer integument 1 cell across, inner integument 1-2 cells across; antipodals persist?; P persistent; dust seeds +, chalazal/funicular elaiosome + [radially elongated cells]; testa thin-walled, mucilaginous, tegmen tanniniferous, exotegmen massively lignified; endosperm +, cellular, ca 1-layered, embryo ca 8 cells [?all], globular; n = ± 12, 16, 30, chromosomes ca 1.4 µm long; plastome 11.3-15.2 kb, trnE [Pi], ndh genes 0.
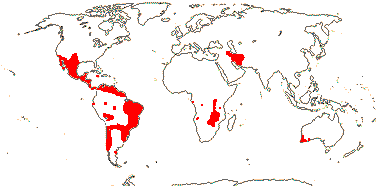
2 [list]/10: Pilostyles (9). New World from the S.W. U.S.A. southwards, S.W. Asia, S.W. Australia and E. (mostly) Africa. Map: from Fl. Austral. vol. 8 (1984), Novoa (2005), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 5 (2010) and Bellot and Renner (2014a). Photo: Flower.
Age. The age of crown-group Apodanthoideae was estimated to be (77-)57-31(-19) Ma while estimates for a stem age (in Cucurbitaceae) ranged from (98-)81-65(-44) Ma (Bellot & Renner 2014b); the crown-group age in L. Cai et al. (2025b) is (66.4-)53.1(-35.9) or ca 55.6 Ma.
Evolution: Divergence & Distribution. Rafflesiaceae are the oldest holoparasitic clade, and L. Cai et al. (2025b) suggested that they have a Gondwana-type distribution. Rafflesioideae in particular may have moved north on the Indian subcontinent - an out-of-India dispersal type - but with substantial diversification and migration since the mid-Miocene.
.If Sapria split off from other Rafflesioideae ca 81.7 Ma (Bendiksby et al. 2010), this raises the issue of when lowland tropical rainforest evolved. Today L.T.R.F. is the preferred habitat for Rafflesioideae in particular and for other parasites, as well as many echlorophyllous mycoheterotrophic taxa such as Burmanniaceae, Thismiaceae (Dioscoreales), Gentianaceae-Voyrieae (Gentianales), etc.. However, the dates in Naumann et al. (2013, q.v. for discussion), with the stem age of Rafflesioideae being around 65.3 Ma, suggest a rather different scenario. Bendiksby et al. (2010, q.v. for more dates; see also Barkman et al. 2008) thought that diversification within the subfamily did not begin until substantially after the origin of the clade, with a phylogenetic fuse of perhaps 80-90 m.y. from the ages in L. Cai et al. (2025b). Rhizanthes and Rafflesia may have separated ca 37 Ma (Naumann et al. 2013), while crown-group Rafflesia may be a mere (15.1-)11.8(-9.2) Ma, and this is the oldest of the crown-group ages suggested - perhaps there was an extinction event immediately prior to this. On the other hand the corresponding ages in Pelser et al. (2018) are ca 68 and ca 50 Ma respectively - all rather confusing. See also Cai (2023) fot the evolution of holoparasitism.
Thinking about the ages of the hosts of Rafflesioideae - all Vitaceae - allows one to look at the diversification of these parasites from another point of view. Fruits of crown-group Vitaceae from the Indian Deccan Traps have been dated to around or a little before the K/P boundary ca 66 Ma (Manchester et al. 2013), while H P. Chen et al. (2011b) suggest ages of (65.3-)50.6(-36.4) Ma for stem Tetrastigma, (49.3-)36.9(-25.7) Ma for the crown group, well before diversification of Rafflesia; ages in Lu et al. (2013) are somewhat older, at (67.7-)57.4(-47.4) and (59.4-)47.6(-36.4) Ma respectively, Peng et al. (2021) estimated that stem-group Tetrastigma was ca 49.4 Ma, while a mere ca 29.6 and 17.2 Ma are the stem and crown-group ages in Adams et al. (2016). If Rafflesioideae have always been obligate parasites of Tetrastigma, these ages again suggest interesting dating problems. Indeed, Cai et al. found that 17/41 of the horizontal genome transfers they examined were older that the others, and of these, 8 were to be found in Ampelopsis and 5 in the ancestor of Vitaceae. Given the likely long pre-diversification history of Rafflesioideae, one would very much like to know what their hosts were over this period. Indeed, although some of the mitochondrial genes that have moved into Rafflesioideae place them as sister to Vitaceae, other genes group with Cucurbitaceae and even Daucus (Apiaceae), suggesting that their hosts may have been rather different in the past (Z. Xi et al. 2013a).
The ca 79-fold increase in flower size during the evolution of stem-group Rafflesioideae over a period of ca 46 Ma may be linked to the adoption of sapromyophily; size increase in the subsequent ca 60 Ma was much more modest (Davis et al. 2007, 2008). However, in a more extensive study of Rafflesia, Barkman et al. (2008) suggested that there had been very considerable changes in flower size even within the last 12 Ma or so, the age of crown group Rafflesia, with repeated considerable increases and moderate decreases in flower size. It has been estimated that the ancestral flower size was (very approximately) 29 cm across (Barkman et al. 2008), the largest flowers are a little more than 1 m across (R. arnoldii) while those of R. consueloae are the smallest at around 10 cm across, although the perianth lobes in the latter are held more or less erect (Galindon et al. 2016). And now we have to factor in the evolution of Apodanthoideae, whose flowers are substantially smaller, being under 1 cm across...
All 12 (of the 13 species total) of Rafflesia from the Philippines studied formed a single clade, Borneo perhaps being their ancestral area (Pelser et al. 2018); inter-island dispersal was poor - Rafflesia is notably diverse in the Philippines. Indeed, most species of Rafflesia are found on but a single island in the Malesian archipelago and are critically endangered (Malabrigo et al. 2023).
Bellot & Renner (2014b) suggested that long distance dispersal was largely involved in allowing Apodanthoideae to attain their present distribution. On the other hand, Arias-Agudelo et al. (2019) thought that Apodanthoideae had become holoparasites before the break-up of the southern continents in the early Cretaceous (see also L. Cai et al. 2025b). However, although Arias-Agudelo et al. note that six genes were found in common in all three Pilostyles species whose chloroplasts were sequenced, suggesting genome reduction before their diversification, they also noted that plastome changes had occurred independently in those species (Arias-Agudelo et al. 2019).
There is a comprehensive and well-documented list of apomorphies for Rafflesiaceae s.l. in Alzate et al. (2024); this is in part the basis for the family characterization above, although there is still work to do in understanding seed coat anatomy, etc.. The two subfamilies have quite a lot in common, as was noted by Thorogood et al. (2021), although they thought that convergent evolution was involved. F. González and Pabón-Mora (2017b) observed that there were no recognizable meristems in Apodanthoideae other than the floral meristem, and A. D. González et al. (2020) found only 5/11 of the canonical gene families responsible for apical meristem maintenance in Pilostyles boyacensis. Furness and Rudall (2004) note a very distinctive combination of microsporogenesis and pollen morphology for Rafflesioideae; for pollen morphology, see also Blarer et al. (2004). The ovary loculi in Rafflesioideae develop by cell separation, i.e. they are schizogenous, unique in flowering plants, and the apex of the floral shoot becomes evident in a similar fashion (Nikolov et al. 2014a). Ng (2025) characterises floral development in Rafflesioideae as being closed, "all the organs in Rafflesiaceae [s. str.] are tightly packed from the start and can only have been formed in situ by internal cleavages in the protocorm" (ibid., p. 137).
Ecology & Physiology. All told, there are 5 genera and 57 species in this clade of holoparasites, and they have been seen growing primarily on Vitaceae and Fabaceae (Hatt et al. 2024c). Thus recorded hosts of Apodanthoideae include almost 40 genera of Fabaceae-Cercidoideae, -Caesalpinioideae, -Detarioideae and -Faboideae for Pilostyles alone (Arias-Agudelo et al. 2019) and Salicaceae-Samydoideae-Casearia for Apodanthes (Bellot & Renner 2014a). Indeed, the distribution of Apodanthoideae given above is probably an underestimate, since in Africa Pilostyles is parasitic on e.g. the widespread Brachystegia and Julbernardia in the Miombo woodland (White 1983), and the plant has been mistaken for a rust fungus... On the other hand Rafflesioideae are parasitic on species of Tetrastigma (Vitaceae) alone, although this association may have evolved more than once (P. Chen et al. 2011a; see also Divergence & Distribution above). In a quite extensive study, all the hosts of the eleven species of Rafflesia in the Philippines examined were found to belong to Tetrastigma, and one or more species of Rafflesia were found on six of the eight species of that genus growing there, but there was not much host specificity (Pelser et al. 2016), earlier, Nais (2001) had suggested greater specificity (only two species were known from the Philippines in 2002).
The vegetative plant body of Rafflesiaceae is endophytic. Wurdack and Davis (2009) early suggested that the plant body of Rafflesioideae might be derived from laticiferous tissue, e.g. as found in Euphorbiaceae. In Rafflesia and Sapria, at least, two or more individuals can grow on a single plant of Tetrastigma, and the different individuals on the one host tend to be closely related to each other (Barkman et al. 2017), and the are both staminate- and carpelate-dominant populations (Wee et al. 2025). The endophyte is quite inconspicuous, almost filamentous, and the cells are undifferentiated (Nikolov et al. 2014b); Mursidawati et al. (2019) suggested that the endophyte did not form continuous strands, rather, there were cell clusters, the cells being squeezed between dividing fusiform initials, separating and and spreading in the host. The parasite obtains all its nutrients from the host. Wicaksono et al. (2021a) thought that the cells somehow maintained contact; there may be up to 13 individuals of Rafflesia on the one host, and those of R. tuan-mudae extended up to 14.7 m along the vine (Barkman et al. 2013). When a vascular strand of the parasite finally developed, it was made up of vessel elements and parenchyma (Wicaksono et al. 2021a). A vascular connection between host and parasite was established only late in development, i.e. at the time of flowering (Teixeira-Costa et al. 2021 - see also Apodanthoideae), indeed, in both subfamilies this vascular connection is established late (Teixeira-Costa et al. 2021). Rafflesia can spread 14 m or more within its host (see also Barkman et al. 2017), and Wee et al. (2025) suggest that generation tome in R. cantleyi, at least, is 3-4.5 years, while in other taxa it is perhaps as much as 5.3 years. Note that Ng (e.g. 2025) has interpreted development in Rafflesioideae rather differently, suggesting that the various floral structures separate from one another in the protocorm, an initially featureless swelling on the Tetrastigma host. However, a clearer understanding of the endophyte in Rafflesioideae depends on finding out more about the germination and establishment of the parasite, although Ng (2025) suggests that germination does not occur here, but somehow or other “their seeds get in to their Tetrastigma hosts", strands or clusters of proembryonic cells being released, protocorms developing, and so on.
Rutherford (1970, and references) and others have looked at the anatomy of Pilostyles thurberi. For the effect of Pilostyles on the wood structure of its host, Mimosa, see do Amaral and Ceccantini (2011).Pollination & Seed Dispersal. Rafflesia may be dioecious or monoecious; agamospermy occurs here (Barkman et al. 2017; Pelser et al. 2017; Twyford 2017; Wicaksono et al. 2021b). The flowers of at least some Rafflesioideae are thermogenic (Seymour 2001; see elsewhere for how thermogenic flowers may function) and pollination, where known, is by flies - oviposition-site mimicry occurs throughout the family (S. D. Johnson & Schiestl 2016). The flowers look and smell rather like a rotting carcass and the pollen is presented in the form of a slurry, a syndrome that occurs in other fly-pollinated flowers (Bänziger 2004; Davis et al. 2008; Nikolov et al. 2014a; see also Jürgens et al. 2013 for this syndrome), however, Ng (2024) did not notice any smell from a Rafflesia flower that he happened to have in his car. Wee et al. (2018) found that females of five species of calliphorid flies were preferentially attracted to Rafflesia cantleyi, only one species actually being a pollinator; dimethyl di- and trisulphides were the main components of the smell.
Ortega-González (et al. 2020) described Pilostyles maya as having cleistogamous and hermaphroditic flowers, M. F. Romero et al. (2025) suggests that P. berteroi may be dioecious, F. González and Pabón-Mora (2017b) found that flowers of P. boyacensis were sometimes male-female chimaeras; Ortega-González et al. (2022) was unsure if P. thurberi was monoecious or dioecious. There may be chalazogamy: "the space [in the funicle of Pilostyles] facilitates the occurrence of chalazogamy" (F. González & Pabón-Mora 2017b: p. 534). Clearly, there is work to do here!
Apodanthoideae seem to be pollinated by a variety of insects - ants, bees, beetles, bugs, butterflies, flies, thrips and wasps (Bellot & Renner 2013; Ortega-González et al. 2022 and references). A variety of insects visit the flowers of Apodanthes, but which of them are the actual pollinators is unclear - the flowers may even be ant-pollinated (Ramírez-Ramírez et al. 2025). The flowers of Pilostyles are visited by blowflies, bees and wasps. The pollen grains of P. berteroi germinate amongst the papillae on the lateral part of the stigma (Romero et al. 2025).
Little is known about seed dispersal and germination in the family as a whole. The minute seeds - up to a quarter of a million per fruit (Cai et al. 2021) - of Rafflesia are embedded in ?placental tissue and the fruits may be eaten by rats (Bänziger 2004), while squirrels and tree shrews were seen eating the oily, custard-like contents of the fruit of the Bornean R. keithii (Emmons et al. 1991). There may be both ecto- and endozoochory (Wicaksono et al. 2021b). During germination of some Rafflesia, at least, the seed is anchored to the host by sticky endosperm tubules and also by the embryonal primary haustorium (Arekal & Shivamurthy 1976), the whole thing looking rather like a T4 bacteriophage. Wicaksono et al. (2021a) suggest that the seed, which they draw as being faintly like a seahorse, germinated by its micropylar tip, i.e. by the tail of the horse (see also Ng 2024).
The fruits of Apodanthoideae are dispersed by animals (Bellot & Renner 2013), around 14-400 or so seeds per fruit being produced by Pilostyles thurberi (Ortega-González et al. 2022). Crematogaster carinata ants, which live on the host plant of A. caseariae, Casearia sylvestris, eat the secretions produced by the fruits and bracts of Apodanthes; myrmecochory is likely here (Ramírez-Ramírez et al. 2025).
Genes & Genomes. Sapria himalayana (Rafflesioideae) has lost some 44% of its eurosid genes, over double the loss in Cuscuta, and many of these genes are functionally rather similar to those lost in other parasitic plants, however, the ability to synthesize abscisic acid seems also to have been lost here (Cai et al. 2021). Despite the loss of so many genes, the overall genome size is quite large. However, a number of housekeeping genes are very small, having fewer introns even than in the carnivorous Genislea, while at the same time introns in other genes are far larger, over a quarter being over 1 kb long and the longest ca 97.8 kb long (Cai et al. 2021). X. Chen et al. (2023) compared the genome of Sapria with that of Balanophora (Balanophoraceae), and in some respects they were quite similar - parallelism (see also Molina et al. 2023 and such parallelisms in parasitic plants). Molina et al. (2023) looked at the transcriptome of the seed of Rafflesia speciosa comparing it with that of some other parasites, etc...
Xi et al. (2012a, see also Kado & Innan 2018) showed that slightly over 2% of the nuclear genome in the host Tetrastigma rafflesiae may have moved to the parasite, Rafflesia cantleyi, some Tetrastigma genes apparently functioning in their new host, and even in a number of vertically transmitted Rafflesia genes, codon usage was Tetrastigma-like, perhaps facilitating the intimate association of the host and parasite (see also Molina et al. 2023 for genes involved in horizontal transfer that may effectively camouflaging the parasite). Cai et al. (2021) teased apart this movement of genes from Vitaceae to Rafflesioideae in some detail. They noted that while genes from Tetrastigma and the ancestor of the [Tetrastigma + Cayratia] clade have moved widely into members of Rafflesioideae, a number of genes from Ampelopsis and other genes pegged to the common ancestor of Vitaceae are associated with stem Rafflesioideae (Cai et al. 2021: see also Divergence & Distribution above). Of course, at least some movement of nuclear genes from host to parasite is not uncommon, especially when the latter is holoparasitic (e.g. Y. Zhang et al. 2013; Z. Yang et al. 2014, 2016; Kado & Innan 2018; Sanchez-Puerta et al. 2023); see also Orobanchaceae and Cuscuta (Convolvulaceae)).
For the apparent absence of the plastid trnE gene from the whole family, see Ceriotti et al. (2025: ?Apodanthes). Molina et al. (2014) thought that the plastome in Rafflesia lagascae had been very largely lost (see e.g. Bellot & Renner 2015; Naumann et al. 2016; S.-M. Ng et al. 2018 for discussion). Those gene fragments that could be detected lacked open reading frames, they were not expressed in floral bud tissues, and a third of them had sequence similarities with Tetrastigma genes; any chloroplast remnants might be in the nucleus (see also Krause 2015). There is also evidence for the loss of the plastome in Sapria (Cai et al. 2021); note that although plastomes of parasites may be lost, substantial numbers of plastid genes may move from the host via mitochondria to the parasite (Xi et al. 2013a; Molina et al. 2014; Davis & Xi 2015). Ng et al. (2018) suggested that in R. cantleyi some essential plastid genes had moved to and were encoded by the nucleus, but that synthesis of things like haeme might still go in the plastid.
Bellot and Renner (2015) examined the plastomes of two species of Pilostyles (from different continents) and found only five to six functional genes remaining, and the minute but probably circular plastomes lacked an inverted repeat. This plastid genome is about the smallest known (Barrett & Kennedy 2018: Rafflesioideae not included; Arias-Agudelo et al. 2019), and all tRNAs needed for plastid protein synthesis must be imported (Bellot & Renner 2015: see also Balanophora). There were numerous plastome regions in both the nucleus and particularly the mitochondrion (c.f. mitochondrial genome of Cucurbitaceae), although some of these may have come from the host (Bellot & Renner 2015). Arias-Agudelo et al. (2019) suggest that some parasite chloroplast genes may have moved via horizontal transfer from their leguminous hosts, but what was going on with other genes that were showing initial similarities with a variety of non-leguminous taxa was unclear. For gene loss, especially ndh genes, see also Lin et al. (2017) and Mower et al. (2021: possible connections between various distinctive life styles that might affect the photosynthetic process).
C. C. Davis and Wurdack (2004) noted that the sequence of the chondrome nad1B-C gene in Rafflesioideae strongly suggested a relationship with Vitaceae; they reasonably thought that the presence of this gene in Rafflesioideae was the result of horizontal gene transfer from Vitaceae. Barkman et al. (2007) suggested that there had also been a transfer of the mitochondrial atp1 gene from host to parasite. Xi et al. (2013a) confirmed major gene movement in the chondrome, 24-41% of the genes examined having moved from host to parasite, probably by homologous recombination, and again these genes seemed to be functional in their new environment (see also Davis & Xi 2015). This is the closest integration of host and parasite genome so far known in land plants. For the much increased rate of variation in synonymous substitution in some mitochondrial genes of Apodanthoideae, see Mower et al. (2007 and references). Barkman et al. (2007) found that mitochondrial genes (atp1) from host Fabaceae had moved to Pilostyles thurberi.
Chemistry, Morphology, etc.. Rafflesioideae are tanniniferous (Gottlieb et al. 1989). Although there are stomata in Rafflesia, they are clearly abnormal, having three or more guard cells (Cammerloher 1920; see also Wicaksono et al. 2021b).
And flowers... There has been much debate as to what the perianth and the diaphragm/annulus (the latter forming the floral chamber) of the flowers of Rafflesioideae might represent, although this has largely been cleared up by Nikolov et al. (2013, 2014a). Sapria can be interpreted as having a biseriate perianth (the spreading lobes of the flower), while the diaphragm in the middle is a corona of sorts (perhaps similar to such structures in Passifloraceae); it arises where the expression of B and C genes changes (Nikolov et al. 2013: morphological and gene-expression studies). However, in Rafflesia the tubular structure below the diaphragm seems to have an inner and outer portion, suggesting that the spreading lobes represent the outer portion and are the calyx/outer perianth whorl and the diaphragm is the inner portion, equivalent to the connate corolla/inner perianth whorl (also D. Boufford, pers. comm.), while a rim at the base of the tube/around the gynostemium is the annulus. Rhizanthes lacks a diaphragm, but its perianth tube clearly has an inner and outer portion, and a slight bump on the perianth may be is where a fragmented annulus has become adnate to the perianth. Thus Nikolov et al. (2013) showed that the tubular structures found in these three genera developed in different ways - in Rafflesia it is a K/C tube alone while in Sapria and Rhizanthes there is a K/C tube in large part formed by the activities of the annulus (Nikolov & Davies 2017).
The outer integument of the ovule of Rafflesioideae, when present, is one cell layer thick, but it is not easy to interpret the ovule (see Solms-Laubach 1874; Ernst & Schmid 1913 for more details). Although the funicle is bent, the integument is not adnate to it; in taxa with a single integument, there is a swelling on the chalaza, perhaps an indication of the other integument.
Turning to Apodanthoideae, there are cushions of hairs at the bases of the inner perianth parts. Interpreting the merosity of the flower and the nature of the perianth parts is not easy, and the androecium in particular is difficult to understand (Blarer et al. 2004; F. González & Pabón-Mora 2017b). Pilostyles, at least, has a distinctive pedicel with schizogenous intercellular spaces. Ramírez-Ramírez et al. (2025) noted that the seeds of Apodanthes caseariae were exotestal.
For additional information, see see Harms (1935a), the Parasitic Plants website (Nickrent 1998 onwards), Heide-Jørgensen (2008) and the references in Alzate et al. (2024), all general; for Rafflesioideae in particular, Meijer (1993), Nais (2001: superb photographs), Ng (2024: Rafflesia) Takhtajan et al. (1985: pollen), Bouman and Meijer (1986: seeds, 1994: ovules and seeds) and Baskin and Baskin (2021: seeds, etc.). For Apodanthoideae, see Kuijt (1969), Nickrent (2020), also Takhtajan et al. (1985: pollen), Visser (1981), Blarer et al. (2004: floral morphology) and Baskin and Baskin (2021: seeds, etc.). For vesicular cells, see Blarer et al. (2002, 2004), for a detailed study of Pilostyles ingae, see Endriss (1902) and for the embryology of Pilostyles, see Rutherford (1970).
Phylogeny. Relationships within Rafflesioideae are [Sapria [Rhizanthes + Rafflesia]] (Davis et al. 2007; Pelser et al. 2018; see also Nickrent 2020). For relationships within Apodanthoideae, see Filipowicz and Renner (2010) and Arias-Agudelo et al. (2019); the Australian species of Pilostyles are sister to those from the New World.
Classification. Having two subfamilies seems reasonable - c.f. their positions in the old Rafflesiales (Nickrent et al. 2004a).
Previous Relationships. Rafflesiales of some authors included a number of other echlorophyllous, parasitic groups such as Apodanthaceae (see above), Cytinaceae (here Malvales), Hydnoraceae (Piperales) and Mitrastemonaceae (Ericales). Many authors have sought an affinity between Rafflesioideae and taxa like Aristolochiaceae (references in Takhtajan 1997), perhaps in part because of a belief that the pollen of the former had only a single aperture, as did that of Aristolochiaceae; there is a gynostemium of sorts and extrorse anthers in both. Cocucci and Cocucci (1996) saw connections of Rafflesiaceae first with Apodanthaceae and then with Annonaceae. However, Ng (2024) thought that the vegetative and floral growth of Rafflesiaceae was so bizarre that the plant was not an angiosperm, nor even a phanerogam - although he said at the same time that it had a full complement of recognizable plant structures; Ng (2025) floated the idea of a novel group, the rafflesiosperms.
Thanks. To Lachezar Nikolov, for helpful discussions on floral morphology of Rafflesiaceae, and to S. Renner for comments on Apodanthoideae.
[[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]]] [Euphorbiaceae [[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]]] / Clade 1 of Xi et al. (2012b): ?
Age. This node is estimated to be ca 82 Ma (T. Xue et al. 2020: only Salicaceae and Euphorbiaceae included), (111.8-)107.9(-104.5) Ma (Xi et al. 2012b: Table S7) or (103.6-)83.4(-59.6) Ma (K. Yan et al. 2025).
Phylogeny. For discussion of relationships in this clade, see below.
[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]]]: endosperm persistent.
Age. This node is around (110-)105.7(-101.6) Ma (Xi et al. 2012b: table S7) or some 105.8 Ma (Tank et al. 2015: table S2).
HUMIRIACEAE A. Jussieu, nom. cons. - Back to Malpighiales
Trees; ellagic acid +; cork subepidermal; vessel elements solitary, with scalariform perforation plates; true tracheids +; vestured pits +; sieve tube plastids with protein crystals and starch; secretory glands/canals +; nodes 5:5; petiole bundle annular, with wing bundles; rhombic calcium oxalate crystals +; secretory ducts +, mucilage cells frequent; stomata anomocytic; branching from previous flush; leaves often two-ranked, lamina vernation involute, venation bronchidodromous, glands/extrafloral nectaries +, teeth glandular-deciduous, ?type, (margins entire), petiole short, stipules small or 0; inflorescence cymose; pedicels articulated; K connate, at least at base, quincuncial, C (quincuncial/cochlear); A 5 + 5 staminodes opposite K or 10 (obdiplostemonous)-30 or (A 5 X 3 opposite K, filaments connate + 10 or more, filaments ± connate at least basally, with interdigitated hairs higher up, forming a tube, connective broad, prolonged; pollen exine usu. microreticulate; nectary prominent, annular; G [(4-7)], opposite K, (carpels with 5 traces), style +, unbranched, stigma capitate, ± 5-lobed/-radiate, ?type; ovules epitropous, micropyle exo(endo)stomal, outer integument 2-3 cells across, inner integument 2-3 cells across, parietal tissue 3-6 cells across, nucellar cap +, endothelium 0; fruit a drupe, stone operculate, 1-3(-5)-seeded, surface sculpted, with "resin" cavities; exotestal cells thick-walled, lignified, tegmen multiplicative [ca 5 cells across], cross layer of fibres beneath exotegmen; endosperm copious [?always], perisperm +, slight, embryo somewhat curved, green; n = 6 [Sacoglottis gabonensis].
7 [list]/63. Tropical America, W. Africa. Map: from Thorne (1973) and Trop. Afr. Fl. Pl. Ecol. Distr. Vol. 1 (2003); see also Prance (2021). Photo: Flower, Fruit.
Age. Crown-group Humiriaceae are estimated to be (32.1-)20.7(-10.4) Ma (Xi et al. 2012b: table S7).
1. Vantanea Aublet
Stomata paracytic; C with 3 traces; A ≥50 in 3 or more whorls, anther thecae 2, bisporangiate, sporangia dehiscing by slit; ovules 2/carpel, superposed.
1/22. Tropical America.
[Humiria + The Rest]]: stomata anomocytic; A 10-30, sporangia 2, separate, valvate.
2. Humiria Aublet
G opposite C; ovules 2/carpel, superposed.
1/5. Tropical South America.
(Sporangia 4, in superposed pairs); ovule 1/carpel.
6/36: Humiriastrum (16), Sacoglottis (11). Tropical America, W. Africa (Sacoglottis gabonensis only)
Evolution: Divergence & Distribution. Herrera et al. (2010, see also 2014a) rejected all fossils placed in this family other than some from South America, and they suggested that Humiriaceae originated there.
Ecology & Physiology. In Panama species of Humiriaceae grew in soils with low phosphorus (Condit et al. 2013).
Seed Dispersal. The fruits are dispersed by bats or by water, empty cavities in the stone affording bouyancy.
Chemistry, Morphology, etc.. Costa et al. (2021) describe secretory canals in Humiriaceae as forming by the coalescence of cavities of various shapes. Wurdack and Zartman (2019) surveyed the family for foliar extrafloral nectaries; they also reported nectariferous connective glands for the family (see also de Almeida et al. 2023b) - nectar from these would be in addition to that from the annular nectary at the base of the ovary.
Although D. A. Link is sometimes cited as the author of a paper on the nectaries of Humiriaceae, and he promised such a paper himself, it seems never to have appeared. The androecium is very variable. Species of Schistostemon have 20 stamens, each of those opposite the sepals having three anthers (= 3 stamens fused), and Sacoglottis itself is similar, but with only 15 stamens (Herrera et al. 2010; esp. Wurdack & Zartman 2019). Prance (2021) described the seeds as having a double testa, the outer testa being more or less adherent to the endocarp.
Some information is taken from Herrera et al. (2010) and Kubitzki (2013b), both general, and Prance (2021: monograph), also Herrera et al. (2014a: summary of what is known about wood anatomy), Narayana and Rao (1977 and references: floral morphology), Bove and Melhem (2000: pollen), Mauritzon (1934d: ovules), and Boesewinkel (1985a: ovule and seed).
Phylogeny. Morphological phylogenetic analyses initially suggested that Vantanea was sister to the other Humiriaceae; it has many stamens in three or more whorls (Bove 1997). Herrera et al. (2010: detailed morphological analysis of 40 characters) suggested that Vantanea and Humiria were successively sister to the remainder of the family, although support for this topology was weak; the [Humiria + the rest] clade had quincuncial corollas and 30 or fewer stamens with unilocular anthers, the stones often had "resin" cavities, etc.. However, Schistostemon was sister to the other three genera included (including Vantanea) in a molecular analysis, but support was weak (Xi et al. 2012b); M. Sun et al. (2016) found that Schistostemon was embeddded in Sacoglottis, sister to the rest of the family examined, but the interrelationships of the latter were unclear. Prance (2021) noted that relationships at the base of the family were generally [Vantanea [Humiria ...]], but that the topology recovered by Chave (no further details given) was [Humiria [Vantanea ...]].
Previous Relationships. Bove (1997) suggested that Ixonanthaceae were sister to Humiriaceae, both having ellagic acid, a "free" annular nectary encircling the ovary, and an undivided style with a more or less entire stigma. Humiriaceae have also been linked with Linaceae and Erythroxylaceae, and thence to Geraniales (Narayana & Rao 1978b), or the three families together are placed in Linales (Cronquist 1981).
[Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]] / Parietal Clade: at least some vessel elements with simple perforation plates, crystals in ray cells; sieve tubes with non-dispersive protein bodies; cuticle waxes usu. 0; (foliar glands +); pedicels articulated; nectary outside A, annular; G with median member abaxial; placentation intrusive parietal, ovules several/carpel, nucellus massive; seeds arillate; endotegmen persistent; endosperm oily.
Age. This node has been dated to (72-)69, 65(-62) Ma (Wikström et al. 2001), ca 99.2 Ma (Tank et al. 2015: table S2), (114-)108(-104) Ma (Davis et al. 2005a: note topology), (89-)81, 79(-74) Ma (Bell et al. 2011: Ixonanthes also included!), (104.6-)99.2(-93.1) Ma (Xi et al. 2012b: Table S7), ca 129.4 Ma (Ogutcen et al. 2024) and ca 107.9 Ma (L. Cai et al. 2025b).
Evolution: Divergence & Distribution. All the families in this group and alse the three major subfamilies of Passifloraceae may have diverged in the Cretaceous-Albian 111-100 Ma, or a little later (Davis et al. 2005a: q.v. for details). At the other extreme, Wikström et al. (2001) suggested that many of the clades did not diverge until (well) after 63 Ma. Elioxylon, from Intertrappean Indian deposits around 65 Ma, has been identified as Achariaceae or Salicaceae (Srivastava et al. 2018). The authors emphasize its pentagonal pith, which, they note, is also known from Lindackeria and Populus and, one might add, Goupia (see below); that of Eliosylon is particularly distinctive in having peripheral, palisade-like cells (Srivastava et al. 2018). Interestingly, given the relationships in Wurdack and Davis (2008), the distinctive cyclopentenoid glycosides scattered in this clade may have evolved more than once and/or been lost.
Plant-Animal Interactions. Larvae of butterflies such as Nymphalidae-Acraeinae and N.-Nymphalinae-Heliconiini, -Vagrantini and -Argynnini commonly eat members of this group (Ehrlich & Raven 1964; see also Arbo 2006; Simonsen 2006; Silva-Brandão et al. 2008; Nylin & Wahlberg 2008; etc.); this is also discussed under the individual families below. Thus Turneroideae are the hosts of caterpillars of several genera of Nymphalidae, alternate hosts include Salicaceae, Passifloroideae, and Violaceae (Arbo 2006 and references). Some Acraeinae in particular may cue in on the presence of the cyanogenic glucoside gynocardin in potential food plants, indeed, that larvae of Acraea horta, which normally ate leaves of the gynocardin-containing woody Kiggelaria africana, also ate herbaceous Achariaceae s. str., prompted the successful search for that compound in the latter (Steyn et al. 2002). Toxic compounds like gynocardin may be sequestered by the larva and passed on to the adult.
Phylogeny. For relationships in this clade (= the parietal clade), which has strong support, see Xi et al. (2012b).
Chemistry, Morphology, etc.. Spencer and Seigler (1984, 1985b) and Spencer et al. (1984) and references discuss cyclopentenoid cyanogenic glycosides and Nielsen et al. (2017) cyanogenic glycosides in general. There is much information on seed anatomy in Takhtajan (1992) while Krosnick et al. (2006) briefly discuss the evolution of polyandry in this group - in some cases, at least, the numerous stamens form a single whorl. See Mauritzon (1936b) for some information on embryology and Furness (2011) for pollen development and ultrastructure.
Information about Achariaceae, Lacistemataceae and Salicaceae in much of the older literature is to be found under Flacourtiaceae (see next paragraph).
Previous Relationships. It was commonly agreed that the old Flacourtiaceae presented major taxonomic problems. "Flacourtiaceae as a family is only a fiction; only the tribes are homogeneous" (Hermann Sleumer, a monographer of the family, in R. B. Miller 1975: p. 79) - and it was indeed a fiction. Some of the old Flacourtiaceae are now in Achariaceae, a few in Lacistemataceae, while Flacourtiaceae-Berberidopsideae are in Berberidopsidales-Berberidopsidaceae, Aphloia is in Crossosomatales-Aphloiaceae and Gerrardina in Huerteales-Gerrardinaceae. Variation in chemistry, leaf teeth, floral morphology, seed coat anatomy, etc., is largely correlated with the new family limits.
When parietal placentation was considered to be a very important characters, other families with parietal placentation such as Caricaceae (Brassicales), Cucurbitaceae (Cucurbitales), etc., might also be placed here, and the combined group was appropriately called Parietales.
ACHARIACEAE Harms, nom. cons. - Back to Malpighiales
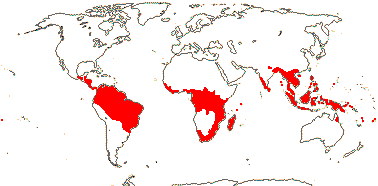
Cyclopentenone cyanohydrin glycosides and/or cyclopentenyl fatty acids [gynocardin; tetraphyllin rare]; fibres septate; axial parenchyma usu. 0; ray cells with scalariform perforations [?distribution]; petiole bundle annular, with two wing/adaxial strands; leaves spiral or two-ranked, lamina margins entire (serrate), (stipules 0), petiole often geniculate; inflorescence spicate or cymose (fasciculate); K and C (not distict), (not in a simple alternating relationship) K 2-5, C 4-15, often in two series, (adaxial basal scales +); disc 0; A usu. many, opposite C or irregularly inserted, initiation centripetal or simultaneous, (from a ring meristem), anthers basifixed, elongate, (dehiscing by pores), (locellate); pollen also tricolporoidate; G [2-10], style (short), branched or not, stigma capitate-peltate to punctate; ovule number various, (1/carpel), ± sessile and broadly attached to placenta, (straight - Xylotheca, Hydnocarpus, Lindackeria, Scaphocalyx), micropyle endo- or bistomal or zigzag, outer integument 3-7 cell layers across, (lobed), inner integument 3-8 cell layers across, parietal tissue 4+ cells across, nucellar cap +, epistase +, ring/cap of tracheids in chalaza; (megaspore mother cells 2), embryo sac penetrating chalaza, forming caecum below tracheids; fruit also a berry; (seed arillate); seed coat thick, vascularized [pachychalazal], testa multiplicative, vascularized or not, endotesta lignified, cells sclereidal (radially elongated), tegmen multiplicative, exotegmic cells elongated, massive, sclereidal; endosperm copious, suspensor 0, embryo chlorophyllous; n = 10, 12, 23.
33 [list: 5 tribes]/180. Pantropical. Map: from Sleumer (1954, 1980), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003), Fl. China vol. 13 (2007), Khan et al. (2014), Serban Prochš (pers. comm. - Africa) and Andrew Ford (pers. comm. - Australia). [Photos - Flower, Fruit, Fruit, Acharia tragodes - Leaves.]
Age. This node has been dated at (71-)59, 58(-44) Ma (Bell et al. 2011: Ixonanthes included!), (60-)57, 54(-51) Ma (Wikström et al. 2001), and (86.2-)65(-48.2) Ma (Xi et al. 2012b: Table S7, Hyd. Pang. Ach.).
Acharia Bentham & J. D. Hooker, Ceratosicyos, Guthriea. 3/3: South Africa.
More or less herbaceous left-twining vines (subshrubs, herbs); leaves palmately lobed (not G.), (stipules 0); inflorescence monochasial-cymose/flowers axillary; flowers imperfect, 3-5-merous; K quincuncial/(open?), C postgenitally connate; vascularized antepetalous glands +; staminate flowers: A 3-5, opposite K, adnate to C [G.], anthers with swollen hairs, thecae (separate), broadly attached to filaments, endothecium 0; pistillode 0/+; carpelate flowers: staminodes 0; G [5], style short, stigma lobed; ovules apotropous, micropyle endostomal/bistomal-zig-zag, outer integument 4-7 cells across, inner integument 3-6 cells across, parietal tissue 6-15 cells across, nucellar cap ca 3 cells across, apex of nucellus pushing into micropyle [check]/not, suprachalazal tissue massive, funicle (well developed); embryo sac bisporic, [chalazal dyad], eight-celled [Allium-type]); fruit a capsule; seeds with pits or tubercules, weakly ruminate or not, chalazal area elaborated, cap-like [= "imbibition lid"], raphe ridged (not - C.); sarcotesta (pubescent - G.), stomata +, testa not vascularized, exotegmen fibrous; embryo short; n = ca 10.
1. Acharieae Bentham & J. D. Hooker —— Synonymy: Kiggelariaceae Link, nom. inval., Pangiaceae Hasskarl
Perennial herbs to trees, climbers; ellagic acid + [Kiggelaria]; vessel elements perforations simple (+ scalariform); plant usu. monoecious or dioecious; K and C distinct, Kcalyptrate, or 3-5, valvate or open, C 3-5(-9), (connate), adaxial basal scales +; A 3-5(-many), (anthers barely elongate - Chiangiodendron); (G with nectariferous hairs - Scaphocalyx [S.]); (ovules immersed in G wall - S.); fruit berry or capsule, seeds 1-many; (testa with crossing layers in middle - Pangium), (mesotesta with sclerenchyma, palisade layer, and sclereids - S.).
12/32: Ryparosa (25). Old World: Sri Lanka, Malesia, Australia (Queensland), Assam, and also Mexico (Chiangiodendron).
Age. This node has been dated to around (33-)23, 22(-12) Ma (Bell et al. 2011).
[Hydnocarpeae [Phyllobotryeae [Erythrospermeae + Lindackerieae]]]: ?
2. Hydnocarpeae Zmarzty & P. C. Bailey
Trees to shrubs; vessel elements with scalariform perforation plates; plant usually dioecious; K and C distinct, usu. cyclic, free, K (3-)4-5(-11), imbricate, C 4-5(-16), basal adaxial scales +; A 5-many; fruit indehiscent, seeds many.
2/46: Hydnocarpus (45). Sri Lanka and India to Malesia.
Age. Intertrappean fossil wood of Hydnocarpoxylon, early Palaeogene in age (ca 65 Ma), definitely has features of Hydnocarpus (Wheeler et al. 2017; Srivastava et al. 2018).
[Phyllobotryeae [Erythrospermeae + Lindackerieae]]: ?
3. Phyllobotryeae Warburg
Single-stemmed shrub; vessel elements?; inflorescence epiphyllous; K and C distinct, cyclic, 3-5, imbricate, C adaxial scales 0; A 5-many; fruit capsular, seeds few.
2/7: Phyllobotrya (5). West and Central Africa.
[Erythrospermeae + Lindackerieae]: ?
4. Erythrospermeae de Candolle —— Synonymy: Erythrospermaceae Doweld
Trees to shrubs; vessel elements with scalariform perforation plates; P spiral, K and C intergrade, 9-14 or K (calyptrate - Prockiopsis) 3-6, imbricate, C 4-11, adaxial scale +/0; A usu. many; fruit usu. a capsule, seeds 1-few.
5/34: Erythrospermum (12). Africa, Madagascar, Erythrospermum Mauritius to Fiji.
5. Lindackerieae Zmarzty
Shrubs to trees; vessel element perforations various; (stomata anomocytic - Carpotroche); K and C distinct, K (2) 3 (4), imbricate (valvate), C 4-25, adaxial scale 0; A many; fruit various, seeds 1-many.
12/51: Lindackeria (14), Carpotroche (12), Caloncoba (10). Central and South America.
Evolution: Divergence & Distribution. Of the some 33 genera of Achariaceae, 14 are monotypic and 12 more include 2-5 species, furthermore, the family has little in the way of distinctive characters, but there is extensive variation at the tribal and generic levels (Zmarzty et al. 2025).
Pollination Biology & Seed Dispersal. Guthriea (Acharieae), from high altitudes in the Drakensberg, is pollinated by lizards, both the floral scent and the bitterness of the nectar perhaps coming from metabolites involved in the saffron pathway (Cozien et al. 2019).
Acharia and Guthriea may be myrmecochorous, the ants being attracted by the sarcotestal-chalazal region; the seeds of Kiggelaria are dispersed by birds (Steyn et al. 2002a).
Plant-Animal Interactions. The feeding behaviour of Acraeini butterfly larvae are consistent with the family limits adopted here, for instance, caterpillars of Acraea horta may be found on both Kiggelaria and Guthriea (van Wyk in Dahlgren & van Wyk 1988; Steyn et al. 2002a, 2003 and references). Species of Ryparosa consistently produce glycogen-containing food bodies, and in a number of species there are associations of varying closeness with ants (Webber et al. 2007).
Chemistry, Morphology, etc.. There are large and medium intervascular pits; the wood also has solitary pores and lacks tracheids (R. B. Miller 1975). Lindackeria has superficial cork cambium.
Pollen variation is considerable (Wendt 1988; see also Gavrilova 1998), as is that in ovule development and seed coat anatomy (Dathan & Singh 1979; van Heel 1973, 1974, 1977a, 1979; Steyn et al. 2001, 2002a, b, 2003).
For general information, see Lemke (1988), Judd (1997a), Van Wyk in Dahlgren and Van Wyk (1988: Acharieae), Chase et al. (2002) and Groppo et al. (2010), also Hegnauer (1966, 1989: chemistry), Thadeo et al. (2014: some lamina anatomy); Endress and Voser (1975: floral development, Caloncoba), Bernhard (1999b: ditto, Ceratiosicyos), and Bernhard and Endress (1999: androecial initiation). Information in the older literature is to be found under Flacourtiaceae.
Phylogeny. For the circumscription of Achariaceae, see Chase et al. (2002) and Sosa et al. (2003). Chase et al. (2002) found that the family could be divided into three quite well-supported clades, [1-Hydnocarpus [2-Erythrospermeae + Lindackerieae]] [3-Pangieae, Acharieae]], and support for the monophyly of the family as a whole is strong. However, Sosa et al. (2003) did not find much support for the last clade. Groppo et al. (2010) also questioned some tribal limits in the family, and the relationships suggested by M. Sun et al. (2016) and Pagart (2017) should also be consulted; i.a. the latter noted that Hydnocarpus tended to be peripatetic and that a number of support values were low. Zmarzty et al. (2025: all 33 genera and a number of possible relatives included) looked at the phylogeny here using the Angiosperms353 probe set and found relationships that in part agreed with those apparent in earlier work, but tribes like Hydnocarpeae were unexpected. Support for the basal branches in the tree were quite strong, but some branches in Lindackerieae were only weakly supported, Lindackeria is polyphyletic, andCaloncoba, Camptostylus and Xylotheca may best be combined - Caloncoba is polyphyletic, the two other genera being embedded in it (Zmarzty et al. 2025).
Classification. I have not followed the classification in Chase et al. (2002) since the four tribes recognised there do not map (in terms of monophyly) on to the tree, but rather, the recent classification of Zmarzty et al. (2025), although those authors observe that there are still possible problems with the circumscriptions of couple of their tribes and the placement of a couple of the genera.
Previous Relationships. The bulk of Achariaceae had almost universally been included in Flacourtiaceae s.l. until quite recently (e.g. Cronquist 1981; Takhtajan 1997).
Botanical Trivia. Immature fruits of Australian Ryparosa have the highest concentrations of cyanogenic glucosides known - 12 mg g-1 dry weight (Webber & Woodrow 2004).
Thanks. I thank Sue Zmarzty for comments.
[[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]: A = and opposite K; G [3]; exotegmen of ± cuboidal cells.
Age. The age of this node is (69-)66, 63(-60) Ma (Wikström et al. 2001: Goupiaceae sister to rest), (86-)79, 76(-71) Ma (Bell et al. 2011: Goupiaceae not included), or (102.9-)97.1(-90.6) Ma (Xi et al. 2012b: Table S7).
Evolution: Ecology & Physiology. Perhaps just coincidence, but Violaceae, Salicaceae and Passifloraceae all include nickel hyperaccumulators.
[Goupiaceae + Violaceae]: petiole with ± annular and wing bundles; cuticle waxes 0; anther connective ± developed apically; stigma proper ± punctate, receptive area small, appearing recessed/hollow.
Age. This node has been dated to (100.8-)92(-81.9) Ma (Xi et al. 2012b: table S7) or around 96.2 Ma (Tank et al. 2015: table S1, S2).
GOUPIACEAE Miers - Goupia Aublet - Back to Malpighiales
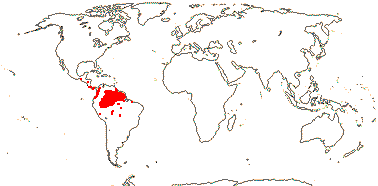
Trees; plants Al-accumulators, ?chemistry; vascular cylinder and pith 4-5-angled; vessel elements with scalariform perforation plates alone; petiole with inverted medullary bundle; branched sclereids +/0; hairs thick-walled, with pitted bases; orthotropic axes lacking expanded leaves, branch leaves two-ranked, lamina tooth ?type, secondary veins actinodromous, 3ary veins scalariform; inflorescences umbellate, axillary, pedicel articulation?; C induplicate-valvate, long, apical part narrow, differentiated; nectary annular; connective stout, shortly prolonged, with long hairs; pollen with endexinal folds; G [5], opposite C, placentation basal-axile, styluli short, adaxially channeled, on outer shoulders of carpels [ovary with roof], stigma type?; ovules few/carpel, ?morphology; fruit a berry; seeds not arillate, testa reticulate, both it and tegmen ca 3 cells across, exotegmen ridged, with 1 layer of ± laterally flattened sclereids, wall thickenings U-shaped; endosperm copious; n = ?
1 [list]/2. Central and N. South America. Map: from Tropicos (xii.2010).
Evolution: Divergence & Distribution. Diversification in the Goupiaceae clade seems to have slowed down (Xi et al. 2012b; Magallón et al. 2018).
Ecology & Physiology. Goupia glabra is a common and sometimes abundant member of the Amazonian tree flora, where it ranks #7 in terms of above-ground biomass and #10 in productivity (ter Steege et al. 2013; Fauset et al. 2015).
Chemistry, Morphology, etc.. Kubitzki (2013b) thought that the node was unilacunar, but Hoyos-Gómez (2015) found that it was trilacunar, the lateral traces departing from the central stele well before the central trace. It is often suggested that only seedlings have dentate leaves, those of the adult being entire, but leaves of flowering specimens are frequently toothed.
If Takhtajan (2000) is correct that there is a lignified endocarp, the fruit is technically a drupe.
For general information, inc. anatomy, see Hoyos-Gómez (2015) and Kubitzki (2013b); information on wood anatomy is taken from den Hartog and Baas (1978) and on pollen from Lobreau-Callen (1977, 1980) and Furness (2011).
The family is poorly known, especially embryologically.
Previous Relationships. Cronquist (1981) included Goupiaceae in Celastraceae, Takhtajan (1997) in Celastrales, A.-L. de Jussieu and others have placed it in Rhamnaceae. Furness (2011: pollen size) suggested that Goupiaceae were closest to the Lacistemataceae-Salicaceae clade.
VIOLACEAE Batsch, nom. cons. - Back to Malpighiales
Trees; vessel elements long to short with simple or long-scalariform perforation plates; petiole bundles arcuate; leaf teeth with a deciduous apex [Salicoid - ?level]; pedicels articulated; flowers weakly monosymmetric; K quincuncial; K persistent in fruit; exotesta subpalisade to tabular, ± thickened, (mesotesta sclerenchymatous), endotesta usu. crystalliferous; exotegmen cells tracheidal, lignified, thickened on all walls.
26/1126: [list]. World-wide.
Age. This node is around (86.8-)72.9(-57.4) Ma (Xi et al. 2012b: table S7).
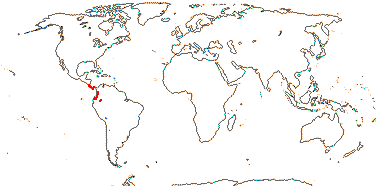
1. Fusispermoideae Hekking - Fusisperma Cuatrecasas
Pith with long cells (50-200 μm long], walls thin [2-3 μm across]; nodes 5:5; petiole with an elliptical medullary bundle, phloem internal; ?stomata; C contorted; nectary fleshy, 5-lobed, lobes alternating with A, filaments ± adnate to inner surface at indentations; anther thecae cordate/trapezoid, confluent apically?, connective with short paired fringed adaxial apical scales; capsule ca 3 mm long; seeds elongated, longitudinally winged, aril 0, ?exotegmen only moderately developed, of somewhat elongated cells; n = ?
1/3. Costa Rica, Panama, Columbia, Peru (Amazonas). Map: from S. Hoyos-Gómez (pers. comm.).
2. Violoideae Beilschmied
Pith with cells to ca 50 μm long, walls 4-5 μm thick; nectary opposite A; connective with large abaxial scale, as long as and broader than anther, ± free, margin entire or erose; (capsule with explosive dehiscence).
2A. Briberia Wahlert & Ballard
Vessel elements?; anther connective subapical, thecae visible from abaxial side; style broadening apically; ovule 1/carpel; n = ?
1/3. Costa Rica to Peru.
2B. The Rest. —— Synonymy: Alsodeiaceae J. Agardh, Leoniaceae A. L. de Candolle
(Annual to perennial herbs; lianas); plants often Al accumulators; cyclotide proteins +, tannins 0 [woody members?]; calcium oxalate often as crystals; (petiole bundles arcuate); stomata also para- or anisocytic; leaves spiral or two-ranked (opposite), lamina margins involute, colleters +, (stipules petiolar; lobed); flowers often strongly monosymmetric, (papilionoid); C quincuncial, abaxial C spurred or not; A (3), basally connate or not, nectariferous appendage on abaxial surface of filaments (on 2 abaxial A only), (filaments connate), anthers connivent, thecae (horizontal), obscured by connective from abaxial side, (connective ± 0); pollen ((3-)4-5-(-6) colporate - Viola); G [(2-5)], (when 5 opposite K), styles separate or style +, straight or curved, apex subcapitate, asymmetric or not, (stigma hollow); ovules 1-many/carpel, with zig-zag micropyle (endostomal), outer integument 2-4 cells across, inner integument ca 3 cells across, parietal tissue 2-3 cells across, nucellar cap ca 2 cells across, hypostase +; (fruit a berry, nut); seeds (winged), (not arillate/carunculate), (exotesta ± thick-walled, lamellate), mesotesta (sclerenchymatous), (cells periclinally elongated - Viola), endotesta crystalliferous (not - Ionidium), (exo)tegmic fibre layer 1-3 cells across, endotegmen thick-walled, not lignified or elongated; embryo (small), green [Viola], (suspensor 0), cotyledons accumbent/(oblique), (with sinuous foldings); n = (5-)6(7)8(-13+); nuclear genome [1C] (1973-)1404(-1144) Mb.
22-32/1,180: Viola (727, of which 43 undescribed), Rinorea (?230-250), Hybanthus (?120), Pombalia (44), Afrohybanthus (25+). World-wide; woody taxa esp. in the lowland tropics, mostly the Neotropics. Map: from Hultén (1958, 1971), Hultén and Fries (1986), Hekking (1988), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003) and Australia's Virtual Herbarium (consulted i.2013) - incomplete for South America. [Photo - Leonia, Alexis fruit and flowers, Viola.]
Evolution: Divergence & Distribution. Marcussen et al. (2012, especially 2014) begin to disentangle the complex reticulate history of the polyploid northern hemisphere species of Viola subgenus Viola where reticulations as old as ca 29 Ma have been detected (the crown age for the genus is ca 31 Ma - see also Marcussen et al. 2022); there have been 16-20 or more major allopolyploidization events, section Abyssinicum being 12x and some species up to over 20x. Indeed, only sections Rubellium and Chamaemelanium, both n = 6, are diploid, as are the very few counts from subgenus Neoandinium (n = 7) (Marcussen et al. 2022). The origin of the genus seems to have been in South America, with two shifts to North America, and speciation in subgenus Viola has commonly been by allopolyploidization. There is a radiation of Viola on Hawaii; these are also polyploids and include secondarily woody species; their ancestry is to be sought among west North American species, and they reached Hawai'i via long distance dispersal (Marcussen et al. 2012). The genus as a whole may have had its origin in Andean South America (Ballard et al. 1998; Marcussen et al. 2022), and there the large and poorly-known section Andinum (= subgenus Neoandinium), with perhaps one fifth (ca 111 spp., plus 40 to be described) of the genus and including some remarkable rosette- and tussock-forming species (J. M. Watson et al. 2022), is to be found.
Van Velzen et al. (2015) optimized the evolution of a number of characters in African Rinorea.
Ecology & Physiology. Violaceae are notably common in terms of both numbers of species and individuals with stems at least 10 cm across in the Amazonian tree flora, and they are disproportionally common in the 227 species that make up half the stems in Amazonian forests (ter Steege et al. 2013). Rinorea is common and can dominate in the lower strata of African forests, and several species may grow together (van Velzen et al. 2015).
Metal hyperaccumulators are quite common in the family, and include both herbaceous - e.g. Hybanthus on New Caledonia (Gei et al. 2020) - and woody members (Brooks 1998 for a summary; Marcussen et al. 2022); the genera involved are Viola subgenus Viola, Afrohybanthus, Agatea, Pigea and Rinorea, immediately unrelated groups.
Pollination Biology & Seed Dispersal. In the northeast U.S.A. oligolectic bees are notable visitors to Viola (Fowler 2016). Cleistogamy is widespread in this genus.
The largely temperate Viola is myrmecochorous (Lengyel et al. 2010). However, most species are really diplochorous, in that their seeds are initially dispersed by ballistic discharge, and only subsequently by ants. Relatively few species are strictly myrmecochorous, and in such plants the capsules are held close to the ground, the seed and elaiosome are larger, etc., making the seeds available for the ants (Beattie & Lyons 1975). Myrmecochory, as well as dispersal by larger animals and by wind, also occurs in more tropical taxa.
Plant-Animal Interactions. Violaceae are the preferred food plants for the caterpillars of the majority of fritillaries, Nymphalidae-Argynnini (Simonsen 2006; Nylin et al. 2014). In Africa, caterpillars of perhaps half the species of Cymothoë (ca 75 spp., Nymphalidae-Limenitidinae) feed on Rinorea, while Salicaceae and Kiggelariaceae (= Achariaceae), both quite closely related to Violaceae, are also reported to be hosts. Although individual species of butterfly and plant may be closely associated, the current geography of the two may differ from that in the past (McBride et al. 2009).
Glucosinolates were found in the roots of Rinorea subintegrifolia, from Gabon, but not in the leaves or stems of R. woermanniana (Montaut et al. 2016).
Genes & Genomes. Viola (Rinorea sister) has a genome duplication - or perhaps two, one quite recent (Cai et al. 2017/18).
Viola tricolor, the pansy, and the related V. arvensis were important in early studies of genetics and speciation.
Chemistry, Morphology, etc.. Inulin has been reported from Hybanthus (Beauvisage 1889); for cyclotides, widely distributed in the family, see Burman et al. (2010). From the description of the root of Ionidium (= Hybanthus) ipecacuanha by Beauvisage (1889), the cork cambium may be mid-cortical or superficial. Viola has storied cambium.
For information on the flowers of Fusispermum, see Cuatrecasas (1950) and Hekking (1984), the former describes the scales as being ventral appendages of the connective. Feng and Ballard (2005) suggested that even those Violaceae with polysymmetric adult flowers were monosymmetric earlier in development, so "flowers monosymmetric, at least in bud" may be an apomorphy for all/most of the family (see also Arnal 1946 for monosymmetry). The anthers and stigmas of many species are very complex (e.g. Kuta et al. 2012: Viola); there may be a nectariferous connective gland (see de Aleida et al. 2023), but its distribution is unclear. The seeds of Anchietea (winged) and Decorsella (round, red) mature exposed on the open carpels; in the latter genus they look like a bunch of little red grapes.
For general information, see Ballard et al. (2013), also Espeut (2022: earlier classifications, esp. Viola), Hoyos-Gómez (2015) and Wahlert et al. (2017), both focus on basal pectinations, and Munzinger and Ballard (2003: family key, but dated). An unpublished thesis by Feng (2005) includes a phylogeny of the family and details of the floral development of seven genera. For chemistry, see Hegnauer (1970, 1990), for growth and branching patterns, for pollen morphology, see Mark et al. (2012: Fusispermum has two pollen size classes), and see Hekking (1988) for embryology, etc., Singh (1970), Singh (1963), Singh and Gupta (1967), and Dathan and Singh (1974) for seed anatomy, etc., and Raju (1958) for fruit dehiscence; see also Leins and Erbar (2010: flowers of Viola).
Phylogeny. There is good support for the relationships [Fusispermum [Rinorea apiculata group (now = Briberia) [Rinorea s. str. [[Viola etc.], [Leonia, etc.], [Melicytus, etc.]]]]] (Tokuoka 2008, see also Feng & Ballard 2005; Ballard et al. 2009; Wahlert & Ballard 2012; esp. Wahlert et al. 2014). The para-/polyphyly of Rinorea is well established, but some major polytomies make it difficult to clarify details of relationships much further. Viola is included in a well-supported clade the other members of which are woody and also have strongly monosymmetric flowers - with the exception of most species of Allexis (?buzz pollinated), which is is sister to the rest of that clade; other taxa with strongly monosymmetric flowers are not immediately related (Wahlert et al. 2014). Hybanthus pops up all over the place in both the Leonia (the type of Hybanthus is there) and Melicytus clades, but unfortunately relationships in the latter clade are not well understood (Wahlert et al. 2014). Some of the relationships suggested by M. Sun et al. (2016) are somewhat different.
Bakker et al. (2006b), Wahlert and Ballard (2012) and in particular van Velzen et al. (2015) discuss relationships in the speciose African Rinorea, where the clade [R. ?exappendiculata + R. woermanniana] may be sister to the rest of the genus. Ballard et al. (1998) looked at relationships within Viola using ITS data, and to summarize: Sections largely seemed to be in a mess. Relationships in the Calcarata complex of section Melanium of Viola were poorly resolved, there may well be hybridization, and there is no evidence that the complex is monophyletic (Krause & Kadereit 2020). Indeed, hybridization, some very old and with ploidy levels up to 20, is common in the genus, at least in subgenus Viola (Marcussen et al. 2012, 2014, 2022).
Classification. The current (2022) infrafamilial classification is insupportable, thus the large genus Hybanthus is to be cut up into nine genera, perhaps, and Rinorea is also to be divided. Ballard et al. (2013), Wahlert et al. (2014) and Flicker and Ballard (2015) have begun the process of dismemberment.
Marcussen et al. (2022) have divided Viola into two subgenera, Viola, with 20 sections and 18 subsections, and the little-known Neoandinium, with 11 sections.
[Passifloraceae [Lacistemataceae + Salicaceae]] / Salicoids: plastid rps7 and rpl 32 genes to nucleus, rps16 replaced by nuclear gene.
Age. The age of this node may be around (66-)63, 60(-57) Ma (Wikström et al. 2001: Hymenanthera included), (100.5-)94.4(-87.5) Ma (Xi et al. 2012b: table S7), ca 98.8 Ma (Tank et al. 2015: table S2) or ca 93 Ma (M.-M. Li et al. 2019).
Evolution: Genes & Genomes. For plastid gene movement, see Shrestha et al. (2020).
Phylogeny. This clade has strong support in the analysis of Xi et al. (2012b).
PASSIFLORACEAE Roussel, nom. cons. - Back to Malpighiales
Cyclopentenoid cyanogenic glycosides and/or cyclopentenyl fatty acids + [esp. tetraphyllin; ?derived from valine and isoleucine]; (plant with unpleasant smell); (colleters +); leaves spiral, (foliar glands +); K often ± mucronate/narrowly acute; K + C forming a tube, corona or scales towards mouth (0), K ± petal-like, C base rather broad; pollen grains with ± reticulate surface; styluli +, stigma expanded; micropyle bistomal, ± zigzag; seed coat ridged, endotestal cells massive, exotegmen sclereidal oblique-palisade, endotegmen persistent; endosperm persistent, oily; x = 7 (?6), nuclear genome [1 C] (0.035-)1.098) pg; (plastid transmission biparental/paternal), atpF intron lost.
27 [list: to tribes]/1,035. Tropical, esp. America and Africa, also warm temperate - 4 subfamilies below.
Age. This node has been dated to around (38-)36, 32(-30) Ma (Wikström et al. 2001: [M + T] P]), (59-)47, 43(-32) Ma (Bell et al. 2011: [T [M + P]]), (80.3-)62.8(-47.7) Ma (Xi et al. 2012b: table S7) and ca 84.7 Ma (L. Cai et al. 2025b).
1. Malesherbioideae Burnett - Malesherbia Ruíz & Pavón —— Synonymy: Malesherbiaceae D. Don, nom. cons.
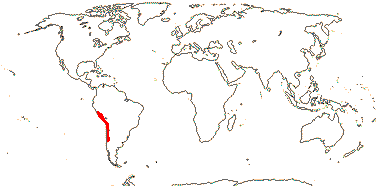
Herbaceous or subwoody, (caespitose); hairs conspicuous, multiseriate, often glandular; plant with unpleasant smell; tannins?; (cork cortical); nodes also 1:1; lamina margins often deeply lobed, (margins), (stipules foliaceous, to deeply 3-lobed/0); inflorescence with lateral cymules, (flowers single); K + C tube long, with 10 [?always] longitudinal veins, K valvate, C valvate; androgynophore +; nectary at base; (G [4]), gynophore +/0, styles slender, stigmas capitate-clavate, ?type; ovule with large protrusion at chalazal end; K + C tube persistent; seeds spherical, 1.3-2.7 mm long, ridged-foveolate, raphe prominent, aril 0; exotestal cells in vertical and horizontal series, then layer of massive cells, thickening U-shaped, protruding into palisade layer, then ca 3 layers of small unthickened cells; endosperm type?; n = 7.
1/27. South America from Peru southwards, esp. N. Chile. Map: see Ricardi S. (1967: Fig. 1) and Gengler-Novak (2002). [Photo - Habit]
Age. Crown-group Malesherbioideae are ca 25 Ma (Guerrero et al. 2013).
[[Turneroideae + Pibirioideae] Passifloroideae]: extrafloral nectaries + [often on stipules/petiole/base of lamina]/0; lamina vernation conduplicate; anthers long; tapetum amoeboid; aril ± raphal.
Age. This node has been dated to ca 73.2 Ma (Muschner et al. 2012) and (66.8-)50(-35.5) Ma (Xi et al. 2012b: table S7).
[Turneroideae + Pibirioideae]: ?
2. Turneroideae Eaton —— Synonymy: Piriquetaceae Martynov, Turneraceae Candolle, nom. cons.
Herbaceous or woody; plant with unpleasant smell; ellagic acid 0; cortical vascular bundles [= leaf traces] common; stomata various; hairs tufted/stellate, (glandular); colleters +; stipules 0 (+ - e.g. Erblichia); inflorescences racemose, (epiphyllous - some Turnera and Piriqueta); bracteoles +, often large; (flowers heterostylous); (hypanthium +); glands or corona at mouth of K + C tube (0), C contorted [left-handed], ± clawed, deliquescent; nectary near base of tube (on K/filaments); (androgynophore +); (G [2]), (half inferior), stigmas laterally expanded, concave, often ± penicillate; ovule (1/gynoecium, basal - Stapfiella), outer integument ca 2 cells across, inner integument 3(-4) cells across, (nucellar cap ca 2 cells across), parietal tissue 2-5 cells across, suprachalazal zone massive, hypostase +/0; K + C tube deciduous; seed ± elongated, curved, 1.2-5 mm long, surface ridged-reticulate or striate, aril + (fimbriate); exostome persistent, testa with stomata; n also = 5 (13); plastid transmission biparental [T.].
12/233: Turnera (130), Piriquetia (44). Tropical to warm temperate America and Africa (inc. Madagascar and Rodriguez I.). Map: from Wickens (1976), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003), Heywood (2007, in part), Arbo (2008) and Thulin et al. (2012a - T. ulmifolia widely naturalized). [Photo - Flower.]
Age. Crown-group Turneroideae are dated to (42.5-)32.3(-42.5) Ma (Thulin et al. 2012b) or (40.3-)27.8(-14.6) Ma (Xi et al. 2012b: table S7).
Thanks. To M. M. Arbo for species numbers, etc..
3. Pibirioideae Chase & Christenhusz - Pibiria flava Maas
Shrub; ?nodes; stomata anisocytic; extrafloral nectaries 0, stipules 0; inflorescence fasciculate, ?cymose, bracteoles ?0; K + C not forming a tube, K imbricate, not mucronate, C imbricate, broadly attached, long-lived, corona 0; anthers sagittate, versatile; pollen surface rugulate; ?nectary; stigmas ± punctate; 1(-2) ovules/carpel, ?embryology; ?fruit; ?seeds; ?n.
1/1. Guyana.
4. Passifloroideae Burnett
Woody; anthers versatile; G [(2-)3(-7)], styuli ± separate, long [>2/3 length of G]; exotestal cells not in lines.
17/706. Tropics to warm temperate, especially Africa and America - three tribes below.
4A. Jongkindieae Breteler & F. T. Bakker - Jongkindia mulbahii Breteler & F. T. Bakker
Small tree; plant glabrous; leaves 2-ranked, teeth with abaxial gland; flowers single, axillary, bud covered in resin; pedicel articulated; 4-merous, corona 0, K imbricate, ± fimbriate, apex not mucronate, C valvate, apex fimbriate; ?nectary; stigma peltate; ovules straight[?]; testa rugose; embryo long, cotyledons 1/2 or more its length; n = ?.
1/1. West Africa, Liberia. Map: Breteler et al. (2022: Fig. 1).
[Paropsieae + Passifloreae]: (lamina margins entire); ?inflorescence; flowers (3-)5-merous; K mucronate, corona of (1-)2-several rows of filaments or membranes; nectary ± on K + C tube; anthers versatile; (styluli short), stigma/s capitate-clavate; (apex of nucellus pushing into micropyle); seeds flattened, 1.4-14 mm long, surface ± foveolate, reticulate-foveolate, transversally grooved, bony; endosperm foveolate/ruminate; n also = 6.
Age. The crown-group age of this clade is around 65.5 Ma (Muschner et al. 2012), (29-)27, 26(-24) Ma (Wikström et al. 2001) or (42.6-)26.6(-11.6) Ma (Xi et al. 2012b: table S7).
Martínez (2017) described Late Eocene (45-34 Ma) seeds from Columbia as Passifloroidesperma sogamosense and placed the species in crown-group Passifloroideae. Seeds described as Passiflora axsmithii and placed in supersection Laurifolia, series Tiliifolia from 25-23.8 Ma Late Oligocene deposits in Mississippi have a ruminate endosperm, palisade seed coat, etc. (Stults et al. 2024).
4B. Paropsieae de Candolle —— Synonymy: Paropsiaceae Dumortier, Smeathmanniaceae Perleb
Trees or shrubs; vessel elements in multiples, perforation plates simple/scalariform; leaves reduced [orthotropic axes], two-ranked [plagiotropic axes], lamina with glands especially on margin and apex, (stipules 0); inflorescence racemose; androgynophore +; A (-30, partly connate); pollen (3-)6[Paropsia]-colporate, (5-porate, surface echinate-granulate - Barteria), endaperture various, (operculum) pontoperculum +, sexine reticulate, (5-porate, sexine echinate (apices of spines rounded), granulate - Barteria); nectary 0 (annular); gynophore + (0); (styluli short - Paropsia), (style undivided, stigma massive, capitate - Barteria); ?ovules; (fruit dry, indehiscent); seeds scrobiculate; n = ?
5/22: Paropsia (12), Barteria (6: inc. Smeathmannia). Africa-Madagascar, esp. West Africa, also the Malay Peninsula. Map: from Sleumer (1970), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003) and de Vos and Breteler (2009).
4C. Passifloreae de Candolle —— Synonymy: Modeccaceae Horaninow
Vines or lianas, (herbs, shrubs), (trunk/stem base much swollen); tendrils + [modified branches], simple (branched), (0); cyclopentenoid cyanogenic glycosides diverse, flavonols +, ellagic acid +/0, tannins 0; wood often fluorescing; (vascular tissue deeply fluted and lobed), (successive cambia), (ectopic cambia); colleters +; supernumerary buds +, superposed; leaves (palmately compound), lamina venation palmate, vernation conduplicate-plicate/plane, (margins entire), glands on lamina surface, paired glands on petiole/0; plant (dioecious), inflorescence cymose/main [primary] axis 0/(flowers single); (flowers monosymmetric); C (0, 1), corona of (1-)2-several rows of filaments or membranes (0), nectary ± on K/C tube; (androgynophore +); A (-8), (basally connate), (basifixed); tapetal cells binucleate; pollen 3(-12-colporate - Passiflora); (G [4-8]), (gynophore +), stigmas (divided - Adenia), with multicellular papillae; ovules with outer integument 2-5 cells across, inner integument 3-5 cells across, parietal tissue 6-20 cells across, (nucellar cap ca 2 cells across), hypostase +, funicle often long; fruit a berry, (capsule - P. section Xerogona); seeds hairy or not, often sculpted; testa multiplicative, sarcoexotestal, or exotesta palisade, endotesta crystalliferous, lignified or not; cotyledons accumbent; n = 6(7-)9(-12); nuclear genome [1 C] ca (675-)1661(-2905) Mb; plastid transmission biparental [P.], two inversions in the LSC region, rps19 gene incorporated into the IR, rpoA gene non-functional, rpl22 gene transferred to nucleus, etc. [P.].
10/725: Passiflora (571), Adenia (100). Tropics to warm temperate, especially Africa (most genera) and America (most species). Map: from van Balgooy (1975), Fl. Austral. vol. 8 (1982) and Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003). Photo: Collection.
Age. An age for this clade may be (80.9-)71.3, 66.4(-49.1) Ma (Hearn 2006, but c.f. topology) or ca 49.5 Ma (Muschner et al. 2012).
Passiflora bulgarica (subgenus Decaloba, supersection Cieca) has been described from Middle Miocene deposits in Europe (Hermsen 2021). Seeds of P. axsmithii, subgenus Passiflora supersection Laurifolia series Tiliifolia, with ruminate endosperm, were described from rocks 25.0-23.8 Ma from Mississippi (Stulz et al. 2024), while seeds identified as Passifloroideae, with pitted-reticulate coats and from ca 24.3 Ma, have been found in Peruvian Amazonia (Antoine et al. 2021).
Evolution: Divergence & Distribution. For a review of the fossil record of Passifloroideae, see Hermsen (2021). Note that the ages og taxa in Passiflora conflict other ages for the genus.
Weber and Agrawal (2014) suggested that the evolution of extra-floral nectaries in Turnera was associated with an increase in diversification rates, while extrafloral nectaries may be a key innovation in Passiflora (Krosnick et al. 2011: E. O. Silva et al. 2016: anatomy and secretions). The rate of diversification may have increased in Passifloroideae or the [Turneroideae + Passifloroideae] clade (Xi et al. 2012b).
Hermsen (2023) thought that Passiflora might have dispersed to Europe via the North Atlantic land bridge and to the Antipodes-Southeast Asia via Antarctica. Section Decaloba may have originated in Central America; one clade diversified there, moving throughout the Caribbean and into North America, the other clade, initially northern Andean, moved elsewhere in South America and on to the Galapogas (Acha et al. 2021). The centre of diversity of Passiflora in terms of species numbers/area is Ecuador (Kuethe et al. 2025).
Ocampo Pérez and Coppens d'Eeckenbrugge (2017) surveyed the morphological variation in Passiflora comparing it with groupings evident in molecular analyses. They looked at 127 characters and with a focus on the analysis of 24 quantitative descriptors, and found that agreement between morphology and molecular groupings was on the whole good except in the bee-pollinated taxa of subgenus Passiflora.
Sader et al. (2019) examined changes of chromosome number in Passiflora (x = 6), where they estimated diversification began (48.2-)42.9(-37.7) Ma, integrating it with changes in genome size and general flower type. They suggested that Miocene diversification in subgenus Passiflora could be linked with increasing chromosome numbers (by dysploidy) and genome size; polyploidy was not associated with diversification increase (Sader et al. 2019). There are a number of changes in the plastome that may end up being apomorphies, for example, Pacheco et al. (2020b) noted that two inversions in the LSC region were shared by Adenia and Passiflora and, for similar reasons, that the IR had expanded with the resultant duplication of the ycf1 gene (see also Shrestha et al. 2019).
Hearn (2009b) suggested that where vascular strands and associated parenchymatous storage tissue in root and/or stem developed varied in Adenia, hence helping to generate the diversity of growth forms in that genus, and Hearn (2013) emphasized that transport, support, and storage functions in the plants were semi-independent, perhaps facilitating evolutionary change.
Malesherbia grows in more or less mesic areas to deserts, i.e. areas with ≤50 mm rain/year, in the Atacama-Sechura area of western South America, and are likely to have invaded the deserts ca 20 Ma after the onset of aridity ca 33 Ma - interestingly, the region has been semiarid, with ≤250 mm rain/year, since the Late Jurassic ca 150 Ma (Guerrero et al. 2013).
Arbo and Espert (2009: relationships from a morphological analysis - see below) discuss the morphology and biogeography of Turnera. Thulin et al. (2012b) suggest that Mathurina, the only Turneroideae with wind-dispersed seeds, is older than Rodrigues Island, to which it is now restricted. The majority of genera of Turneroideae are African, but include only a few species; most species are New World, most in Turnera itself, but also Oxossia, and there are three or so links across the South Atlantic; biogeographical relationships, which can be summarized as [[New World -> movement to Old World] [New World (1 species) + Old World]], were obscured by the pre-2012 taxonomy (Thulin et al. 2012b).
It has been suggested that there is floral mimicry between Turnera and Malvaceae in Argentina (Benitez-Vieyra et al. 2007).
Rocha et al. (2019: Fig. 6) discussed inflorescence evolution with a focus on Turneroideae; note that their peduncle (bottom left diagram) is part of what is called a pedicel here. For androgynophore evolution, see Tokuoka (2012).
Ecology & Physiology. About 625 species of Passifloraceae-Passifloreae are vines/lianas of one sort or another (estimate derived from Feuillet & McDougal 2007; see also references in Schnitzer et al. 2015), so they make up a major group of climbers. Passiflora is one of the ten most diverse genera of climbers in the world, and Passifloraceae are one of the most speciose families of climbers in the Neotropics (Sperotto et al. 2023). The tendrils, usually unbranched, spiral in opposite directions along their length, reversing at what is called the perversion (Goriely & Tabor 1998; Gerbode et al. 2012); they twine around their support, although in some species the tendrils may also be adhesive (Bohn et al. 2015).
Passiflora foetida (section Dysosmia) may be able to take up nitrogen from insects that become stuck on the hairs of the three highly fimbriate "bracts" (= bract + two bracteoles) surrounding the flower (Radhamani et al. 1995); this should be confirmed.
Species of both Turnera and Piriquetia from Goiás, Brazil, can accumulate high concentrations of nickel (Reeves er al. 2007).
Pollination Biology & Seed Dispersal. A complex corona described by a similarly complex set of terms (for further discussion, see Chemistry, Morphology, etc. below)) is particularly conspicuous in Passiflora, and it is variously involved in nectary protection and presentation, tube formation, etc.; a more or less well developed K/C tube with some kind of corona is common in Passifloraceae as a whole. For the pollination of some 37 species of Andean Passiflora supersection Tacsonia by the hummingbird Ensifera ensifera, which has the longest bill of any hummingbird, see Abrahamczyk et al. (2014: many support values in the phylogeny low), and there have been reversions to shorter tube lengths; the Tacsonia clade is ca 8.4 Ma, Ensifera is about a million years younger (Tacsonia ca 11.6 Ma, Ensifera ca 10.7 Ma - Soteras et al. 2018). Aetanthus and Tristerix (both Loranthaceae), Salvia (Lamiaceae) and Brugmansia (Solanaceae) are also members of the same pollination guild (Soteras et al. 2018). There is modest movement of the androgynophore stimulated by and towards the pollinator in species of Passiflora section Xerogona (Scorza & Dornelas 2014). Sazima and Sazima (1978) note that the bat-pollinated flowers of Passiflora mucronata become zygomorphic as the stamens move after the flowers open (see also Endress & Matthews 2006a). De Mattos et al. (2022) suggested that pollination by bees may be the original condition, and they looked at nectaries and nectar production in the context of the various pollinators, i.e. birds, bats and bees. It would be interesting to know what pollinates P. nosferatu, which has but a single petal - there is only one other species with a single petal - and reddish flowers that open in the afternoon... (Kuethe et al. 2025).
Heterostyly is common in Turnera, Piriqueta, and some other Turneroideae; a supergene is involved (Barrett & Shore 2008; Cohen 2019; Gutiérrez-Valencia et al. (2021) and especially Shore et al. 2019). Endress et al. (2013) suggested that there was also heterostyly in Malesherbioideae, but Shore et al. (2006) thought that it was not found there.
Most Adenia may be dioecious, however, serial monoecy, the plant initially producing male flowers, then female flowers as it becomes robust, but male flowers again if conditions deteriorated, has been noted in some species (Condon & Gilbert 1988).
There is myrmecochory in Turnera (Lengyel et al. 2010), numerous species of ants playing a variety of roles may be associated with species such as T. ulmifolia (Rico-Gray & Oliveira 2007 and references). Some species in which the aril is fimbriate may be dispersed by wind (Arbo et al. 2015).
Plant-Animal Interactions. Passiflora in the New World - only 5% of its species are found in S.E. Asia-Malesia and Oceania - has a very close association with Heliconius butterflies (Nymphalidae-Heliconiini) (e.g. Ehrlich & Raven 1964; Quicke et al. 2025). These butterflies are noted for forming complex mimicry rings (Merrill et al. 2015 for a summary), and Hoyal Cuthill et al. (2019) provide an example of co-evolution in the Müllerian mimicry developed in such situations; hybridization and introgression are widespread in the genus (Edelman et al. 2019; Cicconardi et al. 2023). There are some 525 species of Passiflora and 45 species - but ≥200 subspecies - of Heliconius (de Castro et al. 2018). Heliconius caterpillars, and those of Heliconiini in general, use Passiflora, as well as its relatives like Dilkea and Mitostemma, as their only food source (Benson et al. 1975). The plants vary greatly in leaf morphology (see also below) and have foliar glands of various kinds (Gilbert 1982; Cusset 1965; McDougal 1994 for literature); some of the glands are yellow-coloured egg mimics (Vanderplank 2007 for references), and since the butterflies tend to lay eggs on plants that they think lack eggs, the glands may protect the plant. The leaves may (also) have epithelial extrafloral nectaries and attract ants that defend the plants (Krosnick et al. 2011 for a summary) and/or hooked hairs that can penetrate the cuticle of the caterpillars (e.g. de Castro et al. 2018), however, caterpillars of species like H. charithonia deal with such hairs on P. lobata by covering them with silk mats, biting off the tips by of the trichomes, etc. (Cardoso 2008). Chemical defences include β-carboline alkaloids, a variety of cyanogenic compounds, etc. (Gilbert 1982; Spencer 1988; Jaroszewski et al. 2002; Bak et al. 2006; de Castro et al. 2018; Quicke et al. 2025). These alkaloids and cyanogenic compounds may be sequestered by the Heliconius caterpillars feeding on Passiflora and used in their defence and/or even as nitrogen sources, and they can also be synthesized by the caterpillar; caterpillars that have lost the ability to synthesize cyanogenic cyclopentenoid glycosides may be restricted to particular species of Passiflora that can provide the glycosides that they need for their defence (Engler-Chaouat & Gilbert 2007; Opitz & Müller 2009). However, the relationships between specialist and generalist Passiflora herbivores, sequestration and/or glycoside synthesis, and the overall amount of cyanogenic glycosides in the body of the caterpillar is complex (Beran & Petschenka 2022).
Female Heliconius butterflies in particular are noted for their ability to find Passiflora plants and to remember where they are; they have very big mushroom bodies, involved in memory and learning, in their brains (see also Cicconardi et al. 2023; F. A. Young & Montgomery 2020), and also very large eyes, and they also seem able to sense plant chemistry, for instance when they tap the leaves with their antennae (e.g. Gilbert 1982; de Castro et al. 2018). However, their ability to distinguish between different leaf shapes does not necessarily translate into changes in egg-laying activities (Dell'Aglio et al. 2016). In any one locality a single species of butterfly is often associated with a single species of Passiflora, although there are many exceptions (Benson 1978; Merrill et al. 2013; de Castro et al. 2018). Heliconius lays eggs on young parts of the plant, but some heliconiines, Dryas iulia, for example, lay eggs on older leaves (Benson 1978; de Castro et al. 2018). Adult butterflies, at least in the Melpomene clade, are also closely associated with Psiguria and Gurania (Cucurbitaceae) and relatives, from the staminate flowers of which they obtain pollen (female butterflies take more pollen than males); members of the Erato clade visit flowers of other families. Pollination by Heliconius butterflies is likely to be a derived feature here, the butterflies collecting and digesting pollen, although details as to how they do the latter are unclear (Cicconardi et al. 2023; see also Young & Montgomery 2020); the ability to do this seems to have been lost in the Aeode clade. Interestingly, the home ranges of members of the Melpomene clade may be centred on the plants - they are territorial, trap-lining insects. The plant gets pollinated, the pollen sticking to the base of the proboscis of the butterfly, but also, mixed with saliva, the pollen is a source of amino acids that are involved both in egg-making and, as cyanides, also in the defence of the pollinating insects. The butterflies are very long-lived, up to six months or so (e.g. Gilbert 1972, 1975; Boggs et al. 1981; Eberhard et al. 2009; Steele 2010; Corlett & Primack 2011; Young & Montgomery 2020). Note that pollen-feeding is extremely uncommon in other lepidoptera with ptobosces, and within Heliconiinae is restricted to Heliconius...
The butterflies may have diversified on the foothills and lower slopes of the eastern Andes from Peru northwards (Rosser et al. 2012). However, Hermsen (2021) noted that seeds attributed to Passiflora supersection Cieca had been reported from Europe, although that group is not found there now (Fordyce 2010; Kozak et al. 2015 for references and diversification rates, but see Brower & Garzón-Orduña 2018 for a critique).
Given the very close association of Heliconiini in particular with Passiflora and relatives, basically obligatory for both parties, ages of the main protagonists are of interest. It has been estimated that crown Heliconius is less than 12.5 Ma (Merrill et al. 2015) or (20.4-)18.5(-16.5) Ma (stem) and (13.4-)11.8(-10.5) Ma (crown: Kozak et al. 2015), however, crown-group Heliconiini may be around 26 Ma (Kozak et al. 2015). Young and Montgomery (2020) estimated that the age of Heliconius was only some 18 Ma, while Cicconardi et al. (2023) suggested that the age of Heliconiinae was (55.5-)45.3(-35.9) Ma, that of Heliconiini, particularly associated with Passiflora, was some (32.7-)ca 26(-20.7) Ma, while that of the speciose Heliconius itself was (13.8-)9.6(-8.8) Ma. Ca 40.5 Ma is the age estimated for crown Passiflora, although diversification within its four main clades did not begin (subgenus Decaloba) until ca 29 Ma (Muschner et al. 2012). Hermsen (2021) evaluated the fossil record of the genus, and noted that seeds attributed to Passiflora supersection Cieca were known from mid-Miocene deposits in Europe.
The larvae of some Acraeinae, also nymphalids, and also of brightly-coloured Notodontidae-Dioptininae moths are also often found on Passiflora (J. S. Miller 1992; Silva-Brandão et al. 2008), and at least the former are also found on Barteria (Paropsieae); MacDougal (1994) lists other herbivores that eat Passiflora. Turneroideae are the hosts of caterpillars of several genera of Nymphalidae, alternate hosts include other members of the clade with parietal placentation (Arbo 2006 and references). For the trenching behaviour of foliovores on Passiflora, see Dussourd (2016 and references).
For other similar tropical systems, see Eugenia, Inga, Ocotea, Piper, Protium and Bursera, Psychotria, and sundry Solanaceae - the discussion about Inga is perhaps particularly detailed. For general patterns of diversity in seed plants, with the greatest diversity being at low latitudes, a pattern that may in part be driven by the factors just discussed, see elsewhere.
Dias et al. (2020) looked at the marginal foliar glands in Passiflora and the intersection between hydathodes, colleters and extrafloral nectaries (see Rios et al. 2020 for leaf teeth and colleters in Turnera - general survey of such structures would be useful); ants may be attracted to the exudate. Details of the association between the African ant-plant Barteria fistulosa and the ant Tetraponera aethiops are given by Dejean et al. (2008). The evolution of this association, which involves all four species of Barteria and both specialist and generalist species of ants, is complex (Peccoud et al. 2012; see also Davidson & McKey 1993: ant species replace each other within the association?; Kokolo et al. 2019). Ascomycete fungi inhabiting the domatia are an immediate source of nitrogen for T. aethiops (Blatrix et al. 2012); for information about the fungi, Chaetothyriales, see Vasse et al. (2017).
Vegetative Variation. Anatomically, the old Flacourtiaceae-Paropsieae (Barteria, Paropsia, etc.) and Passifloreae (Chase et al. 2002) are rather similar. Indeed, the major variation in Passifloroideae seems to be associated with habit, lianas versus trees, and as might be expected the stem anatomy of the former tends to be rather distinctive, although not always (Ayensu & Stern 1964; Rajput & Baijnath 2016).
There is extensive foliar variation in Passiflora in particular, variation evident both when comparing leaves produced at successive nodes in an individual after germination (heteroblasty) and also between species that is especially evident when comparing adult plants; seedlings can be difficult to identify (Chitwood & Otoni 2017b; also 2017a: different approaches to describing Passiflora leaf shapes). In species of Passiflora with strongly bilobed leaves, vernation may be modified conduplicate: When folded in bud, the blade makes a V with an inverted V at the end of each arm. For more on vegetative variation in the genus, see also references in MacDougal (1994).
In Passiflora and its immediate relatives stem collenchyma, cymose inflorescences or branches may develop from the accessory (superposed) bud. Tendril morphology is discussed by Cusset (1968); the tendrils are stem tendrils, modified axillary shoots, and their prophyllar buds may give rise to single flowers, 3-flowered cymes, or even additional tendrils, as in the branched tendrils of Passiflora subgenus Deidamioides. These prophyllar buds are ?usually non-basal, the terminal bud forming the tendril (Hernandes-Lopes et al. 2019). Accessory buds are common in these taxa with non-basal prophyllar buds since the latter are "used up" in producing the inflorescence/tendril (see also Hernandes-Lopes et al. 2019) and so cannot play any role in regeneration or vegetative branching in general. There is some debate as to whether the tendril is the pedicel of a terminal flower or the ?axial terminus of the inflorescence (Prenner 2014). For what is known about the molecular control of tendril development, see Sousa-Baena et al. (2018a, b). For what is known about the molecular control of tendril development, see e.g. Scorza et al. (2017) and Sousa-Baena et al. (2018a, b). The former group noted that PeAP1 in P. edulis was expressed in flowers and the tendril, and they thought the tendril was the primary axis of the inflorescence; indeed, in taxa like P. tetrandra and P. aurantioides (subgenus Tetrapathea) the tendril is replaced by a flower (Scorza et al. 2017).
Hearn (2006, 2009a) discussed the considerable anatomical variation in Adenia as well as variation in life form. Many species have more or less grotesquely swollen stem bases, there are vines, lianes, erect herbs, shrubs and so on, and most succulent taxa have a long-lived photosynthetic epidermis and hypodermis. Hearn (2006) estimated that succulent stems evolved ca 4 times and tubers ca 8 times in the genus. Leaf morphology also varies considerably here. Adenia globosa has minute leaves and green stems (as do some other species), also stout thorns,which are modified tendrils (de Wilde & Eggli 2023). S.-Z. Yang et al. (2025) looked at the stem anatomy of some species of Passiflora from Taiwan, and as might be expected from climbing plants, tissue anatomy showed extensive variation.
Genes & Genomes. Passiflora has a genome duplication (Cai et al. 2017/18) - Malesherbia sister in this study, lacked this duplication; a duplication (?the same) of the [Turneroideae + Passifloroideae] clade, the PAEDα event, has been dated to ca 55.8 Ma (Landis et al. 2018). Given the sampling, these two duplications could be the same event. Sader et al. (2019 and references) looked at the evolution of chromosome numbers and genome size - ca (0.21-)0.41-1.31(-2.68) pg - in Passiflora, and they suggested that its base number was x = 6. Hansen et al. (2006) also discussed chromosome number evolution in Passiflora, and they thought that x = 12 might be the basal number; see also de Melo and Guerra (2003) and Mayrose et al. (2010). There is ten-fold variation in genome size in Passiflora, but this seems to have nothing to do with polyploidy, rather, two families of long terminal repeat retrotransposons accumulated in P. quadrangularis in particular, which has the largest genome, while P. organensis, with the smallest genome, nevertheless has by far the most satellite DNA (Sader et al. 2021). There seemed to be active elimation of these repeats, but not sufficient to counteract their accumulation (c.f. Liliaceae-Fritillaria).
Species of Turnera have paternal (predominant) or biparental transmission of plastids (Shore et al. 1994; Shore & Triassi 1998). Plastid transmission in Passiflora is also complex, where biparental transmission is especially evident in crosses between, but not within, species (Hansen et al. 2007). Incompatability between chloroplasts from one parent and the hybrid genome (plastome-genome incompatability - PGI) in Passiflora may result in the death of those chloroplasts and thus to variegation (Ruhlman & Jansen 2018 and references). Shrestha et al. (2021) found that in subgenus Decaloba plastid inheritance was predominantly maternal and biparental, while in subgenera Astrophea and Passiflora it was predominantly paternal, some maternal (species crosses in subgenus Deidamioides failed). For heteroplasmy here and in general, see Zhang and Sodmergen (2010), Ramsey and Mandel (2019), etc..
There have been very substantial rearrangements of the plastome in Passiflora, with both expansions, contractions and even loss of the inverted repeat (I.R.), inversions (some homoplasious; also in Adenia) and gene loss as well as movement to the nucleus; the plastomes of Adenia, Dilkea and Mitostemma (and Populus) were "normal" from this point of view (Rabah et al. 2018; Shrestha et al. 2019, 2020; Cauz-Santos et al. 2020). There are extensive parallelisms with goings on in the plastomes of Geraniaceae and Campanulaceae (Shrestha et al. 2019; Cauz-Santos et al. 2020). Recent work records the loss of the I.R. in two species of subgenus Decaloba section Xerogona, species that also had the smallest chloroplast genomes of those sampled (Cauz-Santos et al. 2020). Gene loss includes that of the ribosomal subunit genes rps7 and rpl20, as of vii.2019 not known to have been lost from any other photosynthetic angiosperms. rpl 20 has been replaced by a nuclear gene, rpl 22 has moved to the nucleus, while ycf1 and ycf2 have independently been replaced by nuclear genes in two subgenera of Passiflora (Shrestha et al. 2020; Cauz-Santos et al. 2020). The PEP (plastid-encoded RNA polymerase) subunit α is encoded by the rpoA gene in Passiflora subgenus Decaloba (Blazier et al. 2016a); highly divergent when compared with that of other angiosperms, it is probably still functional. For more on the passiflora genome, see Pacheco et al. (2020a).
Chemistry, Morphology, etc.. Cyanogenic glycosides in this family are diverse and have a variety of precursors, both protein and non-protein amino acids (R. E. Miller et al. 2006 for references); see also Spencer and Seigler (1985a: Malesherbioideae), Spencer et al. (1985: Turneroideae) and Spencer and Seigler (1984, 1987: Passifloreae).
Do the sieve tubes have non-dispersive protein bodies? For anatomical features of the family, see also Maas et al. (2019). Cronquist (1981) suggested that Malesherbioideae lacked stipules. For the floral and extrafloral nectaries of Passifloraceae, see Krosnick et al. (2008a, b, 2011). The latter are anatomically quite different from the former (i.a. they lack nectarostomata) and the CRABS CLAW gene is not expressed in them (Krosnick et al. 2008a), and so they are arguably not "homologous".
In Passiflora and its immediate relatives stem collenchyma, cymose inflorescences or branches may develop from the accessory (superposed) axillary bud. There is some debate as to whether the tendril is the pedicel of a terminal flower or the terminus of the inflorescence (Prenner 2014). For what is known about the molecular control of tendril development, see e.g. Scorza et al. (2017) and Sousa-Baena et al. (2018a, b). The former group noted that PeAP1 in P. edulis was expressed in flowers and the tendril, and they thought the tendril was the primary axis of the inflorescence; indeed, in taxa like P. tetrandra and P. aurantioides (subgenus Tetrapathea) the tendril is replaced by a flower (Scorza et al. 2017).
For a survey of floral morphology in Turneroideae, see Arbo et al. (2015). Bull-Hereñu and Ronse de Craene (2020) looked at floral development in several Malesherbioideae, and they noted that the calyx was early quincuncial, furthermore, corolla, etc., size was inversely correlated with the rate of growth of the flower. Monosymmetric flowers in which stamens, etc., are adaxially positioned are found in species of Passiflora like P. ampullacea, and at least sometimes here the odd sepal is abaxial (Macdougal 1994: there are also four carpels); P. unipetala has but a single petal in the adaxial position. Although the tubular flowers of Passifloraceae s.l. are often described as having a hypanthium, the floral tube is nearly always formed from calycine and corolline elements only (Puri 1948 and references). For general floral morphology in Passiflora see Endress (1994b) and for that in subgenus Decaloba, see Krosnick et al. (2006).
There has been work on the development of the complex series of fimbriae and membranes at the base of the androgynophore and in the throat of the KC tube in Passiflora (see e.g. Vanderplank 2000 for terms used to describe them). In subgenus Passiflora at least, genes normally involved in stamen development are expressed in the centripetally developing fimbriae/corona, some considering these to be staminodial, although the limen, a rim structure at the base of the androgynophore protecting the nectar, is from this point of view an organ sui generis (Hemingway et al. 2011). These fimbriae – there are up to five series or so - are also unique (e.g. Prenner 2014: P. lobata; Scorza et al. 2017), Claßen-Bockhoff and Meyer (2015) noting that they are initiated as the receptacular meristem expands, and this is well after all the main floral organs have been initiated. These fimbriae are unlikely to be staminodia (Prenner 2014: P. lobata), certainly, the five stamens themselves come from the androgynophore, a part of the flower quite separate from that where the fimbriae are borne. Claßen-Bockhoff and Meyer (2016) also discuss the development of the corona, and Bernhard (1999a) looks at similar structures here and in other Passifloreae. Although genes involved in staminal development have also been coopted into the development of the fimbriae/corona, as with the corona and related structures in Amaryllidaceae, strict homology cannot be invoked. For more on floral anatomy of Passiflora, see Puri (1947).
Passiflora has pollen that is 6- or more colporate (subgenus Tetrapathea unknown) and the colpi are at least sometimes in pairs, six pairs being quite common in subgenus Decaloba; pollen can germinate prematurely in the anthers forming a mass of pollen grains - "sticky pollen" (Vanderplank & Vanderplank 2022). For pollen in Brazilian species of Turnera, rather similar to that of Malesherbioideae, see De Freitas Cruz et al. (2025).
Adenia seems rather different from other Passifloroideae, perhaps being phenetically more like the other subfamilies and in having an only moderately developed corona and tricolporate pollen (e.g. see Feuillet & MacDougal 2006). Adenia also has a nectary often made up of separate glands, a hollow style, and its stigma lacks multicellular papillae (Bernhard 1999a, c), in addition, it may be dioecious, it lacks an androgynophore but may have a gynophore, its stamens are sometimes connate, and some species have a true hypanthium (de Wilde 1971b).
The styles of Malesherbioideae are shown as being commissural by Schnizlein (1843-1870: fam. 198). Ricardo S. (1967: c.f. p. 24 & Fig. 5) described the micropyle as being endostomal, but it is clearly illustrated as being bistomal; the structure of the seed coat in the micropylar region is odd. The palisade layer in the seed coat of Passifloroideae is described as being "Exotegmic with prismatic cell" by Martínez (2017: Table 3), Turneroideae have a question mark, and Kramerioideae are not mentioned; the ruminations/alveoli in the endosperm of Passifloroideae are caused by inpushings of that layer or cells. For a discussion on aril development, see Kloos and Bouman (1980); although the aril is often described as being funicular, they incline to call it raphal.
General information on Malesherbioideae is taken from Ricardo S. (1967) and Kubitzki (2006b); for chemistry, see Hegnauer (1969, 1990).
Embryologically Malesherbioideae are largely unknown.
For general information on Passifloroideae, see Harms (1925), de Wilde (1971b, 1974), Feuillet and MacDougal (2006), also de Vos and Breteler (2009: Paropsieae) and Vanderplank (2000) and Ulmer and MacDougal (2004), both Passiflora, for chemistry, see Hegnauer (1969, 1990), for anatomy, see Harms (1893) and Ayensu and Stern (1964), for branching and growth, see de Wilde (1971a) and Cremers (1974), for stipules, see Dahlgren and van Wyk (1988), for floral morphology, see Bernhard (1999: Passifloreae), for pollen morphology, variable and rather confusing, see Presting (1965), Spirlet (1965), Mezzonato-Pires et al. (2017: Passiflora, 2019: Adenia, 2022: Paropsieae, extensive variation), Soares et al. (2019), Araúo et al. (2019) and Richardo and Silvério (2019), for carpel orientation of Passifloreae, see Le Maout and Decaisne (1868) and Schnizlein (1843-1870: fam. 197), for embryology, etc., see Raju (1956a) and Singh (1970), for seed morphology in Passiflora, see Hermsen (2023) and for arils, see e.g. Pfeiffer (1891) and Kapil et al. (1980).
For the rather little that is known about Pibirioideae, see Maas et al. (2019).
For information on Turneroideae, see Berger (1919) and Arbo (2006), both general, Arbo (2008: revision of Turnera) and Gonzalez et al. (2012: Adenoa, general); see also Hegnauer (chemistry), González and Arbo (2005: anatomy), Raju (1956b), Vijayaraghavan and Kaur (1967) and Gonzalez and Arbo (2013), all embryology and seed, and Arbo et al. (2015: esp. seeds, diversity of arils).
Phylogeny. Turneraceae were weakly associated with Malesherbiaceae in Chase et al. (2002), the two being strongly associated with Passifloraceae; the same groupings were found by Hearn (2006). Korotkova et al. (2009: only three taxa from the three families) found that Turnera and Passiflora were sister and with 98% jacknife support. Preliminary data seemed to suggest that a paraphyletic Passifloraceae might include Turneraceae and Malesherbiaceae (A.P.G. II 2003), but Tokuoka (2012), Xi et al. (2012b), M. Sun et al. (2016), etc., obtain the basic relationships shown above. The recently-described Pibiria was found to be sister to Turneroideae, and with good support (Maas et al. 2019), but Adenoa, sister to all other Turneroideae, was not included. Partly because of this Rocha et al. (2019) seemed not too happy and included Pibiria in their key to New World Turneroideae although it was not in their molecular analyses.
Thulin et al. (2012b) disentangled relationships within Turneroideae, finding a largely Old World and a largely New World clade (see above). Relationships are [[Adenoa [Piriquetia + Turnera]] [Erblichia [African and Madagascan clade]]] (see also Tokuoka 2012; Sun et al. 2016: Stapfiella a little migratory). Rocha et al. (2019) focussed on the Old World taxa, but retreived the same basic relationships just mentioned, but within the Old World clade there was weak support for the paraphyly of Turnera; morphologically-based sections (e.g. Arbo 2008 and references) not faring too well, only 5/11 turning out to be monophyletic. Arbo and Espert (2009) had carried out a morphological phylogenetic analysis of Turnera that resulted in a basally pectinate tree with little support; relationships suggested by a later study with 91 characters, including several from the seed, were rather different from those apparent in molecular analyses (Arbo et al. 2015; c.f. e.g. Rocha et al. 2019, 2020).
For relationships within Malesherbioideae, see Gengler-Novak (2002, 2003).
Tokuoka (2012) clarified the phylogeny of Passifloroideae, only Paropsiopsis not being included in the study. Paropsieae are monophyletic, with [Paropsia + Viridivia] being sister to the rest (see also M. Sun et al. 2016). Within Passifloreae relationships were [Adenia [[Dilkea, Passiflora, etc.] [Basananthe, Deidamia, etc.]]] (Tokuoka 2012; see also Sun et al. 2016). In earlier studies, e.g. Krosnick and Freudenstein (2005) and Hearn (2006), the position of Adenia was unclear. Hearn (2006) provided a phylogeny for the genus, the basal branches having a fair amount of support.
The phylogeny of Passiflora is beginning to be disentangled, see Yockteng and Nadot (2004), Krosnick and Freudenstein (2005: also morphology, 2006) and Muschner et al. (2012: outline only). Hansen et al. (2006, 2007) noted problems faced in phylogeny reconstruction when using data from the chloroplast genome (see Genes & Genomes above). Shrestha et al. (2019) looking at plastomes found that subgenera Deidamioides and Tetrapathea were polyphyletic, while Cauz-Santos et al. (2020: 68 chloroplast protein-coding genes, 46 Passiflora species) confirmed the polyphyly of the former, but only one species of the latter was included. Krosnick et al. (2013: much information) discuss the phylogeny of Passiflora subgenus Decaloba. Acha et al. (2021: 91 species, 2b-RAD sequencing) looked at relationships within sect. Decaloba, which divided into the Central and South American clades; species limits in the latter were somewhat unsatisfactory.
Classification. Including Turneraceae and Malesherbiaceae in Passifloraceae s.l. was an optional arrangement in A.P.G. II (2003), and because of the basic similarity of the three families, they were combined in A.P.G. III (2009).
For genera in Turneroideae, see Thulin et al. (2012b) and Rocha et al. (2019), while Rocha et al. (2020) provide an infrageneric classification of Turnera with 11 series, and also subseries. The scetional classification of Adenia is not supported (Hearn 2006).
Passiflora includes Hollrungia and Tetrapathea (Krosnick & Freudenstein 2006; Krosnick et al. 2009). For a formal infrageneric classification of Passiflora, see Feuillet and Macdougal (2004) and Krosnick et al. (2009, 2013); there are five subgenera (at least one is polyphyletic), but the vast majority of the species are in subgenera Decaloba and Passiflora (200< species each) and Astraphea (ca 60 spp), while the other three subgenera/clades have at most 7 species. There is a complex classification of subgenus Decaloba summarized by Hermsen (2023).
Thanks. I am grateful to J. M. Macdougal for information.
[Lacistemataceae + Salicaceae]: C 0; anthers ellipsoid to subglobose; endosperm copious.
Evolution: Divergence & Distribution. Optimizing perianth characters on the tree is difficult around here.
Age. This node has been dated to (60-)57, 53(-50) Ma (Wikström et al. 2001), ca 91.1 Ma (Tank et al. 2015: table S2), (108-)100(-96)/(96-)90(-89) Ma (Davis et al. 2005a), (81-)73, 72(-64) Ma (Bell et al. 2010), (94.2-)87.1(-80.5) Ma (Xi et al. 2012b: table S7) and (73-)62(-53) Ma (Percy et al. 2014).
LACISTEMATACEAE Martius, nom. cons. - Back to Malpighiales
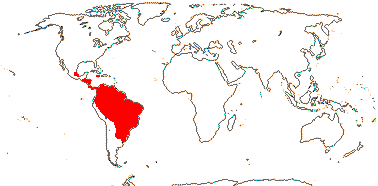
Trees; plants Al accumulators; chemistry?; vessel elements with scalariform perforation plates alone; sieve tubes?; petiole bundle D or deeply C-shaped, also wing bundles +; leaves two-ranked, (lamina entire); inflorescence raceme-like to densely spicate, (flowers 3/node); flowers small [<4 mm across]; P +, uniseriate, cup-like, (1-)4(-6); A 1, thecae ± separated (stipitate); (G [2]), median member adaxial, style branches short, ?stigma; ovules 1-2/carpel, apical, funicles thick, long, ovule type?; fruit a 1(-3)-seeded capsule; ?aril; testa fleshy or not; embryo (short), with foliaceous cotyledons; n = 22, ?31, chromosomes 0.9-2.3 µm long.
2 [list]/17: Lacistema (12). Greater Antilles (Jamaica), Mexico southwards, not in Chile. Map: from Sleumer (1980).[Photo - Flower, Fruit]
Age. Crown-group Lacistemataceae are around (49.3-)19.8(-2.6) Ma (Xi et al. 2012b: table S7, inc. Los.).
Chemistry, Morphology, etc.. The inflorescence may be derived from a more elborate form with determinate branches. Chirtoiü (1918) described the flower as having 4-5 free perianth parts and an irregularly lobed, annular nectary. The presence of an aril in Lacistemataceae needs to be confirmed (see also Corner 1976). Sleumer (1980) records an aril in Lacistema, but a fleshy seed coat for Lozania. In Lozania there are long "hairs" inside the fruit which perhaps support the dangling seed; these hairs are thick-walled but unlignified cells that may be derived from the funicle (see also Casearia, Salicaceae).
Additional information is taken from Sleumer (1980: as Flacourtiaceae-Lacistemeae); see also Lozania - Riviere 270 (anatomy), Gentry et al. 22231 (fruit); Lacistema - Aymard & Delgado 6882 (fruit), Rimachi Y. 11201 (anatomy - stomata tending to anisocytic). See Young (2008 onwards: Lacistemataceae website, focus is on species, nomenclature, etc..
The embryology, etc., of the family remain largely unknown.
Phylogeny. Lacistemataceae did not cluster with the rest of Salicaceae and Kiggelariaceae in an early study by Savolainen et al. (2000a), although they were probably in this area (Chase et al. 2002; see also D. Soltis et al. 1999, 2000). Davis et al. (2005a) place them as sister to Salicaceae s.l. (61% bootstrap, 1.0 posterior probability), as do Korotkova et al. (2009: slightly higher jacknife); as might be expected, they lack salicoid leaf teeth.
SALICACEAE Mirbel, nom. cons. - Back to Malpighiales
Shrubs to trees; cocarcinogens, (cyclopentenoid cyanogenic glycosides and/or cyclopentenyl fatty acids [gynocardin]), (ellagic acid) +, tanniniferous; tension wood fibre walls multilayered [1-5 layers]; cork?; stomata brachyparacytic; leaves two-ranked, lamina vernation supervolute-curved or involute, (margin entire), (glands +), (stipules 0); inflorescence various; flowers 3-6-merous, (hypanthium +); P = K or T; nectary often lobed; anthers (extrorse), (linear); G [2-5], styles separate or fused; ovules anatropous, micropyle usu. bistomal and ± zig-zag, (embryo sac elongated, ± protruding into the micropyle), (embryo chlorophyllous); nuclear genome [1 C] 0.36-0.86 pg.
54 [list: to tribes]/1200. Pantropical, also temperate (but few in the Antipodes) to Arctic. Map: from Sleumer (1954), Meusel et al. (1975), Sleumer (1980), Hultén and Fries (1986) and Trop. Afr. Fl. Pl. Ecol. Distr. Vol. 1 (2003). Photo: Flower, Fruit.
Age. Crown-group Salicaceae have been dated to (50-)47, 40(-37) Ma (Wikström et al. 2001), (71-)63, 61(-55) Ma (Bell et al. 2010), (87-)79.2(-72.8) Ma (Xi et al. 2012b: table S7), 102.1-86.0 Ma (de Mestier et al. 2022), (79.4-)55.3(-33.2) Ma (K. Yan et al. 2025), (136.3-)120.9(-103.3) Ma (Ogutcen et al. 2024) ca 98.9 Ma (L. Cai et al. 2025b).
López-Martínez et al. (2023a: ) noted that Mabelia connectifila, from Turonian deposits in New Jersey, U.S.A., although initially identified as Triuridaceae, was often linked with Salicaceae in their analyses...
1. Samydoideae Reveal —— Synonymy: Bembiciaceae R. C. Keating & Takhtajan, Prockiaceae Bertuch, Samydaceae Ventenat, nom. cons.
(Heartwood brown, rays wide, visible - Irenodendron); petiole bundle arcuate; colleters +; lamina usu. punctate or lineate, teeth theoid [glandular portion ≡ a colleter, water pore +, but no epithem], vernation involute; (inflorescence fasciculate); hypanthium +; P uniseriate, 3-7, basally connate/connate; nectary on base of P/hypanthium; A 3-many, spiral, (uniseriate, initiated simultaneously; staminodes often + - Casearia), (filaments closely adpressed, forming a tube); tapetal cells 2-4-nucleate; styluli +/style +, apically branched, (stigmas bifid); embryo sac straight, outer integument ca 2 cells across, inner integument ca 2 cells across, hypostase + [C.]; (aril as tuft of hairs - some C.), (seed squeezed from fruit, aril vascularized - C.); exotegmen cells laterally flattened, crystalliferous; n = 21, 22; nuclear genome [1 C] 0.68-0.71 pg.
11/235: Casearia (200). Pantropical, especially South America.
Age. Crown-group Samydoideae (Lun-Cas) are (38.8-)37.4(-36.3) Ma (Xi et al. 2012b: table S7) or (Tet-Cas) (95.0-)89.7(-64.9) Ma (de Mestier et al. 2022).
[Scyphostegioideae + Salicoideae]: lamina teeth salicoid [a vein or veins proceeding into the tooth, where they expand/branch, tooth apex a variously coloured spherical gland or stout hair].
Age. The age of this node is (78.7-)68.9(-59.8) Ma (Xi et al. 2012b: table S7) or (58.4-)39.7(-22.3) Ma (K. Yan et al. 2025).
2. Scyphostegioideae Reveal
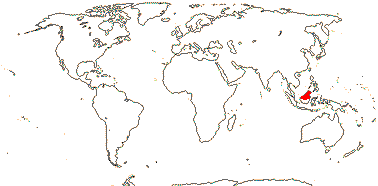
Plant dioecious; flowers ± 3-merous; P ?biseriate, petal-like, connate; staminate flowers: A connate, extrorse; carpelate flowers: placentation basal, stigmas prominent, spreading; ?embryo sac, funicle +; fruit sulcate; aril +; seeds hairy.
2/2: Borneo, S.W. China.
2A. Scyphostegia borneensis Stapf —— Synonymy: Scyphostegiaceae Hutchinson, nom. cons.
Vessels usu. in radial multiples; rays mostly uniseriate; cork superficial; petiole bundle cylindrical, with adaxial mass of xylem and phloem, changing to deeply U-shaped, with adaxially-flattened cylinder - xylem external - between arms; inflorescences terminal, branched, long-lived, ?racemose, bracts large, overlapping, tubular, pedicels not articulated; staminate flowers: nectary as lobes opposite A; A 3, opposite inner T, filaments as long as anthers; pollen ?tricolpate, ?surface; carpelate flowers: G [8-13], style 0, stigmas ray-like, with an opening in the middle; ovules with much elongated micropyle, ?exostomal, outer integument 2-3 cells across, lobed, inner integument 3-4 cells across, nucellar cap +, persistent; fruit a fleshy capsule; seeds with aril from funicle/outer integument; endosperm slight, perisperm +, very scanty; n = 9; nuclear genome ?.
1/1. Borneo, not the southern part. [Photo - Flower, Leaf, Inflorescence.]
2B. Dianyuea turbinata C. Shang, S. Liao & Z. X. Zhang
Plant deciduous; peduncular thorns +; ?stomata; ?petiole bundle; lamina pli-nerved; inflorescences axillary; bracts linear; staminate inflorescence cymose, staminate flowers: sessile; P 6-8; A 6(-8); nectary 0; filaments much shorter than anthers; pollen surface striate; carpelate flowers: single; P 8-11; G [4-6], stigmas bilobed; pericarp thick; seeds long-arillate; n = 19/18 + 2 Bs; nuclear genome [1 C] ca 4.32 pg.
1/1. Yunnan, China.
3. Salicoideae Arnott
Benzoylated glycosides, etc. +; petiole bundle arcuate or annular with wing bundles; inflorescence terminal or axillary; A many, development centrifugal, dehiscence extrorse; G [2-5(-13)], (stigmas flabellately divided); (ovule straight), outer integument 2-5 cells across, inner integument 3-5 cells across, (nucellar cap +), (hypostase +), funicle (long), (0); embryo sac (bisporic [chalazal dyad], eight-celled [Allium-type]); n = 9-11(-12).
40/961. Worldwide, but few Australia, none New Zealand.
Age. The age of crown Salicoideae, or somewhere around there, is (36-)33, 26(-23) Ma (Wikström et al. 2001), (55-)51, 50(-47) Ma (Bell et al. 2010), (65.1-)58(-51.3) Ma (Xi et al. 2012b: table S7 - see below), (58-)52(-48) Ma (Percy et al. 2014), and ca 61 Ma (M.-M. Li et al. 2019).
N.B. - the tribes below are rather notional.
3A. Homalieae (R. Brown) Dumortier / here to Abatieae = Saliceae A - n = 9, 11, nuclear genome [1 C] 0.40-0.42 pg —— Synonymy: Homaliaceae R. Brown
Flowers 4-8(-many) merous; P = ± T, outer whorl valvate to ± imbricate, adaxially with a large disciform nectary gland, stalked or not; A = or in fascicles to 15 opposite inner whorl of T; (G ± inferior); fruit loculi hairy inside/not; n = 10, 11, nuclear genome [1 C] 0.39-0.42 pg.
9/200: Homalium (150-180). Pantropical, esp. America and Africa.
3B. Bembicieae Warburg - Bembicia capitata Oliver
Inflorescence ± capitate, pedicels 0; P = K + C; C valvate; ?A; G semi-inferior.
1/1-2. Madagascar.
3C. Prockieae Endlicher
(Cyanogenic glycosides - Banara); petiole bundle arcuate/annular; lamina venation palmate; P = K + C, K ± valvate, (C 0); disc glands often 0; A many; placentation also axile.
8/45: Banara (31). Tropical America.
3D. Abatieae Bentham & J. D. Hooker
Leaves opposite; inflorescence terminal; hypanthium ± +; P uniseriate, valvate; A 4-many; n = 36.
2/10. Tropical montane Central and South America; south Brazil.
3E. Scolopieae Warburg / Saliceae B
Infloresence various, (epiphyllous); (P = K + C); A often many, centrifugal, (introrse - Mocquerysia); funicle 0; embryo sac enclosed by nucellus; n = 9, 11, nuclear genome [1 C] 0.32(-0.39) pg.
5/45: Scolopia (37). Tropical Africa, Malesia.
3F. Saliceae Reichenbach / Saliceae C —— Synonymy: Flacourtiaceae Richard, Poliothyrsidaceae Doweld
(Plant deciduous); (tension wood fibre walls with single thick [= G-] layer, p-hydroxybenzoate moieties on lignins of Salix and Populus) ; (nodes 2:2 - some Azara); leaves often spiral, (nectaries + - position various), lamina (venation palmate); plant (dioecious); (inflorescence catkinate); P 0/uniseriate/(biseriate), (valvate); A 4-many, (dehiscence part introrse - Idesia); nectary 0/+, position various - see Chemistry, Morphology, etc. below); (pollen inaperturate - Populus); G [2-10]; micropyle exostomal - Idesia, (integument 1, 3-4 layers across, dense multicellular intraovarian hairs + - S./inner integument suppressed - Populus), (funicle 0); antipodal cells often large; fruit baccate/drupaceous/capsular; seeds (winged/with funicular-hilar arillate hairs - S.); (seed coat thin, exotesta alone - S.); (endosperm 0, embryo chlorophyllous - S.); n = 19-22(...); nuclear genome [1 C] (342-)565(-841) Mb, 0.47-1.30(-3.3 - Bennettiodendron) pg.
16/840: Salix (629), Xylosma (85), Populus (35), Flacourtia (15). Worldwide, but not south temperate.
Age. The Azara-Salix clade is ca 62.5 Ma, the Saliceae Clade C are 51-45 Ma (Sanderson et al. 2024), (35.9-)23.3(-12.4) Ma (K. Yan et al. 2025) is the age of a clade including Bennettiodendron, Abatia, etc., and (93.5-)80.3(-68.5) Ma the age of Saliceae s. str. (Ogutcen et al. 2024).
Populus leaves are reported from deposits ca 59 Ma and leaves and fruits from deposits ca 48 Ma in the western U.S.A. (Wing 1987; Manchester et al. 2006; Vanneste et al. 2014a), which leads to questions about some of the ages above.
Evolution: Divergence & Distribution. Boucher et al. (2003) described Pseudosalix, a fossil found in early Eocene to early Oligocene deposits 50-32 Ma (Manchester et al. 2006) from North America. It is morphologically intermediate between Salix and more florally conventional Salicaceae, having a terminal, branched inflorescence and flowers with well developed sepals. Some estimates suggest that the [Salix + Populus) clade is around 34 Ma (see Brundrett 2017a; Tedersoo 2017b; Tedersoo & Brundrett 2017; M.-M. Li et al. 2019) or ca 48.2 Ma (L. Cai et al. 2025b), although such ages are very much at odds with some of those of the fossils mentioned above. On the other hand, Fig. 4 in L. He et al. (2020) suggests that the ancestor of S. triandra diverged from the rest of the genus some time towards 50 Ma while K.-Y. Chen et al. (2024) estimated that the age of crown-group Salix was (50.8-)46.3(-31.8) Ma.
De Mestier et al. (2022) suggest a number of other ages in Casearia-Samydeae; see also K. Yan et al. (2025). Diversification in Scyphostegioideae seems to have slowed down (!: Xi et al. 2012b: as Scyphostegiaceae, one species then, two now).
Casearia may have originated in South America, and there have been a number of moves on to the Antillean islands and one to the Old World between (31.0-)25(-18.0) Ma [stem]/ca 19.8 Ma [crown ages] (De Mestier et al. 2022). The geography of relationships in the Old World, [New Caledonia [Africa + Asia]], is interesting, although note that the focus of the work by De Mestier et al. (2022) was on New World taxa.
As mentioned below, the large genus Salix is a major element of the vegetation in Arctic regions in particular and elsewhere in the North Temperate zone. It is notably diverse in the Hengduan Mountain region of the Qinghai-Tibet plateau, and = there has been increased diversification in the two main clades in the genus (subgenus Salix and the rest) in response to Oligocene-Miocene tectonic and climate change events (K.-Y. Chen et al. 2024). Salix is notorious for its extensive interspecific hybridization and for how difficult it is to identify the species - flowers and leaves are on the plant at different times, etc.. Hybridization between different clades like the Salix and Vetrix clades is not so common, perhaps because of their long sex-linked regions and heteromorphic chromosomes (Z.-Q. Xue et al. 2024). Indeed, the sex expression system in Salix (see Genes & Genomes below) may be involved in the prevention of introgression between the species. Gulyaev et al. (2022) thought about chromosomal rearrangements from this point of view, but only two sex chromosome systems were known then. Xue et al. (2024) found that introgression involved the autosomes, but less the sex-linked regions which were functioning as a sort of supergene; these regions occupied somewhat under 25% of the sex chromosome. There was frequent introgression involving the autosomes when members of the three chromosomal types (15XY, 15ZW and 7XY) crossed, less in the sex-linked regions, and there was no gene flow when 15ZW and 7XY species crossed. A burst of diversification in the Vetrix clade of Salix is apparently linked to the adoption of the 15ZW system there is associated with a major clade from the Hengduan Mountains (15XY⇒15ZW: He et al. 2020; Sanderson et al. 2023; Xue et al. 2024). Diversity within the Chamaetia-Vetrix clade in the Hengduan Mountain region and environs may be the result of range changes over the last glaciation in particular, dispersal and isolation in this mountainous area, and adaptations along altitudinal gradients (He et al. 2020). Marincek et al. (2024, see also N. D. Wagner et al. 2023) found that there was a North American Clade in the shrub willows (there is also a North American clade in Populus) and an European clade which included some North American representatives. Diversification in Salix is associated with widespread polyploidy, some 40% of the species being polyploids, and, as mentioned, hybridization. This latter is discussed by Gambhir et al. (2025) who focused on cases of serial chloroplast capture and subsequent speciation. Within Populus the closely-related P. balsamifera and P. trichocarpa introgress at contact zones and are estimated to be ca 75,000 y.o. using nuclear data - and 6-15 m.y.o. using plastome data - perhaps chloroplast capture from a ghost lineage (Huang et al. 2014)? M. Wang et al. (2019) had found a evidence of ancient gene flow within and between the major clades they recovered in Populus, and there were also a number of ancient transpecific polymorphisms maintained by balancing selection. Hybridization, etc., occurs in both genera, perhaps especially in Salix (Z.-S. Zhang et al. 2021), and they are examples of syngameons (see also Quercus for another example).
Ecology & Physiology. Salix and Populus are often ectomycorrhizal (ECM) plants (Tedersoo 2017b; Tedersoo & Brundrett 2017) and grow with other ECM trees in boreal forests; they represent an early successional stage. In Alaska the leaves of Populus balsamifera, for example, are rich in condensed tannins, and pentamers (and above) of these tannins form complexes with nitrogenous compounds, the latter already in low concentrations, in the humic layers of the soil, and this makes N unavailable for many plants (Fierer et al. 2001); such tannins may be up to 25% dry weight, but any implications of this for plant defence are unclear (Barbehenn & Constabel 2011). (Note that the ECM white spruce, Picea glauca, dominated in the next stage of the succession - Fierer et al. 2001.) M.-H. Li et al. (2022) note that P. trichocarpa uses the bisaccharide (two glucose molecules) carbohydrate trehalose from its fungal associates, the plant having multiple copies of the trehalase gene that can break trehalose down. Salix itself is a major component of the biomass in some tundra ecosystems (Chapin & Körner 1995), and with almost 70 species growing in the Arctic, it is the second largest genus there (Elven et al. 2011).
Like some other woody pioneers in which quick seedling establishment is at a premium, Salix and Populus (?all species) germinate very fast, i.e. they germinate less than one day after the seeds starts imbibing water, and some are pioneer plants in the same habitats as Tamarix, seeds of which also have very fast germination (Parsons 2012; Parsons et al. 2014).
A single clone of Populus tremuloides is recorded as covering 43.6 ha and has around 47,000 stems (DeWoody et al. 2008), numerous vegetative buds developing from the roots (references in Bosela & Ewers 1997). Ally et al. (2010) found that after 500 years or so the clones showed signs of aging, at least in terms of the amount and viability of pollen, and the fertility of the pollen of the oldest clone they looked at, perhaps 10,000 years old, was less than one quarter than that of young plants. A recent estimate of the age of this clone is 116-18,000 years old (Pineau et al. 2024; c.f. Ginkgo, see also age and plants elsewhere).
Boeckler et al. (2011) discuss the anti-herbivore properties of phenolic glycosides, the salicylates (salicylic acid seems not to be an intermediate in their synthesis), that are characteristic of Salix and its immediate relatives, sometimes being up to 30% of the dry weight of the plant although not present in all taxa (see also Pentzold et al. 2014 and references). They act variously as toxins and deterrents against feeding by generalist herbivores , but as with other defences against generalist herbivores a number of specialist insects and fungi have circumvented them (Boeckler et al. 2011; Volf et al. 2015; Julkunen-Tiitto & Virjamo 2017). Also, as in a number of similar systems, closely related species of Salix have rather different kinds of defences (Volf et al. 2015). p-hydroxybenzoate units decorate the lignins in fibres of both Salix and Populus. although their function is unclear (Goacher et al. 2021).
Metal hyperaccumulators (nickel) are notably common in members of the family growing in New Caledonia (Brooks 1998 for a summary; Gei et al. 2020).
Pollination Biology & Seed Dispersal. Populus is dioecious and wind-pollinated. In the northeast U.S.A., at least, oligolectic bees are, perhaps rather surprisingly, notable visitors to Salix, also dioecious (Fowler 2016).
Plant-Animal Interactions. Populus (#5 of the top 29 genera) and Salix (#2) are very important host plants for lepidopteran larvae in the contiguous United States of America (Narango et al. 2020). Furthermore, around 34 species of Phyllonorycter leaf-mining moths (Lepidoptera-Gracillariidae-Phyllocnistinae) are found on the same genera in the Holarctic region (Lopez Vaamonde et al. 2006). Caterpillars of Atella (Nymphalinae) fed on the old Flacourtiaceae and Salicaceae (Ehrlich & Raven 1964) as do several Vagrantini (Nylin et al. 2014), while some Notodontidae moths (J. S. Miller 1992) show similar host patterns.
There has been a remarkable radiation of around 400-500 species of euurine sawflies (Hymenoptera-Tenthredinidae-Nematinae) - the larvae are mostly gallers but some fold or roll leaves - on Salix and Populus, although they have yet to be recorded from other Salicaceae. Leaf folding appears to be the primitive condition, and the sawflies show considerable host specificity (Roininen et al. 2005; Nyman et al. 1998, 2006; Price 2005); high diversity and high host specificity are not commonly correlated in Lepidoptera, at least (Menken et al. 2009). Unlike other galls, sawfly-induced galls result largely from stimuli provided by the ovipositing insect itself, which may inject fluids into the plant, rather than from the activities of the larvae, so the galls assume their mature forms before the eggs hatch (Redfern 2011).
Some 70 species of the gall-forming Pemphigus (Aphidae) are found on Populus, its primary host (where P. saliciradicis grows needs no explanation), sometimes on a single species only. Pemphigus spyrothecae in particular is noted for having soldiers and showing complex social behaviour (Peccoud et al. 2010; Floate 2010).
Extrafloral nectary activity in Populus may be induced by mechanical wounding, etc. (Heil 2015).
Plant-Bacterial/Fungal Associations. Salix and its immediate relatives enter into a variety of associations with both ECM and endomycorrhizal fungi, and dark septate endophytes have also been found in them (e.g. Gardes & Dahlberg 1996; Van der Heijden 2001; Becerra et al. 2009; Bennett et al. 2017: Supplementary Material). Some 179 fungal OTUs were recorded from Salix arctica alone in a Low to High Arctic transect, many of these also being widely distributed outside the Arctic (Timling & Taylor 2012), while a single individual of aspen, Populus tremula, may host as many as 122 ECM species (Bahram et al. 2011). Tedersoo et al. (2013) found a strong correlation between host and ECM community composition and species richness in Salix and Populus - and in a small sample of less closely related hosts. Wounding individuals of Populus x canescens associated with the ECM basidiomycete Laccaria bicolor caused extensive changes to the secondary metabolites produced by the plant, away from phenolic compounds to those that deterred the activities of the leaf beetle Chrysomela populi (Kaling et al. 2018).
Rusts, Melampsora spp., are found on Salix and Populus, M. idesiae on Idesia (Holm 1979; Z. S. Zhang et al. 2021). The role of salicylates in defending against rusts is unclear (Julkunen-Tiitto & Virjamo 2017).
Genes & Genomes. A genome duplication in the common ancestor of Salix and Populus, the salicoid duplication, has been dated to 65-60 Ma (Tuskan et al. 2006; see also Gao et al. 2018; Zwaenepoel & Van de Peer 2019). It will be interesting to know if Saliceae other than Salix and Populus have it, but it is not known from Abatia in Clade A of Saliceae; other members of clade C (containing Salix, etc.) have high polyploid numbers, but have very few species, and members of clades A and B have lower numbers (Sanderson et al. 2023). (Note that the literature quite often implies that Salicaceae = Salix + Populus alone.) This duplication has also been dated to (36.3-)34.7(-32.6) Ma (Vanneste et al. 2014a: they think that this is an underestimate) and ca 33.6 Ma, the SAACα event (Landis et al. 2018), while Murat et al. (2015b) dated a duplication in Populus to 17-7 Ma. However, as noted above, ages are uncertain around here; see also Cai et al. (2017/18) for problems in dating. Jiang et al. (2013) followed the fate of duplicated genes, and Garsmeur et al. (2013) and Y. Liu et al. (2019) suggested that the duplication might have been an autotetraploidy event, the two Populus genomes having remained rather similar despite the age of the event being some 60 Ma. Z.-S. Zhang et al. (2021) placed the duplication at the base of their Clade C (= [[Idesia + Bennettiodendron] [Carrieria [Poliothyrsis [Itoa [Salix + Populus]]]]], Saliceae above, all members of which are polyploids on n = 19-22 - only Abatia of the ten other Salicoideae (including Oncoba) the chromosome numbers of which are known is polyploid. Overall chromosome number changes in this part of Salicaceae are perhaps x = 12 → 24 → 19 (Populus) (Murat et al. 2015b). Populus has lost the `nuclear PHYC gene involved in responding to red-far red in light (Matthews 2010 and references); the phylogenetic extent of this loss is of interest. Z.-S. Zhang et al. (2021: note the topology of the tree that they use - Fig. 1, with Saliceae groups A, B, C) give some genome sizes, for which, see also Sanderson et al. (2023: and chromosome numbers).
Practically the whole [Salix + Populus] clade is dioecious, and Z/W (female heterogametic) and X/Y (male heterogametic) sex determination systems have evolved in both genera, a type-A cytokinin response regulator apparently being a master regulator of sex determination throughout the clade; there has been duplication, transposition and rearrangement of the gene, and the sex determination region has changed its position in the genome quite recently (W. Yang et al. 2020); in Salix there are 15XY (male heterogamety), 15ZW (female heterogamety) and 7XY systems (the number refers to the sex chromosome), the 15XY system being ancestral. At one level there is great fluidity in details of sex expression, and Y. Wang et al. (2024) describe the changes involved in the shift from an XY to a ZW system. Despite the fact that the whole clade is dioecious and perhaps 34 Ma, or even much older, changes in the control of sex expression may have occured within the last 7-6 Ma (Geraldes et al. 2015; Hou et al. 2015; Pucholt et al. 2017). Indeed, some estimates of the age of the evolution of dioecy are as much as ca 65 Ma (see above, also Filatov 2015; c.f. the Cannabis-Humulus system in Cannabaceae, q.v.. See more under Divergence & Distribution above.
An attempt at DNA barcoding failed spectacularly in Salix, and although 2-7 plastome regions were examined, only 1 of 71 species was successfully barcoded (Percy et al. 2014, q.v. for explanations). There is rather little variation in the plastome in the Salix/Populus area, just minor changes in the length of the inverted repeat and variation in simple sequence repeats that are present; the infA gene was not found in the plastome (M.-M. Li et al. 2019, see also K.-Y. Chen et al. 2024). This was confirmed by N. D. Wagner et al. (2021), who suggested that low mutation rates, reticulate evolution and homoplasy were all involved. Indeed, Populus has low mutation rates in all three genomic compartments, nuclear, plastid, and mitochondrial (Tuskan et al. 2006), and plastome variation is commonly slight in Salix and not exactly much greater in the nuclear genome, either (Wagner et al. 2021). K. Yan et al. (2025) looked at plastome variation in Casearia; the boundary of the inverted repeat moved in C. kurzii, and there were associated changes in gene content.
Chemistry, Morphology, etc.. For different kinds of salicylates, the amount of which varies considerably between species and also the age of the plant (Salix, Populus), see Julkunen-Tiitto and Virjamo (2017). Banara is the only genus of Salicaceae reported to have cyanogenic glycosides, and it is well embedded within the family (Chase et al. 2002). For the N→S decrease in the syringyl/guaiacyl ratio in wood lignin in P. trichocarpa, see W. Zhu et al. (2025); a mutation in cell wall laccase seems to be associated with this decrease, and this increases towards the south. Catechyl lignins are also found here. (Note that in Eucalyptus spp. there is no consistent geographical trend in the syringyl/guiacyl ratio - see Stackpole et al. 2011.)
The perforation plates of the tracheary elements are more or less simple and the intervascular pits are small. Variation in the anatomy of tension wood is described by Ghislain et al. (2016), and a number of taxa have tension wood fibres with multilamellate walls; the regaining of normal (for tension wood) cell wall anatomy, i.e. with a thick G layer, in a group of genera that includes Salix and Idesia may be an apomorphy for that group. Xylosma, Flacourtia, etc., have groups of large sclereids in the phloem (Zahur 1959). Both laticifers and resin ducts are recorded from Salicaceae (Prado & Demarco 2018). Xylosma and some Casearia seem to have unilacunar nodes, while leaf traces arise two internodes below the leaf they innervate in Hasseltia. Balfour and Philipson (1962) discuss the nodal anatomy of Salix, clearly a sympdial system.
Variation in lamina anatomy is quite extensive (Thadeo & Meira 2013; esp. Thadeo et al. 2014). The spherical glands/cavities/ducts found in most species of Casearia are surrounded by a single layer of epithelium (Fernandes et al. 2018); secretory canals in Salicaceae are described as forming by the coalescence of cavities of various shapes (Fernandes et al. 2018; Costa et al. 2021: see also Humiriaceae, c.f. Calophyllaceae, etc.). For colleters and other secretory structures here, colleters also being found on the stipules, see Fernandes et al. (2017) and Santos de Faria et al. (2018). Thadeo et al. (2014: table 4, esp. fig. 7c, j) note that there is cambium in the petiole of all the taxa they examined, and illustrate Casearia decandra as having an arcuate petiole bundle, but annular in a petiole with secondary vasculature. The deciduous glandular portions of the theoid leaf teeth of Casearia have been compared with colleters (Fernandes et al. 2016), and they may also have a hydathodal function (Thadeo et al. 2014). Most of the family have some form of salicoid leaf tooth; these are quite variable in morphology, but all have secretory palisade cells over parenchyma and are well supplied by vascular tissue, especially xylem (Wilkinson 2007: check). Thadeo et al. (2008; see also Thadeo & Meira 2009) discuss the similarity between salicoid leaf teeth and foliar nectaries in Salicaceae, in particular the foliar nectaries of Prockia crucis that secrete fructose, glucose, sucrose, etc.; phloem is prominent in the vascularization (Thadeo et al. 2014). Such tooth-associated nectaries are scattered in the family (Rios et al. 2020 and literature). Nectaries at the base of the lamina in two species of Populus differed markedly. In one they were persistent, with continuous nectar flow, while in another, large amounts of nectar were produced over a short time and cell death occured, but new nectaries could be produced (Escalante-Pérez et al. 2012).
Branching in Casearia is phyllanthoid, the orthotropic axes having spirally arranged and reduced leaves while the plagiotropic branches are sylleptic and have fully-expanded and two-ranked leaves. Abatia has opposite leaves with at most very small stipules and there are marginal glands at the base of the lamina. There are also taxa with pli-nerved leaf blades and foliar glands - Salicaceae are vegetatively rather heterogeneous.
For inflorescence development in genera like Idesia and in catkinate Salicaceae, see Cronk et al. (2015). The inflorescences of Scyphostegia are distinctive in being long-lived and having overlapping tubular bracts in the axils of which the flowers arise; these inflorescences are somewhat reminiscent of those of Alpinia sect. Myriocrater. The inflorescence units are described as being racemose by van Steenis (1957), but this should be confirmed.
The valvate perianth members of Abatia are basally connate and bear many filamentous processes, and the flowers lack a nectary. Nectary morphology is very variable in those Salicaceae that do have a nectary. It has been claimed that the nectary of Salix represents a perianth member, but Alford et al. (2009) suggest that it is like that of other members of the family; there are receptacular nectaries adaxial to the stamens in taxa in the clade sister to [Salix + Populus] while in Poliothyrsis the nectaries are outside the stamens on the bases of the valvate perianth members. Nemoto et al. (2025) noted that in Idesia polycarpa the outermost stamens developed very quickly and dehiscence there was introrse, not extrorse as in the other stamens, and as is common in other Saliceae; there was a nectary between the introrse and extrose stamens (and the introrse stamens had adaxial xylem, in the extrorse stamens it was abaxial...). In Bennetiodendron, too, the outer stamens were introrse and the inner stamens extrorse; here there are small glands at the bases of all the stamens; in Xylosma congesta all the stamnens are extrorse, and there is a nectary disc immediately outside/abaxial to the androecium (Nemoto et al. 2025). As ideas of phylogeny improve here, nectary and androecial development will need to be revisited (see also Bernhard & Endress 1999).
Elongated embryo sacs occur in Salicaceae and the old Flacourtiaceae (Steyn et al. 2005a), for eaxmple, the embryo sac more or less protrudes into the micropyle in Arechevaletaia (= Azara)(Maheshwari 1950). The micropyle of Scyphostegia may be exostomal (Corner 1976). Shang et al. (2017) described the fruit of Dianyuea as being indehiscent although it has seeds with very well developed arils, a rather odd combination. The exotegmen of Dovyalis consists of ribbon-like cells.
Much of the older literature for genera in Salicaceae will be found under Flacourtiaceae. See van Steenis (1957: Scyphostegiaceae), Lemke (1988), Judd (1997a), Chase et al. (2002), Alford (2005), and Alford and Dement (2015: Samydoideae), all general information, Hegnauer (1973, 1990, also 1966, 1989), Spencer and Seigler (1985b) and Chai (2009), all chemistry, Metcalfe (1956: Scyphostegia) and R. B. Miller (1975: wood), both anatomy, Bernhard and Endress (1999: androecial initiation), Gavrilova (1998: pollen), and Narayanaswami and Sawhney (1959) and Steyn et al. (2004, 2005a, b), all ovule and seed development, summary in latter paper) and van Heel (1977, 1979: testa anatomy). For Scyphostegia, see Metcalfe (1954: anatomy), van Heel (1967a: flowers and fruits) and Hutchinson (1973: different interpretations of the gynoecium).
Phylogeny. Chase et al. (2002) greatly clarified the phylogenetic situation in the old Salicaceae-Flacourtiaceae area (see also Judd 1997a; Nandi et al. 1998; T. Azuma et al. 2000; Savolainen et al. 2000a), although sampling within the tribes needed to be extended. Three main clades (as subfamilies above) were recognized; see also M. Sun et al. (2016) for relationships within the subfamilies. M.-M. Li et al. (2019) looked at variation in the plastid genomes of 24 species (19 genera) of Salicaceae, most in Salicoideae; Saliceae turned out to be very polyphyletic, but otherwise relationships were [[Saliceae] [[Abatieae + Prockieae] [Homalieae [Scolopieae + Flacourtieae]]], and support was generally very strong. Casearieae: Casearia is sister to the rest of Salicaceae, although in early studies support for this position was weak (Chase et al. 2002; Xi et al. 2012b; M.-M. Li et al. 2019; c.f. D. Soltis et al. 1999, 2000); the genus is paraphyletic (Alford 2005; Shang et al. 2017; de Mestier et al. 2022), although the paraphyly is being cleared up. De Mestier et al. (2022: 4 chloroplast genes, nuclear ITS, ca 20% species sampled) looked at relationships within Casearia with a focus on the New World taxa. There seems to be some conflict between nuclear and chloroplast genes, Trichostephanus is odd; it was placed in Trichostephaneae, although not assigned to a family, by Chase et al. (2002), however, in lacking petals and in having a disc at the base of the calyx it is like Casearia. The old Flacourtia turbinata has turned out to be sister to Scyphostegia, which makes morphological sense (Shang et al. 2017). Ogutcen et al. (2024) found that Samydoideae and Scyphostegioideae were sister to the rest of the family they examined, but other relationships there were unclear, one critical node in particular having very weak support.
In gross morphology Oncoba is remarkably like other members of the erstwhile Oncobeae, but the latter differ in chemistry (they are cyanogenic), leaf tooth type, and stamen initiation (centripetal) and are now included in Achariaceae-Lindackerieae. However, it was thought that Oncoba itself was perhaps to be assigned to Flacourtieae (= Salicaceae-Saliceae) (for seed anatomy, see van Heel 1977a), in which it was embedded - indeed it is sister to Flacourtia itself - in Chase et al. (2002), but note that Flacourtieae there are paraphyletic. Xi et al. (2012b) found the relationships [[Flacourtia + Prockia] [Poliothyrsis + Salix and relatives]], and the assignment of Oncoba with its salicoid leaf teeth - it includes just three species - to Saliceae is holding up (see also Zmartzy et al. 2025: relationships there are [Populus [Hasseltiopsis + Oncoba]]). For the relationships of Salix and its immediate relatives, see Leskinen and Alström-Rapaport (1999). Relationships there are [Microhasseltia, etc. [[Salix + Populus] [Olmediella [Bennettiodendron + Idesia]]]]; characters like hairy seeds, sepals deciduous in fruit, loss of corolla, and dioecy are apomorphies at various levels within this clade (Alford et al. 2009). X. Liu et al. (2016) recovered the quite well supported relationships [Poliothyrsis [[Bennettiodendron + Idesia] [Salix + Populus]]]. This clade was sister to a clade in which seven members of both Scolopieae and Saliceae (see above) were variously combined, but no other Salicaceae were examined. Although Salix, etc., were not examined, K. Yan et al. (2025) looked at plastome variation of some other Saliceae.
Substantial work has ben carried out in the resolution of relationships within Salix itself. For its phylogeny, see T. Azuma et al. (2000) and J.-H. Chen et al. (2010), also X. Liu et al. (2016 and references). Of course hybridization and associated conflicts between nuclear and plastome trees (e.g. Gulyaev et al. 2022; Z.-Q. Xue et al. 2024) have caused complications. Marincek et al. (2024: nuclear data, focus on North American shrub willows, see also N. D. Wagner et al. 2023) was able to obtain substantial resolution in the relationships of species there. Ogutcen et al. (2024) carried out a variety of analyses on some 47 species of Salix, and the results varied according to the data used. Sections were quite often not monophyletic, and subgenera Chamaetia and Vetrix seemed best combined; S. nigra was often sister to the other species examined. K.-Y. Chen et al. (2024) looked at the plastomes of 128 species (44 sections, 7 subgenera) and 124 single-copy orthologues from 66 accessions[; the plastomes provided less resolution. M. Wang et al. (2019: genomes of 80 individuals, 29 species) found extensive conflict between the various analyses they carried out in Populus, but of the taxa that they examined P. mexicana seemed to be sister to the rest of the genus.
Classification. Alford (2003) recognised three families for the New World genera previously included in Flacourtiaceae and here included in Salicaceae (e.g. A.P.G. 2017). Chase et al. (2002) provide a detailed tribal classification for Salicaceae s.l.. Comparing their tribes with the three groups of genera of Salicoideae recognized by Z.-S. Zhang et al. (2021), we find Banarieae, Bembicieae, Homalieae and Prockieae are all in Group 3, and if the topology there were to hold up, at least two more tribes would be necessary - a more detailed phylogeny that includes data from the nuclear genome is much needed. Oncoba is a very distinctive plant (see above); it is unplaced; it belongs in Salicaceae, although the other genera that were included in Oncobeae are in Achariaceae-Lindackerieae. Generic limits in the Casearia group need attention (Samarakoon et al. 2010); Samarakoon and Alford (2019) delimit Casearia broadly - see also de Mestier et al. (2022).
Applequist (2017) provided a sectional classification for the large genus Homalium, and L. Zhang et al. (2017: 9 species examined) provide a chloroplast (maternal) sectional phylogeny of Populus recognizing six sections; not all these sections in could be recovered in the more extensive studies of M. Wang et al. (2019). Marincek et al. (2024, also N. D. Wagner et al. 2023) noted that both subgenera and sections in Salix needed re-evaluating, but Marincek et al. hold out hope for the future.
Previous Relationships. It had been observed in the past that Salicaceae s. str. and Flacourtiaceae-Idesieae were close, despite the catkins of the former, previously thought to be an important character, and more conventional inflorescences of the latter (summary in Chase et al. 2002) - they both have similar, distinctive leaf teeth, phenolic-type compounds such as salicin, etc. (R. B. Miller 1975), while rusts and caterpillars, perhaps keying in on chemical characters, show similar distributions (e.g. Meeuse 1975b; see above). However, in the Englerian system Salix was often kept with the wind-pollinated Amentiferae, not at all close to Flacourtiaceae, a family that was also recognised at the time and which encompassed the bulk of Salicaceae above and also Achariaceae, etc.; see also Gilg's (1914) emphatic rejection of Hallier's suggestion that there might be a link between Idesia (Flacourtiaceae) and Salicaceae. Cronquist (1981) placed Salicaceae in a monofamilial order, but next to his Violales.
Thanks. I am grateful to S. Zmarzty for comments.
[Euphorbiaceae [[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]] / Euphorbioids: flowers small, often imperfect and plant often monoecious, 3-merous or not; ovules 1-2/carpel, inner integument usu. thicker than outer, epitropous, (apex of nucellus decidedly pointed [= nucellar beak]), obturator +; fruit a part-septicidal + loculicidal capsule/schizocarp; cotyledons longer and broader than radicle.
Age. This node has been dated to (74-)71, 69(-66) Ma (Wikström et al. 2001), ca 82.8 Ma (Tank et al. 2015: table S1, S2) and (108.6-)102.5(-94.8) Ma (Xi et al. 2012b: table S7).
Evolution: Divergence & Distribution. Lee et al. (2011) found that genes involved in oxygen and radical detoxification clustered at this node, but Populus and Bruguiera were the only other members of Malpighiales in their study.
On strict parsimony grounds the distinctive fruits of Euphorbiaceae s.l., i.e. including Phyllanthaceae and Picrodendraceae, could be a synapomorphy for the whole clade, or acquired twice or more, but in any case, they have been lost several times.
Seed Dispersal. Esser (2003b) emphasized that over 50% of Malesian Euphorbiaceae s.l. were zoochorous, notably more than in other tropical areas.
Chemistry, Morphology, etc.. A nucellar beak, that is, a protrusion on the parietal tissue of the ovule above the embryo sac and through the micropyle, is known from Euphorbiaceae, Phyllanthaceae and Picrodendraceae, but not from Ixonanthaceae or Linaceae. Recent work has shown that in Pera the nucellar apex is rounded when the megaspore mother cell divides and the parietal tissue is 2-4 cells across, but as the ovule develops the parietal tissue becomes much thicker and the apex of the nucellus becomes pointed, although remaining enclosed by the inner integument (see Franca & De-Paula 2017: c.f. Figs 3d, e, with 4a, 5a - no comment made on the change). In some Euphorbiaceae, e.g. Alchornea, the nucellar beak does not protrude through the endostome (Gama et al. 2019). Further work is needed to clarify both the character itself - is the pointed apex developed from a nucellar cap, or from periclinal divisions of other nucellar cells? - unclear in Pera, huge nucellar cap in Codiaeum (Bor & Bouman 1979) - and its distribution. Franca and De-Paula (2017) suggest that the patterns of cell division in the young embryo vary in an interesting way.
Many older works on Euphorbiaceae also contain information about Phyllanthaceae and/or Picrodendraceae, not to mention other families in Malpighiales like Putranjivaceae - see e.g. Baillon (1858: something of a classic), Michaelis (1924: floral morphology), especially Webster (1967, 1994a, b, 2013), Radcliffe-Smith and Esser (2001), Hegnauer (1966, 1989: chemistry), Uhlarz (1978: stipules), also other papers in Ann. Missouri Bot. Gard. 81. 1994, etc., indeed, older literature for the individual families will be under Euphorbiaceae - check that family in particular for possible general references.
Classification. Merino Sutter and Endress (1995) argue for a rather broadly delimited Euphorbiaceae (including both Phyllanthaceae and Putranjivaceae), Huber (1991) for a narrower circumscription, with the biovulate taxa (Phyllanthaceae, Picrodendraceae, Putranjivaceae) being considered to be closer to Linales s. str., while Meeuse (1990) also suggested that the family should be split - into eleven families. There is no molecular evidence for a broadly delimited Euphorbiaceae (unless - perhaps - Linaceae et al. were to be included), yet Euphorbiaceae s. str, Phyllanthaceae and Picrodendraceae do have similar and rather distinctive capsules (see also Merino Sutter et al. 2006). Recent work by Xia et al. (2020) also confirms that the break up of Euphorbiaceae s.l. makes sense, although exactly where the bits are all to go remains unclear there.
Molecular analyses by Wurdack and Chase (2002), and especially by Wurdack et al. (2005, see also Tokuoka 2007) and Xi et al. (2012b), all suggested that substantial changes were needed to the groupings that had been recognised in Euphorbiaceae s.l.. The reclassification they proposed is followed below, but the inclusion of Rafflesiaceae (see C. C. Davis et al. 2007) and the associated recognition of Peraceae are no longer necessary.
Previous Relationships. Cronquist's Euphorbiales included Simmondsiaceae (Caryophyllales here), Pandaceae (elsewhere in Malpighiales) and Buxaceae (Buxales), whilst Takhtajan's (1997) Euphorbianae included Pandaceae and Dichapetalaceae (here elsewhere in Malpighiales), as well as Thymelaeaceae (Malvales) and Aextoxicaceae (Berberidopsidales), but these groups clearly have little to recommend them. Bhatnagar and Kapil (1994) discuss the bearing of embryological data on the circumscription and relarionships of Euphorbiaceae s.l. at a time when it was becoming apparent that many of the previous ideas of the circumscription and relationships of Euphorbiaceae were questionable.
EUPHORBIACEAE Jussieu, nom. cons. - Back to Malpighiales
Vessel elements with simple perforation plates; (laticifers +); plant dioecious; flowers imperfect; G [3], styles ± separate; ovule 1/carpel; fruit with outer pericarp often separating from the woody layer, valves falling off, central column persistent; seeds large, micropylar caruncle + (0); exotegmen sclereids laterally flattened, oblique [Malpighian cells, "palisade"].
223 [list]/6,880 (6,252)- four groups below. Pantropical, also (cool) temperate. Map: see Meusel et al. (1978, Canada not very accurate). Photo Flower, Flower, Fruit, Fruit.
Age. The age of this node is (103.8-)97.2(-87.1) Ma (Xi et al. 2012b: table S7).
1. Peroideae Hasskarl —— Synonymy: Peraceae (Baillon) Klotzsch
Shrubs to trees; petiole bundles interrupted arcuate to annular, complete annular, (also with medullary plate and wing bundles); stomata?; leaves spiral, lamina vernation involute, margins entire, venation pinnate, stipules (large), small or 0; (plant dioecious, inflorescence pseudanthial, coloured inflorescence bracts +); staminate flowers: C (clawed), (0 - Pera); nectary lobes opposite K (0 - Pera); A 2-8, (androgynophore +; A extrorse, opposite C - Clutia); tapetal cells 1-2-nucleate, with prismatic crystals and styloids; pistillode + (0); carpelate flowers: (K 0, C 0 - Pera); staminodes 0; stigma bilobed (trumpet-shaped - Pera); endothecial cells reverse T-shaped/pyriform; micropyle zig-zag, outer integument 3-6 cells across, inner integument 3-6 cells across, parietal tissue initially 2-4 cells across, hypostase becoming massive [cells in files]; fruit septa membranous, without visible vascularisation [with aerenchyma, disintegrates in mature fruit], (valves connected at base), perianth [when present] persistent; seeds carunculate [testa] or arillate, black, shiny; (exotesta palisade, lignified), endotesta with crystals, exotegmen tracheoidal, oblique; endosperm copious; n = 18, chromosomes 0.5-1.1 µm long.
5 [list]/135: Clutia (75), Pera (40). Pantropical, probably not East Malesia (one doubtful report). Map: see van Welzen (1994a).
Age. The age of this node is (93.4-)63.5(-32.1) Ma (Xi et al. 2012b: table S7 - Pog Per Clu).
[Cheilosoideae [Acalyphoideae [Crotonoideae + Euphorbioideae]]: (Al-accumulators); cucurbitacins [triterpenes], (polyacetylenes), ellagitannins [geraniin and mallotussic acid], lectins [hemagglutinins], cocarcinogens + [phorbol ester diterpenes]; cork also outer cortical (pericyclic); vessel elements often in multiples, (with scalariform perforation plates); (pits vestured); sieve tubes with non-dispersive protein bodies; (nodes also 5 or more:5 or more); petiole anatomy very variable, often ± annular, etc.; (epidermis silicified); stomata various; leaves spiral, two-ranked or opposite, lamina vernation variable, venation pinnate, margins entire or single veins running into opaque persistent tooth, paired abaxial subbasal glands +/0 (also elsewhere), petioles often apically pulvinate, stipules (0), with axillary colleters; P/K (2-)3-6(-12), (connate); nectary ± annular (0), outside A; staminate flowers: A introrse, loculi 2; pistillode 0; carpelate flowers: staminodes 0; G [(2-)3(-many)], median member usu. abaxial, style (short), stigmas prominent, often branched or with adaxial furrow, dry or wet; micropyle exostomal, nucellar beak and cap +; endosperm usu. copious, embryo chlorophyllous or not; x = 11 (?10), nuclear genome [1 C] (0.045-)1.064(-25.072) pg.
Age. Estimates for the crown group age of this clade are ca 102 Ma (van Ee et al. 2008) and (94.7-)89.9(-81.2) Ma (Xi et al. 2012b; Table S7 - check).
Fruits, Euphorbiotheca deccanensis, some 66 Ma have been described from the Deccan Traps (Reback et al. 2021).
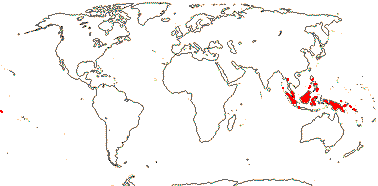
2. Cheilosoideae (Müller Argoviensis) K. Wurdack & Petra Hoffmann —— Synonymy: Cheilosaceae Doweld
Petioles pulvinate; C 0; staminate flowers: A 5-12; pollen echinate; carpelate flowers: style bifid; G ([2]); ovule with outer integument 8-10 cells across, inner integument 8-12 cells across; seeds not carunculate; testa vascularized, exotesta fleshy; endosperm +, ?embryo morphology; n = ?
2/7: Neoscortechinia (6). Myanmar, Malesia. Map: from van Welzen (1994b).
[Acalyphoideae [Crotonoideae + Euphorbioideae]]: (herbs, lianas); (diterpenoids, inc. phorbol esters +); laticifers +; [?here]; (leaves opposite); monoecy common; C 0, 3-8; pistillode 0/+; outer integument 6-10 cells or so across; (fruit indehiscent), caruncle +/-.
Age. This node is estimated to be around (90-)86.4(-81) Ma (Xi et al. 2012b: Table S7, Acalypha + Suregada) or (84.1-)62.8(-45.0) Ma (K. Yan et al. 2025).
Intertrappean fossils (late Cretaceous: Maastrichtian) from India ca 66 Ma have features of Mallotus and especially Croton (Wheeler et al. 2017).
3. Acalyphoideae Beilschmied
(Nodes 1:1 + split laterals); (cristarque cells +); petiole with ring of (fused) bundles (with fibrous sheath), medullary bundles 1-several; lamina (palmately compound/venation palmate); P (one-trace); (nectary 0); staminate flowers: P valvate; A 2-many, (connate); tapetum (amoeboid), cells 2-4-nucleate; pollen grains (tricolpate), (inaperturate), etc., (tricellular); (G 0); carpelate flowers: P decussate [?level]; (A 0); G [(2-4)]; (ovule adnate to axis), (micropyle bistomal/zig-zag), outer integument 3-6(-16) cells across, inner integument 3-24 cells across, (nucellar beak 0), hypostase +; (testa vascularized/outer 2 layers persistent), exotegmic cells radially elongated, palisade, slightly curved; (endosperm 5-10-ploid), cotyledons longer and broader than radicle; n = 9-12.
99/1,865: Bernardia (>50), Ditaxis (45). Pantropical.
Age. Crown-group Acalyphoideae may be (101.6-)92.6(-84.2) Ma (Cervantes et al. 2016).
3A. Plukenetieae Hutchison —— Synonymy: Tragiaceae Rafinesque
Herbs, twining lianas or vines (to small trees); (ectopic cambia + - Dalechampia [D.]); plant with stinging hairs, (vascukarized) colleters +; (inflorescence pseudanthial, inflorescence bracts two, large, petal-like - D.); K 3-6; A 3-many, connective apiculate, (anthers shorter than filaments, thecae separate), (stinging hairs at apex of filaments); pollen 3(-5) colpate, (3-porate), (apertures 0); style slender to stout, stigma lobes short to long, recurved/stigma-style massive, ± spherical or truncated/pretty much indescribable; (outer integument vascularized - Dalechampiinae).
Ca 18/350: Tragia (150), Dalechampia (120). Pantropical.
3B. Acalypheae Dumortier —— Synonymy: Acalyphaceae Menge, Mercurialaceae Berchtold & J. Presl, Ricinaceae Martynov, Trewiaceae Lindley
(Annual) herbs, shrubs to trees; (anthers long, pendulous, twisting after dehiscence - Acalypha [A.]), (loculi 3–4 - Macaranga); pollen brevicolporate; (styles multifid - A.); nucellar cap ca 8 cells across [Micrococca]; embryo sac tetrasporic, 12-16 celled - Penaea type.
Acalypha (500), Macaranga (260), Mallotus (115), Claoxylon (80). Pantropical (temperate), mostly Old World.
Age. Macaranga/Mallotus-type fossils are reported from the Patagonian Laguna del Hunco Early Eocene deposits ca 52 Ma (Wilf et al. 2023).
3C. Alchorneae Dumortier
endothelium + [Alchornea].
7/: Alchornea (40).
[Crotonoideae + Euphorbioideae]: petiolar/lamina glands common [?level]; pollen grains (tricellular - ?level), (inaperturate).
Age. The age of this clade is estimated to be (69.4-)47.1(-26.1) Ma (K. Yan et al. 2025).
4. Crotonoideae Beilschmied
Cyanogenesis via the valine/isoleucine pathway; laticifers articulated or not; hairs often stellate or lepidote; (petiolar glands +), leaf (palmate); (inflorescence thyrsoid, female and male flowers at base, male towards apex) [= crotonoid]; (K connate), staminodes +, secretory, opposite P/K; staminate flowers: A 3-many, (filaments 0); carpelate flowers: (micropyle endostomal), outer integument 4-8 cells across, inner integument (8-)18-25 cells across, nucellar beak +; micropylar megaspore functional; seeds arillate or not, often pachychalazal; (sarcotesta +), (exotesta palisade, endotestal cells ± palisade, thin-walled, slightly lignified), tegmen vascularized, exotegmen cells elongated ?anticlinally, quite stout; (perisperm +, slight); (100+ bp deletion in trnL-F spacer); n = (9-10) 11 (12, 14).
68/2,065. Pantropical (warm temperate).
4A. Suregadeae Reveal
Pollen pantoporate.
2/32: Suregada (31). Old World tropics, esp. Madagascar.
[[Elateriospermeae [Manihoteae [Adenoclineae + Micrandreae]]] [[Crotoneae + Jatropheae] [Trigonostemoneae, Aleuritideae, Codiaeae, Elateriospermeae, Ricinocarpeae]]] / [Articulated + Inaperturate Crotonoids]: inner integument vascularized.
[Elateriospermeae [Manihoteae [Adenoclineae + Micrandreae]]] / Articulated Crotonoids: pollen pantoporate.
Elateriospermeae G. L. Webster - Elateriospermum tapos Blume
Trees; C 0; staminate flowers: A 10-20, anthers with apical gland; G ± 0; carpellate flowers: staminodes many, subulate; style dilated apically; seed ecarunculate, ± flshy; endosperm sparse, cotyledons large.
1/1: Southern Thailand and West Malesia, but not the Philippines.
[Manihoteae [Adenoclineae + Micrandreae]]: ?
Manihoteae Pax
(Herbs) shrubs to trees; (stinging hairs + - Cnidoscolus); venation palmate.
2/285: Manihot (170), Cnidoscolus (93). New World, mostly tropical, inc. the Antilles.
[Adenoclineae + Micrandreae]: pollen 3-colpate.
Adenoclineae G. L. Webster
6/42: Tetrorchidium (23), Endospermum (11). Tropics.
Micrandreae G. L. Webster
3/23: Micrandra (12), Hevea (10). South America.[[Crotoneae + Jatropheae] [Trigonostemoneae, Aleuritideae, Codiaeae, Elateriospermeae, Ricinocarpeae]] / Inaperturate Crotonoids: pollen with ± triangular supractectal elements attached to muri with short and irregular columellae [Crotonoid pollen], apertures 0.
[Crotoneae + Jatropheae] / C1 group: ?
Crotoneae Dumortier —— Synonymy: Crotonaceae J. Agardh
Herbs ro trees; hairs variously branched/stellate (simple); foliar extrafloral nectaries pale, abaxial, paired/on petiole, also in sinuses of lamina margin; colleters at apex of petiole; C ± 0; staminate flowers: filaments incurved in bud; carpelate flowers: K (unequal), C (filamentous); A 0; chalazal endosperm haustorium to ca 1000 µm long.
6/1,178. Pantropical.
Croton Baillon
staminate flowers: C+; a 11-20; carpelate flowers: C reduced/0; style branches bifid.
1,150 spp. Pantropical/warm temperate, esp. Brazil, Madagascar, etc..
Outer A whorl opposite C
(A development chaotic)
(K, nectaries monosymmetric); (C 0); outer A whorl opposite C first to develop, centrifugal development 0 G ([2], 1).
?
3. Jatropheae Baillon (inc. Joannesieae)
Herbs to trees;
8/181: Jatropha (170). Tropical.
[Trigonostemoneae, Aleuritideae, Codiaeae, Ricinocarpeae] / C2 group: ?
3. Trigonostemoneae G. L. Webster
Trees to shrubs; leaves ± glandular; C often conspicuous; staminate flowers: A 3-5, connate, extrorse, connective expanded; pollen (supratectal elements as spines/± connate); G 0; carpellate flowers: styles ± free, bifid (undivided); K ± enlarged in fruit (not); seeds ecarunculate.
4/84: Trigonostemon (65). China to Malesia.
Aleuritideae Hurusawa (inc. Neoboutonieae)
(stipules minute - Garcia); plant often dioecious.
14/55: Crotogyne (16). Tropics, esp. Africa-Madagascar, also mainland Southeast Asia to Australia and Polynesia.
Codiaeae Hutchinson
?
16/108: Codiaeum (17), Dimorphocalyx (17), Baloghia (15). India, China, the Himalayas to Eastern Auatralia and New Caledonia, Mexico and the Antilles to South America.
Ricinocarpeae Müller Argoviensis —— Synonymy: Bertyaceae J. Agardh, Ricinocarpaceae Hurusawa
Latex +, reddish; indumentum stellate (unbranched or dendritic); lamina involute, stipules 0 (+); plant monoecious (dioecious), inflorescences terminal (axillary or pseudoterminal); disc +/0; pistillate flowers: C +, well-developed (0); A many, extrorse, filaments often partly connate; pollen (supratectal elements indistinct); fruit (drupaceous); seeds carunculate (0).
8/87: Bertya (28), Ricinocarpus (28), Beyeria (24). Mainly Australia and New Caledonia, few East Malesia, one Borneo (Sabah).
5. Euphorbioideae Beilschmied
Laticifers not articulated; (nodes 1:1); lamina vernation involute [Madea]; staminate flowers: A not covered by P, (extrorse); A 1-20(-80); pollen grains (tricellular); nectary 0; ovule outer integument (8-22 cells across, inner integument 3-7(-22) cells across, nucellar beak +; testa (mucilaginous), (vascularized), tegmen (vascularized), (two layers of tegmic sclereids, fibres between); n = 6-11; nuclear genome [1 C] (292-)3825(-14034) Mb [?level].
25/2,810. Pantropical, extending into temperate regions.
[[Neoguillauminia + Calycopeplus] + Euphorbieae]: ?<
Age. The age of this node is (61.9-)54.7(-47.5) Ma (Horn et al. 2014).
5A. Neoguillauminia + Calycopeplus: ?
2/6: Calycopeplus (5). Australia, New Caledonia.
5B. Euphorbieae Dumortier - Euphorbia L. —— Synonymy: Tithymalaceae Ventenat
Habit various; (Crassulacean Acid Metabolism +); starch grains much elaborated; pseudanthia + [= cyathia, usu. made up of 5 groups of staminate flowers and 1 carpellate flower]; nectaries +, formed by fusion of stipules of adjacent inflorescence bracts; P 0; staminate flowers: bracteole; A 1; pollen apertures with banded intine thickenings; carpellate flowers: (K +); style hollow; ovule (campylotropous), micropyle (zig-zag), outer integument 4-7 cells across, inner integument 8-16 cells across, parietal tissue 5-11 cells across, nucellar cap ca 2 cells across, chalazal zone ± massive [= postament + hypostase +], (obturator hairy); (archesporium multicellular); seed (carunculate - outer integument); testa (with sphaerocrystals), tegmen (multiplicative).
1/2,157. Pantropical, also temperate.
Age. Crown-group Euphorbia is (54.7-)47.8(-41.0 Ma (Horn et al. 2014); other estimates are (47.2-)36.6(-29.0) Ma (Bruyns et al. 2011; see also Horn et al. 2012).
5B1. Euphorbia subg. Esula Persoon
Stipules 0; tapetum amoeboid.
460 spp. Worldwide, esp. Eurasia, not cold conditions, Malesia, Australia, few in South America. Map: Bruyns (2022: Fig 1:67).
Age. Age estimates for crown-group Euphorbia subgenus Esula are (41.4-)31.4(-23.4) Ma (Bruyns et al. 2011) and (47.7-)40.5(-33.2) Ma (Horn et al. 2014).
[Athymalus [Euphorbia + Chamaesyce]]: ?
Age. The age of this node is (51.4-)44.7(-37.8) Ma (Bruyns et al. 2011); another estimate is over 42.5 Ma (van Ee et al. 2008: subgenus Esula not included). 5B2. Euphorbia subg. Athymalus Reichenbach Plants usu. succulent, cactiform, (stem ridged) [esp. Africa]; thorns +, often peduncular/branches; leaves often reduced, deciduous/caducous, (stipules 0); seeds not carunculate; n = 10. 147 spp. Africa, Macaronesia, Arabian Peninsula to Iran. Map: Bruyns (2022: Fig 1:66). Age. Diversification in crown-group subgenus Athymalus is estimated to have begum (38.2-)28.1(-22) Ma (Bruyns et al. 2011) or (33.5-)24.6(-16.3) Ma (Horn et al. (2014). Age.The age of this clade is estimated to be (45.6-)39.1(-32.9) Ma (Horn et al. 2014). 5B3. Euphorbia subg. Euphorbia Dumortier Plants usu. succulent, cactiform, (stem ridged]) [esp. Africa]/(herbaceous; trees); spines +, of various sorts + [from stem; spine shields; ≡ stipules]/0; lamina (succulent) (deciduous) (much reduced); n = 12-19; seedling with green-mottled stem. 660 (-?1,000) spp. Tropical-Warm Temperate, esp. Africa, Madagascar. Map: see Bruyns (2022: Fig 1:68). Age. Crown-group subgenus Euphorbia may be around (39.6-)29.9(-22.5) Ma (Bruyns et al. 2011) or (36.4-)30.8(-24.6) Ma (Horn et al. 2014). 5B4. Euphorbia subg. Chamaesyce Rafinesque Plants annuals/(succulents)/(trees); (C4 photosynthesis - sect Anisophyllum); main axis aborts or not; (branching plagiotropic - An.); (stipules 0); inner integument 2-3 cells across. 570 spp. Worldwide. Age. The age of Euphorbia subgenus Chamaesyce is estimated to be (37.6-)28.1(-20.6) Ma (Bruyns et al. 2011) or (42.1-)36.0(-30.3) Ma (Horn et al. 2014). 5C. Hippomaneae Dumortier (inc. Hureae, Pachystromateae) —— Synonymy: Hippomanaceae J. Agardh Shrubs to trees; flowers sessile2; K connate1, (C 0); staminate flowers: K (not vascularised); A (13-many, whorled); pollen tricolporate, margo smooth; carpellate flowers: G (-many - Hura), (alt. with 3 P - Excoecaria); ; ovules pachychalazal, bistomal, nucellar beak short, integuments ca ≥6 cells across, outer integument vascularised, inner integument with vascular bundles at base, obturator +; septum with 11/>32 vascular bundles. 24/390: Gymnanthes (45), Excoecaria (40), Mabea (40). I start off by discussing Euphorbia, distinctive and much studied, before turning to the rest of the family.
Divergence & Distribution. Divergence within the very speciose Euphorbia - it includes around 2,160 species - may have begun (63.5-)48.9(-40.5) Ma (Bruyns et al. 2011: Dichostemma, Anthostemma). Indeed, Euphorbia s. str. (i.e. not including Chamaesyce, etc.) was already extremely variable (Keller 1996), but with the inclusion of Chamaesyce and other segregate genera (e.g. Horn et al. 2009a, 2012) there is yet more diversity. Basic relationships within the genus are [Esula [Rhizanthium (= Athymalus) [Euphorbia + Chamaesyce]]] (for subgeneric names, see Bruyns et al. 2011, also above). Divergence within the largely succulent subgenera Athymalus and Euphorbia is estimated to have occurred (38.2-)28.1(-22) and (39.6-)29.9(-22.5) Ma respectively, with much speciation in the latter subgenus in particular occuring within the last ca 13 Ma or so (Bruyns et al. 2011). Subgenera Athymalus and Esula are largely Old World, subgenus Chamaesyce is largely New World, and subgenus Euphorbia is global, but not where conditions are cold. There seems to have been but a single origin of the distinctive cyathium that is an apomorphy for the genus (Park & Backlund 2002; Wurdack et al. 2005), and although it characterises a very species-rich clade, diversification here may also be associated with the evolution of a variety of distinctive life forms (and around 500 species have spiny structures of various morphologies - Wurdack 2023), seed dispersal mechanisms, and CO2 concentrating mechanisms (see below). For fruit and seed morphology in subgenus Esula, see Pahlevani et al. (2015), and for the biogeographic implications of relationships in the genus, see e.g. Dorsey et al. (2013) and Riina et al (2013); there has been but a single dispersal event of Euphorbia to India and Southeast Asia (Horn et al. 2014). Coello et al. (2024) looked at the distribution of a clade in subgenus Euphorbia in which E. canariensis was sister to a group of species in Asia some 11,000 km distant. Probably dispersal + extinction has been involved, however, E. abdelkuri, from Socotra, may be sister to the whole clade and there are other unsampled taxa from the N.E. African region that are apparently involved. Subgenus Chamaesyce has undergone notable diversification on Hawai'i, some species there being C4 trees growing in mesophytic forest (Y. Yang et al. 2009, 2012; Yang & Berry 2011b). Ecology & Physiology. Both growth patterns/plant morphology and carbon fixation pathways are very diverse in Euphorbia, the latter in particular vary little elsewhere in Euphorbiaceae. The evolution of the annual habit and the cactus-like growth form are associated with much speciation (Horn et al. 2009a, 2010b). The annual habit, common in Euphorbia subgenera Chamaesyce and Esula, has evolved eight times or more in subgenus Esula alone (Frajman & Schönswetter 2011; Riina et al. 2013; see also Peirson et al. 2014). The whole plant body of some species of Euphorbia-Chamaesyce-sect. Anisophyllum seems comparable with the inflorescences of other species of the genus - the seedling apex aborts after the production of a single pair of leaves, and several axillary shoots showing complex determinate and unequal branching patterns and with cyathia throughout represent the adult plant (Hayden 1988). Stem succulence in Euphorbia - some are quite massive stem succulents, and they may have spines of various sorts (see below: Vegetative Variation) - is associated with the evolution of a monopodial growth form and axillary inflorescences (Horn et al. 2012, 2014; see also Dorsey et al. 2013); these plants are cactus-analogues in drier areas throughout Africa and into India and even beyond, although they are also known from the New World, e.g. subgenus Euphorbia sect. Brasilienses (Steinmann & Porter 2002; Bruyns et al. 2006, 2011). Succulent species of Euphorbia are a particularly prominent component of the winter rainfall vegetation of the Succulent Karoo of south west Africa (Nyffeler & Eggli 2010b; see also Mauseth 2004b for succulence). Plants of subgenus Euphorbia in Madagascar that are chamaephytes, with tuberiform structures and succulent leaves, grow in the drier, but not particularly warmer, areas, but surprisingly, cactiform species showed no particular correlations with environmental features on the island (Evans et al. 2014). Xeromorphism of one form or another has evolved ca 14 times in the genus (Horn et al. 2012). All told, some 850 species belonging to all four subgenera are succulents, succulence having evolved ten times or so, and in subgenera Athymalus and Euphorbia the succulent habit is particularly common, both subgenera lacking annuals, although basal clades here are at most semi-succulent (Horn et al. 2010b; Bruyns et al. 2011; Morawetz & Riina 2011; Dorsey et al. 2013; see Eggli 2002 for an enumeration of taxa; Frandsen 2017 and Bruyns 2022 for photographs). Indeed, some clades of Euphorbia are important components of the Succulent Biome, especially in Africa-Arabia (the biome also occurs in America); other major groups in that biome include Bursera and co., Fabaceae, Cactaceae, Didiereaceae, etc. (Gagnon et al. 2018 and references). Arakaki et al. (2011) suggest that succulent groups in general radiated/diversified quite recently in the mid to late Miocene to Pliocene around 10 Ma or so, even if the clades involved originated substantially earlier, and they also mentioned core Ruschioideae, Cactaceae-Opuntioideae and Cactoideae, and Agavoideae in this context (in all cases, see Ecology & Physiology) - and add Aloe, Crassula, etc. (M. Lu et al. 2021). Most of the ca 350 species of subgenus Chamaesyce sect. Anisophyllum subsect. Hypericifoliaecarry out C4 photosynthesis. This probably originated once here, and this is the largest C4 clade in the eudicots, around double the size of the next largest, Atriplex (Y. Yang & Berry 2011a; Yang et al. 2012; Horn et al. 2014; Sage 2016). Some C4 species grow in the more arid parts of Africa and are also succulents. Species growing on Hawai'i form a distinctive woody radiation and include some of the largest C4 plants anywhere, some being trees up to 9 m tall that grow in mesic forests (Pearcy & Troughton 1975; Robichaux & Pearcy 1984: see Winter 1981 for slightly larger C4 Chenopodioideae); Horn et al. (2012) discuss how the tree habit might evolve in a clade in which all branches are basically plagiotropic. This Hawaiian clade appears to have evolved from within a small clade of weedy annuals now found in the southern USA, Mexico, and the Caribbean (see also Yang et al. 2009); hybridisation and polyploidy was involved in their pre-dispersal origin, as is also known from elsewhere in the C4 clade (Yang & Berry 2011b). Yang et al. (2018) note the recurrent origin of these forest species on Kauai and Niihau in the Hawaiian archipelago from more widely distributed species of open habitats, a kind of taxon cycle à la E. O. Wilson; the forest species have larger seeds that are neither sticky nor buoyant. The major C4 clade in sect. Anisophyllum, predominantly herbaceous, probably evolved in drier areas of North America, and it is sister to the small subsection Acutae from southwest North America which includes C4, C3, and C2 herbs, although only species with the two latter kinds of photosynthesis have been sequenced (Y. Yang & Berry 2011a, esp. b; Yang et al. 2012). T. Sage et al. (2011) described C2 photosynthesis in subsection Acutae, and they noted that E. angusta, with C3 photosynthesis, had some anatomical similarities with C2 and C4 taxa. Horn et al. (2012) emphasize that it is not simply the evolution of the C4 pathway, but also the adoption of a plagiotropic branching habit, etc., that may have made this clade successful, as well as its movement from the Old to the New World (see also Yang & Berry 2011b). The crown age of the C4 clade is (20.2-)15.3(-10.5) Ma, and its stem age is (29.3-)23.6(-18.1) Ma (Horn et al. 2014). This origin of C4 photosynthesis is somewhat earlier than that of CAM photosynthesis, and aridity may not have been a driver here (Horn et al. 2014), note, however, that some estimates of the age of origin of the C4 pathway are as recent as (13.1-)10.4(-7.3) Ma (Christin et al. 2011b). Given the prevalence of succulence in Euphorbia and its association with drier habitats, it is not surprising that CAM is also quite common here (it is also known from a few species of Jatropha). Horn et al. (2014) looked at diversification here in the context of CO2 concentration mechanisms, and found that clades of perhaps 7 of the at least 17 independent acquisitions of CAM photosynthesis (five were in Africa-Madagascar) and the clade representing the single acquisition of C4 photosynthesis showed increased diversification rates. The former changes occured in the context of drying climates after the Mid-Miocene ca 14 Ma (estimates of the crown ages of the clades are (12.9-)7.1,2.2(-1.3) Ma, although that of Euphorbia-Euphorbia is estimated to be (17.8-)14.0(-10.5) Ma). Ancestors of these CAM clades were more or less woody plants, and in the Old World they had axillary inflorescences; adoption of CAM photosynthesis is quite often associated with increased diversification rates (Horn et al. 2014). Overall perhaps 850 species of Euphorbia are CAM plants, and they are concentrated in subgenera Euphorbia and Rhizanthium; Holtum (2023) discusses the evolution and distribution of these plants - overall, CAM may have evolved independently some 16–21 times, mostly from the Miocene onwards (see also Horn et al. 2014). Pollination and Seed Dispersal. Euphorbia is by far the biggest flowering plant clade characterized by pseudanthia, a.k.a. cyathia (for pseudanthia in Euphorbiaceae, see Baczynski & Claßen-Bockhoff 2023). The cyathium of Euphorbia is best interpreted as a modified cymose inflorescence. There is a single, terminal, carpelate flower surrounded by male flowers all reduced to a single stamen and with an articulation on their stalks representing the pedicel-flower junction (see the basically similar arrangement in Jatropha, etc.). Details of the development of the cyathium are provided by Prenner and Rudall (2007), and although these authors thought that the morphological nature of both the cyathial glands and the petal-like bracts surrounding the cyathium was unclear (see also Hoppe & Uhlarz 1981; Hoppe 1985 and Prenner et al. 2008b), the glands, at least, may be modified commissural stipules (Steinmann & Porter 2002) or derived from involucral bracts (Gagliardi et al. 2016) - not necessarily incompatible descriptions. For more on cyathia and their development, see e.g. Croizat (1937), Gagliardi et al. (2018). "Floral" genes may be expressed in the cyathium as a whole (Prenner et al. 2011). Cyathia are surrounded by bract-like structures that vary considerably in shape and colour, and some or all of these also have conspicuous nectaries. Several species of oligolectic pollinators may visit a single species of Euphorbia, overall, there may be hundreds of species of insect visitors (Ehrenfeld 1979); Asenbaum et al. (2021) discuss pollination - or lack of - by insect visitors in some European species of Euphorbia. New World species of Euphorbia that used to be segregated as Pedilanthus (= Euphorbia subg. Euphorbia sect. Crepidaria) have distinctive red, spurred, monosymmetric cyathia pollinated by birds, called a "key innovation" (Cacho et al. 2010), but there aren't many species in this clade... Around 2,300 species in the family, especially in Euphorbia, are likely to be myrmecochorous (Lengyel et al. 2009, 2010) - strictly speaking, most are likely to be diplochorous. Elaiosomes of one sort or another have evolved ca 13 times in Euphorbia alone (Horn et al. 2012), and are found in many of the ca 480 species of subgenus Esula (Riina et al. 2013; Peirson et al. 2014 for photographs). Seeds can get about in other ways, too. Thus a number of species of Euphorbia subgenus Chamaesyce section Anisophyllum in particular have a testa that becomes mucilaginous when wetted (Grubert 1974; Jordan & Hayden 1992). Vegetative Variation. Euphorbia is succulent in diverse ways. The plants may have variously articulated, cactus-like, or simply pencil-like succulent stems, the latter as in some species of subgenus Chamaesyce, or they may be medusoid, with relatively slender but still succulent branches radiating from a stout central axis. Bruyns et al. (2011) describe in detail the different forms of stem succulence found in the genus, and they note that in a number of species of subgenus Euphorbia the branches become permanently differentiated. Thus orthotropic axes with three-ranked leaves will not develop from plagiotropic branches with two-ranked leaves, and the latter when put upright in a pot simply keep growing vertically. Fleshy Euphorbia can be very like Cactaceae, as Bruyns (2022: p. 14) notes, "Most of the southern African species of Euphorbia have fleshy shoots with persistent photosynthetic epidermis that may live and function for decades, where the development of the bark is retarded or wholly prevented." Similarly, Euphorbia plants may be prickly in different ways (Bruyns et al. 2022 for a discussion). Thus in subgenus Euphorbia there are spine shields, also stipular spines (and there are a variety of other stipular structures), spines in the stipular position that are not actually vascularized, paired spines arising from the leaf base ("Dorsalstacheln"), and branched or simple thorns, and these latter may also do duty as inflorescences (see e.g. Park & Jansen 2007; Carter 2002, esp. illustrations, 2023 - succulent species; Bruyns 2010); Uhlarz (1974: extensive discussion) described the vasculature of his "Dorsalstacheln", showing how some bundles looped in and out, thence proceeding to the rest of the leaf. Stipules may be lacking in species of Euphorbia subgenus Esula (Riina et al. 2013), and 1:1 nodes have been reported from the genus (Sehgal & Paliwal 1974: also somewhat surprising 2:2 nodes). There is somewhat less variation in below-ground parts, although E. namuskluftensis has both tubers and rhizomes. Although latex might a priori seem to protect the plant, a diversity of bacteria and fungi (mean: 44 and 21 species repectively) were found in the latex of 38 species of Euphorbia that were growing in cultivation (Gunawardana et al. 2015). Perhaps some of these were endophytes, but between-species variation was great (up to seven fold), and what is actually going on in this system is unclear. Castelblanque et al. (2016) describe laticifer development in Euphorbia lathyris. Biesboer and Mahlberg (1981) looked at the complex morphology of the starch grains found in the latex of Euphorbia and laticifer evolution, while Bauer et al. (2014) examined latex coagulation, etc., in Euphorbia spp.. For the composition of Euphorbia latex, see references in Gunawardana et al. (2015) and Benjamaa et al. (2022); latex production in Euphorbia is also discussed by X. Zhao and Cai (2020). And now for the rest of the family... Evolution: Divergence & Distribution. A Mallotus/Macaranga-like plant has been found in deposits from New Zealand that are about 23 Ma old - this clade is not currently known from the islands (Lee et al. 2010). The oldest fossils from this taxonomic part of the world are from Eocene deposits 48.6-37.2 Ma (Nucete et al. 2012). There may have been shifts in diversification in the [Acalyphoideae [Crotonoideae + Euphorbioideae]] clade, and these have been dated to (61.9-)59.3(-57.1) Ma (Magallón et al. 2018). Diversification within Acalyphoideae occured within the last ca 70 Ma (Davis et al. 2005a). Some estimates for the divergence of Macaranga and Mallotus are as early as (79.1-)63.8(-63.3) Ma, but Van Welzen et al. (2014a: q.v. for many other ages) found that the two showed general congruence in their historial biogeography, e.g. both originating in Borneo and with similar dispersal patterns, and the crown group ages of the two were ca 32.7 and 34.3 Ma respectively, although their lineage-through-time plots differed considerably. However, the recent discovery of ca 52 Ma Macaranga/Mallotus fossils from Argentinian Patagonia (Wilf et al. 2023) will, if confirmed, completely upend previous ideas of the history of these genera; for more on possible southern ancestors of other extant Australo-Malesian taxa, see Fagaceae. Bansal et al. (2022b) found a diversity of crotonoid pollen morphologies in deposits in India (Intertrappean deposits, also in lignite mines dating from the Palaeocene to early Eocene) that they placed in Adenoclineae, Codiaeae, Crotoneae and Jatropheae. They suggested that Crotonoideae originated in Africa, moving to South America, Europe (the latter by way of the boreotropical dispersal route), and via the Kohistan—Ladakh Island Arc to India in the Late Cretaceous. From India the plants moved to Southeast Asia, although there was some extinction in India as climates changed during the Tertiary (Bansal et al. 2022b; see also Arecaceae, Ebenaceae and Dipterocarpaceae for similar movements). The stem and crown group ages for the large genus Croton (Crotonoideae) itself have been estimated at ca 55 and ca 40 Ma respectively (van Ee et al. 2008). Diversification of Croton on Madagascar represents a substantial radiation, and there has been further movement back to Africa and also to the Mascarenes and other islands close to Madagascar (Haber et al. 2017). Endemism is high in the Caribbean Leucocroton area, probably divergence from a common ancestor and reflecting such factors as adaptation to growth on the archipelago-like serpentine outcrops on Cuba (Jestrow et al. 2012, 2013; Cervantes et al. 2016; see also below). Somewhat under 2/3 the species of Acalypha are in the Americas, and almost 1/3 of these are from Mexico (Levin et al. 2022); understanding the obviously considerable movement within the genus will be of interest. R.-Y. Yu and van Welzen (2019) discussed the evoluion of the mostly Malesian clade [Dimorphocalyx [Ostodes + Trigonostemon]], a clade close to 29 Ma, paying particular attention to movement of the plants across Wallace's line. Matomoro-Vidal et al. (2012) looked at pollen evolution in Euphorbiaceae with a focus on variation in where callose was last deposited on the pollen grain walls, the last points of callose deposition (LPCD), which is affected by various aspects of pollen meiosis, and that in turn is related to aperture development (see also Ressayre et al. 2002). Interestingly, they found that there was variation in both the LPCD and in tetrad morphology in inaperturate Crotonideae, but not elsewhere in the family. Tokuoka (2007) discussed seed and ovule evolution. Ecology & Physiology. "Euphorbiaceae", i.e. including Phyllanthaceae, Putranjivaceae, etc., are often the second most abundant family in tropical rainforests in South-East Asia and Africa (Gentry 1988). Euphorbiaceae s. str. are common in Amazonian forests and they include a disproportionally high number of the common species with stems at least 10 cm across (ter Steege et al. 2013). They are also a notable element in the Neotropical seasonally-dry tropical forest (Pennington et al. 2006b and papers therein; Wurdack 2023). In some Brazilian species of Croton water from fog is taken up through epidermal emergences (including the stellate/lepidote "hairs"); some cells involved are subepidermal. Immediately below the emergences there are very thick-walled but at most slightly lignified sclereids that appear to be part of the whole absorbtive process (Vitarelli et al. 2016). Arévalo et al. (2017a) looked at the evolution of wood anatomy and habit in Croton s.l.. Hyperaccumulation of metals, etc., has been reported from the family. Reeves et al. (2007 and references) record high concentrations of nickel in the latex of genera like Euphorbia, Cnidoscolus, etc.. All 28 species of Cuban Leucocroton (= Croton s.l.) are reported to accumulate nickel (Reeves et al. 1996; Jestrow et al. 2010 and Cervantes et al. 2016 for phylogenies). Pollination Biology & Seed Dispersal. Flowers of Euphorbiaceae are generally small. Pseudanthia have evolved ca 4 times here, and in the perfect "flower" of Euphorbia, for example, each stamen represents a staminate flower - smaller than that you cannot really get. Pseudanthia in Pera, which is dioecious, have groups of its highly reduced flowers surrounded by coloured inflorescence bracts (Gagliardi et al. 2018; Baczynski & Claßen-Bockhoff 2023), while in Dalechampia (Acalyphoideae-Plukenetieae) and some species of Euphorbia inflorescence bracts may also be very large and brightly coloured; the carpelate flowers mature first, although in Dalechampia they are not terminal in the inflorescence (see also Gagliardi et al. 2016). The ca 120 species of Dalechampia have remarkable pseudanthia; here female megachilid, euglossine and meliponine (stingless Trigona) bees visit the "flowers" for triterpenoid resins that they use to build their nests - resin is a very uncommon pollination reward in flowering plants, and in a number of species it is coloured - c.f. coloured nectar (see Dressler 1982; Armbruster 1984, 1993, 1996; Armbruster et al. 2005; Tölke et al. 2019); a few male euglossines collect fragrances. The resin has secondarily become used for defence in some species of Dalechampia - this may have beenb its original fuction - and there is a reversal in the arrangement of the staminate flowers (Armbruster et al. 2009b). In some Madagascan Dalechampia there is buzz pollination, so pollen is the reward; buzz pollination is here derived from generalized pollination, in turn derived from resin flowers - and perhaps generalized pollination is derived from buzz pollination, although the former may have but a single origin (c.f. support values: Armbruster et al. 2013a). In these buzz-pollinated taxa, the staminate flowers are enclosed by the perianth, the pollen coming out of an opening at the apex that functions as a pore - c.f. porose anthers common in other buzz-pollinated taxa. Here the perianth members do not separate further, but in species pollinated in other ways, the perianth members of the staminate flowers reflex (Armbruster et al. 2013). Pollination by thrips (Thysanoptera) has been described in detail in the myrmecophytic Macaranga hullettii - Neoheegeria is the insect involved; here the bracteoles (sic) are the breeding sites, also providing protection, the unicellular trichomal nectaries on their adaxial surfaces providing food for both larvae and adults, the pollen grains are small, close to smooth, and with pollenkitt, and the flowers have a vanilla-like scent (Moog et al. 2002; see also Yamasaki et al. 2015: also disciform glands on bracteoles, or nothing). 24/29 of the myrmecophytic species in the genus may be so pollinated (based on floral morphology; see also Yamasaki et al. 2015), about double the frequency when compared with non-myrmecophytic species growing in the West Malesian localities that Fiala et al. (2011) visited. Wind pollination is known from e.g. some Acalyphoideae (Gama et al. 2019). Interfloral protogyny is common in Euphorbiaceae, perhaps associated with the fact that female flowers are often borne towards the base of the inflorescence, and so might be expected to open first (see Bertin & Newman 1993). Seed dispersal is initially usually by the explosion of the capsule, the walls twisting as they open, and larger seeds can be thrown quite some distance. Thus the seed of Hura crepitans, the sand-box tree, has an escape velocity of up to 252 kph (70 m s-1) and is hurled up to 30 (mode) or even 45 m (Swaine & Beer 1977). The woody wall is mesocarpial in origin and consists of a layer of fibres, a layer of anticlinally elongated sclereids, and then another layer of fibres at right angles to the first (Reback et al. 2021). Smaller seeds in particular may also have nutritive elaiosomes (e.g. Rössler 1943; Narbona et al. 2005) which facilitate local dispersal of the seeds, especially by ants. Ants disperse seeds of 68% of the 186 species of Euphorbiaceae growing in the Brazilian Caatinga, mostly from plants 5 m or less in height (Leal et al. 2013, 2015, 2017); the nature (size) of the reward affects the species of ant attracted, and that determines how far the seed is dispersed. Hamersma et al. (2022) describe seeds of the crotonoid Belenocarpa tertiara from Peruvian deposits 30-28.5 Ma with a well-developed elaiosome, the whole seed being ca 2 cm long; this may be an early example of myrmecochory, although the ants would have had to be rather robust... Plant-Animal Interactions. Caterpillars of nymphalid butterflies are quite common on Euphorbiaceae (Ehrlich & Raven 1964), e.g. caterpillars of the ca 340 species of Biblidinae are found on Dalechampia and Tragia (Euphorbioideae: DeVries 1987; Wahlberg et al. 2009; Nylin et al. 2014). Caterpillars of the spectacular Uraniinae moths are found on Endospermum, Omphalea and Suregada, also Euphorbioideae, throughout the tropics (Lees & Smith 1991); the first two are rather closely related, the position of the last is unclear (Wurdack et al. 2005). It would be interesting in this context to clarify both Euphorbiaceae phylogeny and Uraniinae host plant preferences. In Malesia about 29 species of fast-growing, large-leaved, ant-associated Macaranga are ecological analogues of the New World Cecropia (Urticaceae); for a phylogeny, see Bänfer et al. (2006) and also Blattner et al. (2001) and Davies et al. (2001). Food bodies (Beccarian bodies) and extra-floral nectaries provide food for the ants (Camponotus and especially Crematogaster spp.) that have an obligate association with the plants, living in their stems which are either hollowed out by the ant or become hollow as the stem ages; myrmecophytism seems to have evolved more than once here (Hatada et al. 2001 for references; Davies et al. 2001; Feldhaar 2003a, b). Indeed, recent work (Dixit & Guicking 2024) suggests that that there has been only a single origin of myrmecophytism in Macaranga, 4-6 losses, and myrmecophytism may have been regained once; Borneo is possibly the place of origin of the crown-group; there has not been much change in diversification rates. Myrmecophytes often are in species-poor clades, exceptions including Psychotria s.l., Neonauclea (both Rubiaceae) and the urticaceous Cecropia (Chomicki & Renner 2015). The age of the association of Macaranga with Crematogaster subg. Decacraema ants has been estimated to be less than 7 Ma, and co-speciation of the two partners was suggested (Itino et al. 2001b; see de Vienne et el. 2013), although the aging of the association was rather vague. On the other hand, Ueda et al. (2008) offered an age of 20-16 Ma for the development of some kind of association between ant and plant (see also Chomicki & Renner 2015), the particular association between Crematogaster subgenus Decacrema ants and Coccus scale insects being only 9-7 Ma; Crematogaster species may have replaced each other within an association, or replaced Camponotus ants (Davidson & McKey 1993). Another estimate for the age of the crown node of the myrmecophytic clade is (23.6-)18.1(-16.4) Ma (Dixit & Guicking 2024). As with other cases of myrmecophytism, there is a complex of adaptations in ant-associated species of Macaranga, for instance, whether the stems are waxy, or not (not all insects can run up waxy stems), whether the food bodies on the stipules are exposed or protected, whether or not there are extrafloral nectaries on the lamina margins, and so on, and these are variously correlated (Federle & Rheindt 2005; see also Davidson & McKey 1993). Itino et al. (2001a and references) thought that an association with coccids, which also provide carbohydrates for the ants, represented the original condition for the ant—plant association. Other organisms are involved in this association. These include bacteria that live off the material in colony rubbish dumps (dead ants are not included in this rubbish) in M. bancana and other species of the genus (Maschwitz et al. 2016). These bacteria, and perhaps the fungi also found there, are in turn eaten by thousands of rhabditid nematodes found on the dumps (genera like Diploscapter and Sclerorhabditis, known for their associations with ants, are involved) that may in turn be eaten by the ants, so aiding in the circulation of nutrients. Nematodes (?and bacteria) may move from colony to colony with the queen ants (Maschwitz et al. 2016). Arhopala (lycaenid) caterpillars are also to be found in the domatia, and they eat ant larvae yet are not attacked by the ants, rather, the products of a tentacle-like gland that is extruded by the caterpillar calm the ants (Maschwitz et al. 1984). Davies et al. (2001) suggested that there were extrafloral nectaries on the leaf margins in myrmecophilous species, but on the leaf surface in non-myrmecophilous species. Macaranga tanarius produces nectar from its extrafloral nectaries in response to damage, or more immediately in response to the jasmonic acid that increased when the plant was damaged; this induced nectar flow attracted ants that defended the plant against herbivores (Heil et al. 2001; see also Heil 2015). Two species of Endospermum are also myrmecophytes, and both they and their associated ant, Camponotus quadriceps, are restricted to East Malesia. However, Schaeffer (1971) thought that this was a facultative association - "the trees need the ants as much as a dog its lice" (ibid: p. 173). For more details of this association, see Davidson and McKey (1993). Herbivore activity may result in the induction of extrafloral nectar in a number of taxa (Heil 2015). The lectins mentioned above are probably involved in defence against insects (Peumans & van Damme 1995; Vandenborre et al. 2011). For laticifers and the trenching behaviour of foliovores in Euphorbiaceae, see Dussourd (2016); for latex in general and plant defence, see Ramos et al. (2019) and references.. Extrafloral nectaries are notably common in Euphorbiaceae (Weber & Keeler 2013). Plant-Bacterial/Fungal Associations. Complex maytansinoids, ansamycin antibiotics, that are likely to be synthesized by fungal endophytes or other plant associates, are found in Trewia (Acalyphoideae: Cassady et al. 2004 for references). The pucciniaceous rust fungus Uromyces pisi causes Euphorbia cyparissias to form nectar-producing pseudoflowers that facilitate transmission of the fungus spermatia. Relationships between plant and visitors to the cyathium and to the pseudoflowers - they are yellow (courtesy of the fungus), secrete nectar, etc. - are complex (Pfunder & Roy 2000). Vegetative Variation. Laticifers in Euphorbiaceae are discussed by Dehgan and Craig (1978: Jatropha), Rudall (1987, 1994a), Wiedenhoft et al. (2009) and Demarco et al. (2013: two articulated laticifer systems in Sapium); see also Vitarelli et al. (2015), Prado and Demarco (2018), the cautionary comments in Wurdack et al. (2005), and the difficulty in distinguishing between articulated and non-articulated laticifers in Prado and Demarco (2018) and Ramos et al. (2019). For laticifer type and associated starch grain morphology - the latter is very variable - see Demarco et al. (2013) and references; latex coagulation in Hevea, is discussed i.a. by Bauer et al. (2014). Vitarelli et al. (2015) looked at the considerable variety of foliar secretory structures in Croton and its near relatives. In addition to recording colleters, the paired basal glands on the lamina (the two have some underlying similarities), etc., they suggest relationships between cells in the lamina filled with secretions ("idioblasts") and short secretory trichomes, whose occurence is mutually exclusive, and in both of which the secretion is some kind of lipid. Feio et al. (2016) examine the variety of secretory structures in other species of Croton, i.a. recording colleters from inside the flower. For colleters and nectaries on the leaves of Sapium, see Strozi Alves Meira et al. (2020). Peroideae are variously described as having lysigenous radial canals in the wood, laticiferous cells, or elongated cells with brown contents (Nowicke et al. 1998; Hayden & Hayden 2000). Laticifers have recently been reported from a number of species throughout Acalyphoideae (but alchorneoids were not examined), and there they are multinucleate, elongated cells with dense cytoplasm and are both articulated and branched (Mouzella et al. 2025). Latex may not be abundant, but it is there, and absence of laticifers is certainly not a feature of Acalyphoideae (Mouzella et al. 2025), but is their presence a feature of the family as a whole? For the morphology and mineral composition of the stinging hairs of Cnidoscolus, see Mustafa et al. (2018b). Although rather little is known about colleters in Euphorbiaceae, they are apparently widespread in the family. Details of their position and development varies. Thus in Mabea fistulifera, for example, they are found both on the stipules and lamina margin, their mucilaginous secretions perhaps protecting the leaf from dessication (A. L. Almeida & Paiva 2019; see also Miguel et al. 2023). Distinctive vascular variants - ectopic cambia - are known from Dalechampia alata (Cunha Neto et al. 2023); perhaps unsurprisingly, the genus includes climbers. Economic Importance. Rubber is a polymer of isoprene, = C5 H8/cis-1,4-polyisoprene, and aspects of the latex composition of the rubber tree, Hevea brasiliensis, the only commercial source of natural rubber, are discussed by Bottier (2020). Papers in Vollmann and Rajcan (2009) focus on oils from Ricinus (Acalyphoideae-Acalypheae). The crotonoid Manihot esculenta, cassava (the starch is called tapioca, etc.), is the most important non-grain source when it comes to providing calories for humans - it provides some 2.6% of the total. Genes & Genomes. A genome duplication in Manihot has been dated at (42.1-)40.4(-38.7) Ma (Vanneste et al. 2014a), while Cai et al. (2017/18) detected a duplication in the [Manihot + Hevea] and the Endospermum clades. An event dated to ca 99.4 Ma (the EUMEα event) involved practcally the whole family (?the basal clade: Landis et al. 2018). The diterpenoid-synthesizing genes form a cluster - except in Manihot (King et al. 2014). Hans (1973) lists chromosome numbers. Chemistry, Morphology, etc.. Phorbol esters in the [Crotonoideae + Euphorbioideae] clade in particular are very diverse; for the biosynthesis of these and other diterpenes, see King et al. (2014). Cyanogens can be derived from nicotinic acid or valine/isoleucine (Seigler 1994). Latex and cocarcinogens are both apparently restricted to Euphorbioideae and Crotonoideae. Distinctive fatty acids in the seed oils are quite common in the family (Badami & Patil 1981). For maytansinoids in Trewia (= Mallotus), see Cassady et al. (2004). The thickness of first-order roots in the family ranges from 1009.6 μm in Endospermum chinense to 72.6 μm in Macaranga sampsonii, the extremes of measurements made on 96 species from southern China (Kong et al. 2014). There may be a multi-layered G layer in the tension wood in some species (Ghislain et al. 2016). Prismatic crystals in wood parenchyma and/or ray cells are common, but these also occur in Putranjivaceae and Picrodendraceae, which used to be included in Euphorbiaceae s.l. (Hayden 1994). Van Welzen et al. (2021) discuss stipule-like structures in Ricinocarpeae where stipules proper are generally absent. Claoxylon has distinctively rough leaves when dry because of the styloids in their tissues (Kabouw et al. 2008). Dioecy may be ancestral in Peroideae, at least, but with subsequent reversions to monoecy (Käfer et al. 2014). The perianth in staminate flowers of Alchornea is first evident as a raised annulus (Gama et al. 2019). Thaowetsuwan et al. (2020) examined the variable development of petals in carpellate flowers of Croton; vascularization of the ?staminodial nectary in Croton and its near relatives varies, and a secretory staminodial nectary may be a high-level apomorphy around here (De-Paula et al. 2011). It can be difficult to understand possible homologies of floral structures in Astraea (c.f. De-Paula et al. 2011; Gagliardi et al. 2017). Prenner et al. (2008a) described the development of the distinctive androecium of Ricinus (Acalyphoideae-Acalypheae) with its highly branched stamens; they thought that these were not cauline as had been suggested, rather, they thought of the androecial unit as being an "extended fascicle" (ibid.; p. 739); each stamen may be a pair of largely separated thecae that have rotated 90o and so face apically, and in between them there is a protrusion that represents the connective. On the other hand, Claβen-Bockhoff and Frankenhäser (2020) prefer to think of each "stamen" as a unistaminate flower subtended by a bract and produced by a floral unit meristem that has divided many times. The tapetum in some species of Euphorbia subgenus Esula is amoeboid (Anisimova 2019). There is extensive ovular variation, but sampling is too poor to know how much of it might relate to phylogeny. Some Euphorbiaceae are reported to have two vascular traces supplying each ovule (Venkata Rao & Ramalkshmi 1967). Johri and Kapil (1953) noted that the vascular tissue in ovules of Acalypha indica proceeded one third the way up the nucellus. The projecting nucellus, a.k.a. the nucellar beak, in Codiaeum variegatum, at least, seems to result from periclinal divisions of the epidermal layer of the nucellus, that is, it is a nucellar cap, although in Euphorbia less striking projections seem to be the result of divisions of the underlying nucellar cells (Bor & Kapil 1975 and references) - but even here they can be quite conspicuous, and may be some kind of nucellar cap (see images in Titova & Nyukalova 2021). Gama et al. (2019) describe the placental obturator of Alchornea sidifolia as being long, with trichomes making contact with the nucellar beak; there are hairs on the inside of the loculus. Mennega (1990) suggested that the subdermal initiation of the inner integument separated Euphorbiaceae from other families (see also Bor & Bouman 1974; Bouman 1984). Vinogradova (2022) mentions multicellular archesporia in some species of Euphorbia. Catechyl lignins have been found in appreciable amounts in the seed coat of the few Crotonoideae and Acalyphoideae studied, but occur in at most low amounts in seeds of Euphorbia (Tobimatsu et al. 2013). The exotegmen of some Peroideae can look U-shaped in transverse section (see illustrations in Tokuoka & Tobe 2003); the exotegmen of Pogonophora is like that of other Euphorbiaceae, while the exotegmen of, say, Pera itself is barely distinct from the rest of the seed coat, and the seed there is exotestal (Franca & De-Paula 2017). The seed coat anatomy of Trigonopleura is unknown. Van Welzen (1994) described Neoscortechinia (Cheilosoideae) as having a thin, red aril, but no aril was mentioned by Tokuoka and Tobe (2003) or Tokuoka (2007). The chalazal endosperm haustorium in Croton can reach 1,000 μm or so in length (Venkateswarlu & Rao 1963). For general information on Euphorbiaceae (but note that references to Acalyphoideae in older literature may include a multitude of sins), see Webster (1967, 1984, 1994a, b [also other papers in Ann. Missouri Bot. Gard. 81. 1994], 2013), Radcliffe-Smith (and Esser, 2001: generic descriptions, etc.), Esser (2001), Eggli (2002: succulent species), Cheek et al. (2016: Crotonoideae), Wurdack and Farfan-Rios (2017: Hippomaneae s.l.), de Sá-Cordeiro et al. (2021: Tragiinae), van Welzen et al. (2021: Ricinocarpeae) and Bruyns (2022: Euphorbia); Hegnauer (1966, 1989), Evans and Taylor (1983: phorbol esters), Jury et al. (1987) and Beutler et al. (1989, 1996) all discuss chemistry. See also Hayden and Hayden (2000: wood anatomy, esp. of Acalyphoideae), Westra and Koek-Noorman (2004: wood end-grain), Mennega (2005: wood anatomy of Euphorbioideae), Cardiel et al. (2020: crystals), Cervantes et al. (2009: leaf anatomy of some Acalyphoideae), Fiser Pecnikar et al. (2012: leaf anatomy of Mallotus and relatives), Rios et al. (2020: leaf teeth and colleters) and Maity (2014: split laterals in Mallotus). See Merino Sutter and Endress (1995: floral morphology), De-Paula and Sajo (2011: anthers and ovules in Croton), for pollen morphology of the family (s.l.), see Punt (1972), for that of Crotonoideae, see Nowicke (1994b), Lobreau-Callen et al. (2000), R.-Y. Yu et al. (2018: Trigonostemon and relatives) and de Souza et al. (2020: Croton), for that of Acalyphoideae s.l., see Nowicke et al. (1998) and Nowicke and Takahashi (2002) and references, for that of Acalypha itself, see Sagun et al. (2006), for that of Euphorbioideae, see Park and Lee (2013: Pimeleodendron, etc., distinct), Suárez-Cerbera et al. (2001: Plukenetieae and Euphorbieae), Sakugawa et al. (2021: Hippomaneae) and also Matomoro-Vidal et al. (2015: function), for embryology, ovules and seeds, see Schweiger (1905), Landes (1946), Singh (1962), Tokuoka and Tobe (1993) and Kapil and Bhatnagar (1994), all general, in Crotonoideae, see Rao (1976) and Tokuoka and Tobe (1998), in Euphorbioideae, see Pammel (1892), Venkateswarlu and Rao (1975), Bor and Bouman (1975), Tokuoka and Tobe 2002), and Vinogradova (2017), in Acalyphoideae, see Kapil (1960), Nair and Abraham (1963), Huner (1991), and Tokuoka and Tobe (2003), Gagliardi et al. (2012, 2013: fruit and seed), Titova et al. (2018: seed development in Euphorbia), and Singh (1969) and Stuppy (1996), both seed anatomy. Phylogeny. Peroideae are sister to other Euphorbiaceae, while Neoscortechinia and Cheilosa are strongly supported as being sister to the remainder (Wurdack et al. 2005; Tokuoka 2007; Xi et al. 2012b). The quotation, "There is no support for a monophyletic Crotonoideae s.l.. Instead, there are four distinct lineages (Adenoclineae s.l., Gelonieae, and articulated and inaperturate crotonoids)" (Wurdack et al. 2005: p. 1413), sums up a major problem along the spine of the family. Tokuoka (2007) also found that Adenoclineae and Gelonieae, with a thinner outer integument and unvascularized testa and tegmen, formed a paraphyletic grade at the base of the family (minus Cheilosioideae), albeit this topology had little support. Crotonoideae were monophyletic, again with little support, Acalyphoideae and Euphorbioideae were monophyletic, but with more support (Tokuoka 2007). Indeed, Tokuoka (2007) found that these last two subfamilies grouped together, as did Matomoro-Vidal et al. (2022), rather than Euphorbioideae grouping with Crotonoideae, as in Wurdack et al. (2005), but in general support was poor; perhaps the differences are a sampling issue (Matomoro-Vidal et al. 2022). For other details of relationships, see Sun et al. (2016). Tokuoka and Tobe (2006) question the inclusion of Pogonophora in Peroideae; indeed, if it is to be included and is sister to other members of the subfamily, then mapping seed coat evolution on the tree does become a bit tricky (it has an exotegmic seed like that of other Euphorbiaceae). M. Sun et al. (2016) also found Pogonophora to be sister to the rest of the subfamily - [Pogonophora [Clutia [Pera ...]]]. There are number of distinctive features in Crotonoideae as broadly construed, but there is as yet no strong evidence that the subfamily is monophyletic (see the C1-5 clades in the tree above, the C1-2 clades are the same as in Wurdack et al. (2005); for the C2 clade, see also Cheek et al. (2016). Many features in the subfamilial characterization above are synapomorphies either for individual clades or groups within them; for instance, the trnL-F spacer deletion is a feature of part of Crotonoideae s. str.. Crotonoideae s. str., the C1-2 clades, may expand from this minimalist circumscription as details of phylogenetic relationships are clarified, and for the most part they have petals, distinctive inaperturate pollen with supratectal processes ("crotonoid pollen"), and the tegmen is usually vascularized. Crotoneae. The large genus Croton (a C1 crotonoid) has been studied by Berry and his collaborators (see Berry et al. 2005; van Ee et al. 2008, 2011, 2015; van Ee & Berry 2009; Riina et al. 2009, 2010; Caruzo et al. 2011; Arévalo et al. 2017a; Riina et al. 2021: section Geiseleria for phylogenies, dates, biogeography, and more). O. L. M. Silva et al. (2020) noted that analyses of nuclear and plastid genes resulted in different topologies here; Sandwithia and Sagotia are either successively sister to the rest of Crotoneae, or the two form a clade. Pinto-Silva et al. (2023) looked at variation in hair morphology in Brazilian species of Croton in particular; only 6/24 sections they examined showed no variation, and the "glandular trichomes" previously reported may be colleters, "papillate trichomes", idioblasts. See also Silva et al. (2020) for relationships in Astraea (Manihoteae), while Maya-Lastra and Steinmann (2019) looked at the phylogeny of Cnidoscolus with its stinging hairs. Trigonostemoneae: R.-Y. Yu et al. (2018) at relationships in Trigonostemon itself, where there is a correlation between pollen morphology and phylogeny, and Yu and van Welzen (2019) those in Trigonostemon and Dimorphocalyx. Van Welzen et al. (2021) examined relationships in Ricinocarpeae, those along the spine of the C2 crotonoids, to which Ricinocarpeae belong, were for the most part poorly supported. Although Acalyphoideae in the old sense are paraphyletic, the great bulk of the subfamily is included in a strongly-supported clade, Acalyphoideae s. str., and Cervantes et al. (2016) found that Erismanthus was sister to all other Acalyphoideae that they examined. Several tribes as currently delimited are para- or polyphyletic, perhaps most notably Acalypheae, members of which are in six clades in the tree recovered by Cervantes et al. (2016). Levin et al. (2022: just over 15% species sampled, 2 plastid genes plus nuclear ITS) looked at relationships in Acalypha and found that just about all the supraspecific taxa in the previous classification of Pax and Hoffmann were polyphyletic. Wurdack et al. (2005) discuss groupings in the subfamily in some detail. Slik et al. (2001: morphology), Sierra et al. (2006, 2010 and references: the latter including both qualitative and quantitative morphological variation), Kulju et al. (2007a, b) and van Welzen et al. (2014a; see also Yamasaki et al. 2015) evaluate the phylogeny of the Macaranga-Mallotus complex ("Acalypheae"); there are three main clades, and in Mallotus s. str. in particular some small, segregate genera are embedded. Cardinal-McTeague and Gillespie (2016) looked at relationships within the monophyletic Plukenetieae (not so much at Dalechampia, for which see Armbruster et al. 2013a, esp. 2009b), where Tragia in particular was found to be very para/polyphyletic; there was not that much correlation between phylogeny and the quite extensive variation in pollen morphology. Zhou et al. (2017) looked at relationships within Epiprineae. Within Euphorbioideae, usually well supported, M. Sun et al. (2016) obtained the relationships [Euphorbia [Hura ...] [[Actinostemon + Maprounea] ...]]. Stomatocalyceae, which include Pimelodendron and Nealchornea, may be sister to the rest of the subfamily (e.g. Wurdack et al. 2005); they often have extrorse anthers and the testa is at least sometimes vascularized. For more details of relationships here, see Wurdack et al. (2005) and M. Sun et al. (2016). Much work has been carried out on relationships within Euphorbia itself over the last few years. For a phylogeny of Euphorbia, see Bruyns et al. (2006), Park and Jansen (2007), Zimmermann et al. (2010) and Wurdack et al. (2011). The last three authors found that subgenus Esula was sister to the other subgenera, although not always with very strong support - relationships are [Esula [Rhizanthium (= Athymalus) [Euphorbia + Chamaesyce]]] (see also the extensive study by Bruyns et al. 2011; Horn et al. 2012). Sampling was initially poor, however, this has steadily improved. Some characters are particularly common/important within the subgenera, although there is very extensive homoplasy. A few of the major characters are [Esula (annuals, cyathium with 4 glands) [Athymalus (cyathium with 5 glands, inflorescences sometimes lateral, plants succulents) [Euphorbia (inflorescences often lateral, plants succulents) + Chamaesyce (annuals, C4 photosynthesis, leaves often opposite)]]] (Horn et al. 2012, q.v. for references and many more details). For relationships in the leafy Euphorbia subgenus Esula in particular, with a considerable number of north temperate taxa, see Frajman and Schönswetter (2011) and Riina et al. (2013); the position of E. lathyris is unclear, but it may be sister to the rest of the subgenus. See also the summary in Geltman et al. (2011) and the study or relationships in the New World section Tithymalus by Peirson et al. (2014). For relationships in the Old World subgenus Athymalus, in which the only Malagasy species, E. antso, appears to be sister to all other species, see Morawetz and Riina (2011) and Peirson et al. (2013). For relationships in the mostly New World subgenus Chamaesyce, see Y. Yang and Berry (2007, 2011) and Yang et al. (2012), for those in Macaronesian taxa, see Molero et al. (2002), and for those in the large subgenus Euphorbia, see Dorsey et al. (2011, 2013). See also above for cyathia, growth forms, succulence, thorns and spines, and photosynthetic pathways. Classification. For a comprehensive checklist and bibliography of the family, now dated, see Govaerts et al. (2000). Wurdack et al. (2005) discuss morphology, groupings and relationships in Euphorbiaceae in considerable detail. Cardinal-McTeague and Gillespie (2016) discuss generic limits in Acalyphoideae-Plukenetieae, Jestrow et al. (2010) those in the -Adelieae, van Welzen et al. (2021) discuss generic limits in Ricinocarpeae, and Kulju et al. (2007a) and Sierra et al. (2007) those in the Macaranga-Mallotus area; for an outline of the classification of the speciose Macaranga, see Whitmore (2008). For a sectional classification of Croton, see van Ee et al. (2011, 2015: Australian sections), for an infrageneric classification of Trigonostemon, see R.-Y. Yu et al. (2019), and for that of Cnidoscolus, see Maya-Lastra and Steinmann (2019). Levin et al. (2022) recognized four subgenera in Acalypha; the great bulk of the genus was in subgenus Acalypha, within which there were five groups, of which the core Acalypha clade included about 2/3 the species examined. For information on Euphorbia, see EuphORBia (Esser et al. 2009) and there is a developing inventory, etc., of the genus, at Tolkin (Riina & Berry 2012), but not if you use Internet Explorer only... Euphorbia is best broadly circumscribed, so including the whole of the Euphorbiinae of Webster (1994b), i.e. genera like Chamaesyce, Pedilanthus, Monadenium, Synadenium, etc., and it is well characterized by its cyathium (e.g. Bruyns 2010 and references). Four subgenera are recognized, although they have no synapomorphies - Y. Yang et al. (2012) provide a classification of Euphorbia subgenus Chamaesyce, Riina et al. (2013) that of subgenus Esula, Dorsey et al. (2013) that of subgenus Euphorbia, and Peirson et al. (2013) that of subgenus Athymalus. For comments on Euphorbia and its classification, see Muñoz-Rodríguez et al. (2023). Previous Relationships. Although Airy Shaw (1976) recognised Peraceae as separate from Euphorbiaceae, his Peraceae included only Pera, a genus long considered to be very distinctive within Euphorbiaceae, even if not separated from it. Esser (2003a) drew attention to the distinctiveness of the whole group.
Botanical Trivia. Croton has got little to do with the cultivated croton, which is Codiaeum.
Acalypha indica "is widely known for its effect on domestic cats, which react very strongly and favorably to the root of the plant". (Wikipedia viii.2022).
Thanks. I am grateful to Hajo Esser for comments and to Ken Wurdack for help with the generic synonymy.
[[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]: stomata paracytic; ovules 2/carpel.
Age. Estimates of the age of this node are (108.6-)102.5(-94.8) Ma (Xi et al. 2012b; Table S7).
Phylogeny. Although this clade was found by M. Sun et al. (2016), it was separate from other members of Clade 1 and relationships within it were largely only weakly supported.
[Phyllanthaceae + Picrodendraceae] / Phyllanthoids: lamina margins entire; flowers small; anthers extrorse (introrse); micropyle bistomal, parietal tissue 10 or more cells across, nucellar beak +; fruit with outer layer of the pericarp often separating from the inner woody layer, valves falling off, central column persistent; endosperm copious; x = 13.
Age. Estimates for the age of this node are (101.6-)94(-86.5) Ma (Xi et al. 2012b; Table S7), ca 94.8 Ma (Tank et al. 2015: table S2), or the Cretaceous-Albian 111-100 Ma or a little later - (114.0-)108.1(-105.8)/(101.9-)97.1(-95.6) Ma (Davis et al. 2005a).
Evolution: Divergence & Distribution. Merino Sutter et al. (2006) suggest additional possible similarities between the two families.
Chemistry, Morphology, etc.. Pre-2005 references to Euphorbiaceae may contain information about this clade. For general information, see Webster (1994a, b, 2013), see also Radcliffe-Smith and Esser (2001: description of genera), Hegnauer (1966, 1989) and Jury et al. (1987), all chemistry, Punt (1972) and Köhler (1965), both pollen, and Schweiger (1905: ovules and seeds).
Phylogeny. The clade [Phyllanthaceae + Picrodendraceae] had only very slight support in a rbcL analysis, although the two families have morphological similarities (Wurdack et al. 2004). Support is stronger (75% bootstrap, 1.0 posterior probablility) in a recent 4-gene analysis (Davis et al. 2005a; see also Tokuoka & Tobe 2006; Korotkova et al. 2009) and even stronger in Soltis et al. (2011). Thus the two are probably sister taxa.
PHYLLANTHACEAE Martynov, ±= the old Euphorbiaceae-Phyllanthoideae Beilschmied - Back to Malpighiales
(Plants Al accumulators); cyanogenesis via the tyrosine pathway, tropane, piperidine and pyrrolizidine alkaloids, cucurbitacins [triterpenes], nonhydrolysable tannins [geraniin] +, ellagic acid 0; cork?; (axial parenchyma 0); (mucilage cells [epidermis] +); (stomata anisocytic); lamina vernation involute or conduplicate, glands usu. 0; plant monoecious/dioecious; K 2-8(-12), often biseriate, imbricate, often basally connate, C (0, 3-)5(-9), (small); nectary extrastaminal, ± annular and/or variously lobed, (central); staminate flowers: A 2-35; pollen surface reticulate; (pistillode +); carpellate flowers: (staminodes +); G (1)[2-5(-15)], style branches spreading, stigmas with adaxial furrow, smooth, wet; ovules with outer integument 2-many cells across, inner integument 2-3(-10) cells across, both integuments initiated from dermal cells; fruit a septicidal capsule/schizocarp; seeds large, often 1/carpel, aril/caruncle 0; (vascular bundles in testa), tegmen 2-5(-20) cells thick, exotegmic cells ribbon-like; (endosperm 0), (embryo chlorophyllous), cotyledons thin and flat; n = (6-9, 11, 14), x = 7 (?6), nuclear genome [1C] (936-)1278(-1809) Mb/(0.023-)1.133(-55.408) pg.
66[list, to tribes]/2,330 - two subfamilies, ten tribes below. Pantropical, but esp. Malesia, some temperate. Map: from Webster (1970, 1984, 1994a, etc.), Wickens (1976), Frankenberg and Klaus (1980), FloraBase 1 (2011) and Trop. Afr. Fl. Pl. Ecol. Distr. vol. 2 (2006). [Photo - Flower.]
Age. Estimates for the crown group age of the family are (93.7-)81.2(-58.8) Ma (Xi et al. 2012b: Table S7).
Fruits, Phyllanthocarpum singpurensis, with palisade exotegmen and two layers of palisade sclereids in the pericarp, that were found in deposits some 66 Ma have recently been described from the Deccan Traps in India (Kapgate et al. 2017).
1. Phyllanthoideae Beilschmied
(Plants monopodial), growth continuous; (nodes 1:1); (plant dioecious); inflorescence fasciculate; (P/K + C with a single trace); staminate flowers: A free to connate; pollen (to 16-colporate), (colpi diploporate), (pantoporate), (inaperturate); carpelate flowers: G [2-6(-15)], (style single, columnar, apex ± expanded); fruit (indehiscent), wall with veins, (expanding early).
45/1,680. Tropical to Temperate.
1A. Bridelieae Müller Argoviensis
Shrubs to trees; pits vestured; plant monoecious/dioecious; (pedicels articulated); staminate flowers: (androgynophore + - Cleistanthus); pollen (surface spiny - Croizatia); pistillode +; G [2-3(-4)]; fruit a drupe or capsule; endosperm +/-, cotyledons (plicate/fleshy); n = 11, 13.
13/: Cleistanthus (130), Bridelia (50). Tropical.
Age. Early Palaeocene wood fossils ca 65 Ma of Bischofinium deccanii from the Deccan Traps are perhaps similar to Bridelia (Wheeler et al. 2017).
Lachnostylis has a stem age - Phyllanthus sister - of as much as 97-75 Ma (Warren & Hawkins 2006) - check.
[Phyllantheae [Wielandieae + Poranthereae]]: ?
1B. Phyllantheae Dumortier
(Annual) or perennial herbs or shrubs (small trees); vessel elements with simple ["Glochidion"] perforation plates; (plagiotropic axes phyllodinous); leaves on orthotropic axes often reduced, spiral, on plagiotropic axes two-ranked (reduced); plant monoecious/dioecious; inflorescences (terminal), on plagiotropic axes; K (connate), C usu. 0; nectary annular / of three or six separate glands / (0); staminate flowers: A 1-7(+), anther ± connate or not, (thecae separate), dehiscence (transverse to oblique); carpellate flowers: G [3(-15)], (styles connate, with central cavity), (stigma papillae 0); ovules usu. hemitropous, (basal); fruit (berry); (sarcotesta +), (exotegmic cells short-palisade); endosperm 0; n = 11.
18/1,340: Glochidion (350), Phyllanthus (213), Moeroris (199), Dendrophyllanthus (161), Breynia (89), Nymphanthus (86), Cicca (45), Emblica (45), Cathetus (40). Tropical, some Temperate.
Age. Phyllantheae (Flueggea, Glochidion, etc.) are dated to (94.2-)51.8(–48.6) Ma (van Welzen et al. 2015).
[Wielandieae + Poranthereae]: ?
1C. Wielandieae Hurusawa
Shrubs to trees; (petiole pulvinate); plant monoecious/dioecious; (C 0); endosperm +/-.
6/80: Actephila (35), Savia (25). Rather scattered, tropics and some warm temperate, inc. S. China, not W. Africa or Australia.
1D. Poranthereae Grüning —— Synonymy: Porantheraceae Hurusawa
Herbs to trees; plant usu. monoecious; (pedicels articulated); endosperm usu. +, cotyledons (linear/plicate).
8/ . ± Worldwide, not polar.
2. Antidesmatoideae Hurusawa
Plant growth rhythmic; epidermal cells tanniniferous; leaves spiral; plant often dioecious; inflorescence with axis; pedicels not articulated; C often 0; staminate flowers: tapetal cells 2-3-nucleate; carpelate flowers: fruit often indehiscent/tardily dehiscent.
21/450. Tropics and subtropics.
2A. Bischofieae Hurosawa - Bischofia Blume —— Synonymy: Bischofiaceae Airy Shaw
Trees; sieve tubes with non-dispersive protein bodies; leaves trifoliolate, margins toothed, teeth deciduous; plant usu. dioecious; ; disc 0; staminate flowers: carpellode?; carpellate flowers: staminodes +, minute; micropyle bitegmic, parietal tissue 10-15 cells across, nucellar cap +, nucellar beak ± 0, obturator +, hypostase +; fruit capsule/schizocarp; exotesta brown, membraneous, tanniniferous, exotegmen as radially elongated sclereids near micropyle, otherwise much enlarged, with ribbon-like thickenings, endotegmen tanniniferous; endosperm +, suspensor spherical, multinucleate.
1/2. China and India to Australia and W. Pacific islands.
[Uapaceae, Spondiantheae [Scepeae [Jablonskiieae + Antidesmateae]]]: ?
2B. Uapaceae Hutchison - Uapaca Baillon —— Synonymy: Uapacaceae Airy Shaw
Shrubs to trees, stilt roots common, ectomycorrhizal; exudate resinous; leaves crowded at apex of branch; stipules usu. deciduous; plant dioecious; disc 0; endosperm 0.
1/60. Tropical Africa, Madagascar.
2C. Spondiantheae G. L. Webster - Spondianthus preussii Engler
Trees; exudate resinous; leaves crowded at apex of branch; plant dioecious; C usu. +; endosperm +.
1/1. Tropical East Africa.
[Scepeae [Jablonskiieae + Antidesmateae]]: ?
2D. Scepeae Horaninow —— Synonymy: Aporosaceae Planchon, Scepaceae Lindley
Shrubs to trees, Al accumulators [Baccaurea]; vessel elements with scalariform perforation plates; leaves (crowded at apex of branch); plant dioecious, (flowers perfect); (pedicels articulated); (disc 0); (stigma plumose); (coloured sarcotesta); endosperm +.
8/: Aporosa (90), Baccaurea (50). Pantropical, not Australia.
[Jablonskiieae + Antidesmateae]: ?
2E. Jablonskieae Petra Hoffmann
Shrub to tree; plant monoecious/dioecious; seed sarcotesta +, thin/minutely carunculate; endosperm +.
2/2. Northern South America.
2F. Antidesmateae Bentham & J. D. Hooker —— Synonymy: Antidesmataceae Loudon, Hymenocardiaceae Airy Shaw, Stilaginaceae C. Agardh
(Perennial herbs), shrubs to trees; (petioles pulvinate); plant often dioecious; anther thecae usu. separated; endosperm +/(0).
8/: Antidesma (150). Pantropical.
N.B. In the discussion below on the evolution of morphology and biology of Phyllanthus and its relatives, including the distinctive growth forms of the plants and the remaerkable plant-pollinator interactions, the names will need to be reviewed to fit the new generic nomenclature of Phyllantheae of Bouman et al. (2022).
Evolution: Divergence & Distribution. Shukla et al. (2016) discuss fossils dating to the Cretaceous that may belong around here.
Lachnostylis (Phyllanthoideae-Bridelieae), a small Cape genus, seems to be a very old (up to ca 97 Ma) relict element there (Warren & Hawkins 2006), while the distribution of the Baccaurea group (Antidesmatoideae) may initially have been affected by drift events ca 80 Ma (Haegens 2000); these clade ages should be re-examined. For the possible (post-)Miocene E → W dispersal of Bridelia across the Indian ocean, see Li et al. (2009).
Van Welzen et al. (2015) discussed the diversification of Breynia, with around 85 species mostly in continental Southeast Asia, suggesting that it may be linked with pollination mode (Epicephala moths - see below) and limestone habitats. There are around 106 endemic species of Phyllanthus on New Caledonia alone, many restricted to serpentine, and they represent one of the major radiations on the island (Kathriarchchi et al. 2006; Kawakita & Kato 2017a; c.f. Barrabé et al. 2014). The start of this radiation has been dated to around (27.2-)20(-17.7) Ma, and long distance dispersal was probably responsible for the arrival of Phyllanthus (Nattier et al. 2017). Glochidion s. str. has speciated on the high islands of southeastern Polynesia, probably mostly within the last 5 Ma, and it represents an important angiosperm radiation there (Hembry et al. 2013). The ca 75% level of endemism of Phyllanthus in the Caribbean (of the 80+ species growing there, largely members of a single clade) is particularly high (Falcón-Hidalgo et al. 2020, see also Webster 1956). A clade of three Caribbean species was nestled within the African subgenus Afroswartiani, while subgenus Xylophylla is New World in origin. The considerable speciation in Cuba may in part reflect the archipelago-like outcrops of serpentine - i.e. individually isolated - on which the plants often grow (Jestrow et al. 2012).
Ecology & Physiology. There are hyperaccumulators of various metals in the family. Phyllanthus in particular is often to be found growing on serpentine or limestone, the former being the habitat of many of the endemic Phyllanthus on New Caledonia (Kawakita & Kato 2017a; Gei et al. 2020). 18/37 species (and one hybrid) of Cuban Phyllanthus growing on serpentine soils accumulate nickel which is especially abundant in the plants' exudate (Berazaín et al. 2007), while in New Caledonia 14/76 species tested were accumulators (Reeves et al. 1996; Brooks 1998). Phyllanthus balgooyi, growing on ultramafic rock in Sabah, Malaysia, has around 16% by weight of nickel in its phloem sap, the second highest concentration known in angiosperms; normally nickel is found in the epidermis when it accumulates (Mesjasz-Przbylowicz et al. 2016).
The ectomycorrhizal (ECM) Uapaca (Antidesmatoideae) can locally dominate the vegetation in eastern Madagascar, while on the African mainland it is often a component of Detarioideae-dominated woodlands and savannas (White 1983). It is an early successional species in the latter habitat, and acts almost as a nurse tree to other ECM species, all having similar mycorrhizal fungi (Högberg & Piearce 1986; see also Ducousso et al. 2008; Brundrett 2017a; Tedersoo 2017b; Tedersoo & Brundrett 2017 for literature, dates, etc. - perhaps also the Antipodean Poranthera). Uapaca paludosa is one of the four common species mentioned growing in the ca 145,500 km2 of peat in the Cuvette Centrale in the Congo; the three other species are not ECM plants (Dargie et al. 2017).
Pollination Biology. A fascinating pollination mutualism involves females of perhaps 500 species of the moth Epicephala (Gracillariidae-Omixolinae - see Kawahara et al. 2016) and some 500+ woody species in Breynia sect. Breynia, Glochidion, and at least three more clades that are all part of Phyllanthus s.l. (Kato et al. 2003; Kawakita & Kato 2004a, b, 2006; Kawakita et al. 2004; Svensson et al. 2010; papers in Kato & Kawakita 2017a; Falcón-Hidalgo et al. 2020). This is the second largest brood-site pollination mutualism system (Ficus is the largest), although here the association seems to have evolved seven or more times, not once; overall, somewhat over 40% of the taxa in Phyllanthus s.l. are involved (Kawakita & Kato 2009; S.-X. Luo et al. 2017; Kato & Kawakita 2017a; Kawakita et al. 2019). The focus in van Welzen et al. (2023) is on the floral syndrome associated with moth pollination. In plants involved in this mutualism, the perianth and stamens are often fused or are simply erect, forming a tube, and the styles have independently often become reduced and connate, and may form a little cup into which pollen is deposited by the moth, the nectaries are reduced (the moths do not visit the plants for nectar), and the stigmatic papillae have been lost, however, like any pollination syndrome, the match between pollinator and floral morphology is decidedly less than perfect. Female moths have hair-shaped sensilla located on their proboscides, perhaps an adaptation for active pollen collection from male flowers and pollen deposition on female flowers; both male Epicephala and other gracillarids in general lack these hairs (see also Hao et al. 2025). The moth oviposits down the style, the eggs being laid just above the ovules (Kato & Kawakita 2017b, c), although other species lay eggs in different places (e.g. see Kawakita et al. 2015).
Flowers in which the moth lays eggs may lose all their seeds to the growing caterpillars, but the moth pollinates more flowers than those on which it oviposits - furthermore, Gato et al. (2010) found that Glochidion acuminata flowers that had a high egg load, e.g., if a moth had laid eggs in flowers in which there were already eggs, or where there had been extensive ovule damage, were aborted (see also Chheang et al. 2021). The flowers are scented, and male and female flowers may produce different scents, and female flowers may even change their scent after pollination, perhaps to avoid more visits of the moth and the consequent loss of all their seeds to caterpillars; species of Phyllanthus that are pollinated by Epicephala lack nectaries (Svensson et al. 2010; Svensson & Okamoto 2015; see also Okamoto 2017; Bouman et al. 2020 - but see above). The pattern of flowering, fruiting and caterpillar growth in species pairs like G. lanceolarium/E. lanceolaria is complex. Here the eggs of the moth do not hatch for several months after they are laid, the caterpillars eat the subripe seeds and pupate inside the capsule, and the moths hatch about three weeks before the capsule finally opens and releases them, all in all a remarkable system (S.-X. Luo et al. 2017a); Henderson et al. (2019) found the same behaviour in the Australian G. ferdinandi.
Kathriarachchi et al. (2006) had suggested that Phyllanthus colonized the New World three times, and they also found Epicephala larvae and cocoons on herbarium specimens of New World species of Phyllanthus s.l.. Interestingly, the first New World species of Epicephala were described in Kawakita et al. as recently as 2019. The mutualism may well also occur in some Guayanan taxa placed in Phyllanthus that have the same distinctive cup- or tube-shaped stigma morphology as found in the Old World taxa (see also Kawakita & Kato 2017b), although pollination there is as yet unknown. However, the relationships between these New World Phyllanthus and their potential pollinators are complex (see also van Welzen et al. 2023 for the floral morphologies of plants perhaps pollinated by moths, and those of plants with other pollinators). Not all the species of Epicephala examined had hairs on the proboscides of the female moths, there may be other floral visitors (but are they pollinators?), the flowers have well-developed nectaries and the female flowers have flaring stigmas, the moths may lay eggs in young fruits and show no pollination behaviour, even if they have accumulated pollen on their proboscides; nevertheless, Kawakita et al. (2019) did observe genuine pollination behaviour here, and it was quite extensive in some species. (Note that Epicephala is very poorly known, with fewer than 70 of probably several hundred species having been described - see Kawakita & Kato 2017b.) Overall, the most extreme adaptations for moth pollination seem to be in Old World taxa (van Welzen et al. 2023).
There has been discussion as to whether coevolution is involved in this pollination system, and clearly suggestions as to when plant and pollinator evolved presents problems and there may not be congruent patterns of speciation (see Hao et al. 2025). However, Hao et al. (2023), looking at 16 populations of Kirganelia microcarpa and three related species of Epicephala (one of these is a cheater) in China found some agreement between variation in features of the moths that enable pollination and in the K. microcarpa populations they were pollinating, so perhaps there is at least local coevolution - although the overall variation pattern is complex.
There is more discussion on insect/plant relationships and pollination immediately below; see also Asparagaceae-Agavoideae, Ranunculaceae, Saxifragaceae-Lithophragma, Moraceae-Ficus and Caryophyllaceae for similar pollinator-seed eater interactions; Hembry and Althoff (2016) and Kawakita and Kato (2017f) review diversification and coevolution in these systems.
In another variant of plant-pollinator mutualisms, adults of two species of Deltophora moths (Gelechiidae) were found to be the only pollinators of two quite unrelated Chinese species of Phyllanthus, and adults were found only when their host plants were in flower; their larvae eat the foliage of these plants (Luo et al. 2011: part mismatch between plant and moth life cycles?). Finally, Kawakita et al. (2022) found that three species of Phyllanthus s.l., now in Nympanthus, Kirganelia and "P." baladensis, all with dark red flowers, rather open corollas, nectar, etc., were pollinated by gall midges (for the gall midge pollination syndrome, see also Mochizuki & Kawakita 2017), the female midges laid eggs in the staminate flowers, galling them (but not in Nympanthus), the midge larvae then eating the gall, and the carpellate flowers were pollinated as the midges moved around the flower. A number of other species of Phyllanthus s.l. have similar dark red flowers, while galled staminate flowers have been observed on some New Caledonian species (Kawakita et al. 2022); gall midge pollination of one sort or another may be quite common in Phyllantheae.
For records of other pollinators in Phyllantheae, see e.g. Kawakita and Kato (2017b). Flowers using these other pollinators have different morphologies, e.g., the stigmas are bilobed and spreading, petals/tepals may be brightly coloured, and so on. Uapaca, dioecious, has pseudanthia (Baczynski & Claßen-Bockhoff 2023).
Plant-Animal Interactions. Aspects of the mutualistic plant pollination/animal larval support associations between the gracillariid Epicephala moths and Phyllanthus/Glochidion/Breynia are discussed immediately above. These associations seem to have evolved may be nine times, perhaps some time after the initial divergence of the clades in which the mutualisms are now found - 55.2-33.4 Ma (plant) versus 35-20 Ma (moth) - and it has also been lost (Kawakita & Kato 2009; Kawakita 2010; van Welzen et al. 2023). However, the ca 41 Ma age for the origin of the mutualism in Phyllanthus as suggested by Kawakita and Kato (2017c) is that of the common ancestor of a clade in which there have been three independent origins of the mutualism, so an age of ca 35-23 Ma for the first origin of a mutualism is more likely, more in line with the evolution of mutualist Epicephala 30-20 Ma (ibid.: c.f. p. 148, Fig. 6.6). Ages seem to be in a bit of a muddle (or I am), other estimates of plant ages being only (15.9-)9, 5.6(-1.6) Ma (van Welzen et al. 2015, q.v. for discussion; ?sampling; other ages in Kawakita & Kato 2005).
The association between plant and moth is widespread (Chheang et al. 2021). However, any phylogenetic congruence between the two appears to have broken down, at least in part, in the Pacific islands with two separate clades of Epicephala moths being involved in the pollination of a single (almost) plant clade (Hembry et al. 2013; Hembry 2015, 2017). One does wonder how plant and pollinator are able to move together from island to island in the Pacific, as seems to have happened in some cases (see also Hembry 2017). There are host shifts followed by speciation of the moth, not straight coevolution=cospeciation, and different species of plants can share the same pollinator, or a single species of plant may be pollinated by more than one species of moth (Hembry 2015); for the latter, see Chheang et al. (2021) who noted that different species of moth might pollinate the one species of Glochidion, if at widely separated localities. Two species of Epicephala pollinated two species of Phyllanthus in southern China, and although the species of Phyllanthus may be sister taxa, the moths are not, perhaps diffuse coevolution (J. Zhang et al. 2012), and there are other cases where more than one species of moth pollinates a single species of Phyllanthus, although they may at least lay eggs in different places on the flower (Finch et al. 2018). Overall, there is some signal of cospeciation, although it is difficult to distinguish a cospeciation pattern from that generated by other evolutionary processes (see also Satler et al. 2019), and the whole system, as in other cases of purported cospeciation, is complex (Kawakita & Kato 2017c, e; S.-X. Luo et al. 2017a: Fig. 7). Some other caterpillars that eat Phyllanthus seeds, but where the adult moths themselves (unrelated to Epicephala) are not pollinators, have been described as seed parasites (Finch et al. 2018 and references). For further information, see above.
The larvae of most gracillariid moths are leaf miners (Kawahara et al. 2016). However, in these phyllanthoids they are simple seed predators, while in Taiwan, at least, some mutualist Epicephala have secondarily become gallers on Phyllanthus. In this system, studied by Kawakita et al. (2015), a single species of Phyllanthus (the P. reticulatus complex - its limits are unclear) is associated with six closely related species of Epicephala, three being pollinators and three gallers. The galls either have a tough surface or are inflated, and in both cases the moth larvae inside the galls are protected against the unwelcome attentions of parasitic braconid wasps which lay eggs in the caterpillars - the level of parasitism can be pretty high (Kawakita et al. 2015; Kawakita 2017). In general, non-mutualist Epicephala lack hairs on their proboscides, but hairs may be present and pollination may occur in these galling Epicephala. Interestingly, although the larvae inside the gall with a tough surface are protected against braconids, female eulophids, Aprostocetus (chalcid wasps), can penetrate the gall and parasitize the caterpillar (Kawakita et al. 2015).
Plant-Bacterial/Fungal Associations. The ectomycorrhizal fungi of Uapaca group with those from Fabaceae (Högberg & Piearce 1986; Tedersoo et al. 2014a), perhaps reflecting the fact that the two may grow in the same communities. ECM and arbuscular mycorrhizae may occur together (Ba et al. 2012).
Vegetative Variation. Phyllanthoid branching occurs in many, but not all, species of Phyllanthus s.l. (Kathriarachchi et al. 2006). There seems to have been but a single origin of this feature in Phyllantheae, and although it has been lost several times it has never been regained (Kathriarachchi et al. 2006; Kawakita & Kato 2017a; Falcón-Hidalgo et al. 2020; see also Bouman et al. 2020). In such branching the orthotropic axes have reduced, spirally-arranged leaves and the plagiotropic axes usually have two-ranked, photosynthetic leaves and flowers are borne in the axils of those leaves. The plagiotropic axes are often of more or less limited growth. Those of P. acidus, for example, are short-lived and lack flowers, so being the functional equivalent of compound leaves; the flowers themselves are borne on short branches lacking photosynthetic leaves and which arise from separate axillary buds on the main axes. Some Caribbean and Brazilian species are yet more modified. Thus the plagiotropic lateral branches of P. epiphyllanthus and relatives (section Xylophylla) have cladodes/phylloclades, which may in turn bear 2-ranked cladodes, each with flowers and fruits in two ranks in the axils of much reduced scale-like leaves. Brazilian taxa, in section Choretropsis, may have terete phylloclades (Orlandini et al. 2021). Seedlings in all cases have normal-looking leaves along the main axis.
Genes & Genomes. There may have been genome duplications along the Sauropus (= Breynia) and the Bischofia clades (Cai et al. 2017/18). See Hans (1973) for chromosome numbers.
For the plastomes of Phyllanthus - little variation - see Rehman et al. (2021).
Chemistry, Morphology, etc.. The only record of cocarcinogens is from one species of Antidesma (Beuteler et al. 1989). Wood anatomy is variable - but that of Baccaurea and of Aporosa is very similar (Hayden & Brandt 1984; Jangid & Gupta 2017); variation in wood anatomy has not been integrated with the phylogeny. The nodes are quite often unilacunar (Thakur & Patil 2002).
The inflorescence of Uapaca is a pseudanthium. There has been some discussion over the nature of the perianth, especially in Phyllanthus. Phyllanthus urinaria, for example, has 3-merous flowers with six perianth parts and, in staminate flowers, three connate stamens. The six perianth parts are in two whorls and each part has but a single vascular trace, although members of the inner whorl also have traces purely of phloem which supply the nectaries. Because they have these phloem traces and because the perianth members are in two whorls, Gama et al. (2016a) suggest that P. urinaria has sepals and petals, both with open aestivation. The pollen is very variable in Phyllanthus in particular (Webster 1956; Nowicke 1994; Santiago et al. 2004; Webster & Carpenter 2008; Chen et al. 2009; M.-J. Wu et al. 2016: Malesian spp.; Falcón-Hidalgo et al. 2020). The outer integument is variable in thickness. The fruit type of the ancestor of Phyllanthaceae is unclear (Kathriarachchi et al. 2005). The exotegmen is most often described as being ribbon-like or tracheoidal. However, Hymenocardieae (Didymocistus, Hymenocardium) have a collapsed tracheoidal exotegmen and large tanniniferous endotegmic cells; do they belong here (Tokuoka & Tobe 2001)? - yes, say Wurdack et al. (2004) and Kathriarachchi et al. (2005).
For additional information, see Radcliffe-Smith and Esser (2001) and Webster (2013), both general, also León Enriquez et al. (2008: architectural variation), Mennega (1987: wood anatomy), Westra and Koek-Noorman (2004: wood end-grain), Levin (1986: leaves), Z.-G. Zhang et al. (2012: floral morphology), Schweiger (1905: ovules), Singh (1962), Tokuoka and Tobe (2001) and Bhatnagar et al. (2023: Bischofia), both ovules and seeds, and Gagliardi et al. (2014: seeds and fruits), both of Phyllanthus. For pollen, see Punt (1987), Webster and Carpenter (2002: Phyllanthus) and Sagun and van der Ham (2003: Flueggeinae), also Matomoro-Vidal et al. (2015). For a monograph of Baccaurea and relatives, see Haegens (2000), and of Aporosa, see Schot (2004).
Phylogeny. Wurdack et al. (2004: also morphology), Samuel et al. (2005: two gene analysis) and M. Sun et al. (2016) discuss general phylogenetic relationships. Kathriarachchi et al. (2005: five-genes; see also Hoffmann et al. 2006; Xi et al. 2012b) divide the family into two main clades, Phyllanthoideae - they include Lingelsheimia, sometimes previously associated with Putranjivaceae, and Dicoelia, ditto with Pandaceae, and Antidesmatoideae (see also Xi et al. 2014b). Croizatia has previously been associated with genera included in Picrodendraceae, although it differs in several morphological features; it is here placed in Phyllanthoideae-Bridelieae (see Wurdack et al. 2004; Wurdack 2008).
For phylogenetic relationships in the Phyllanthus area, see Kathriarachchi et al. (2006), Lorence and Wagner (2011), Pruesapan et al. (2008, 2012), Z.-D. Chen et al. (2016) and Kawakita and Kato (2017a); Phyllanthus is paraphyletic, and [Margaritaria + Platycladus] are sister to the rest of the tribe (for the latter position, see also Bouman et al. 2020). Falcón-Hidalgo et al. (2020) with a focus on the Caribbean species, but with quite extensive general sampling, found both rather different immediate relatives of Phyllanthus and also relationships within the genus; Old World clades were basal in Phyllanthus, P. maderaspatensis being sister to the rest. In an extensive study including all subgenera of Phyllanthus, almost three quarters of its sections and a quarter of its species, and based on analyses of variation in both nuclear and chloroplast markers, Bouman et al. (2020) found extensive para- and polyphyly of these infrageneric groupings. Two or more specimens of a number of species were included, and they were often quite distinct in the analyses carried out, and there was also some chloroplast-nuclear conflict... (Bouman et al. 2020). Bouman et al. (2022, including Falcón-Hidalgo et al. 2020) looked at two nuclear and three chloroplast markers for some 415 terminals in Phyllantheae, and found good support for the monophyly of the clades of Phyllanthus s.l. that they recognized as genera.
Vorontsova et al. (2007) discuss relationships in Poranthereae. Cleistanthus is polyphyletic within Bridelia (Li et al. 2009).
Classification. Hoffmann et al. (2006, q.v. for more details, subtribes included) provide much of the classification followed here. Within Phyllantheae, although Phyllanthus was already a large genus (see Bouman et al. 2018 for an infrageneric classification), it was thought that it perhaps should be extended to include Glochidion (some 300 species), Breynia, Sauropus (70 spp.), etc., genera that are well embedded in Phyllanthus s.l. (e.g. Hoffmann et al. 2006; Lorence & Wagner 2011; Ralimanana et al. 2013; Kawakita & Kato 2017a; Falcón-Hidalgo et al. 2020; c.f. Pruesapan et al. 2008, 2012; van Welzen et al. 2014b). On the other hand, Bouman et al. (2020, esp. 2022) suggest that in addition to retaining genera like Glochidion, Phyllanthus itself should be divided into some 12 genera. Indeed, Bouman et al. (2022) ended up making 645 new combinations in the course of recognizing 10 genera (none new) to take care of Phyllanthus s.l., and they also provided infrageneric classifications for genera in Phyllantheae as a whole. Their classification is being followed here, and all told there are some 18 genera in Phyllantheae - in fact, some genera, like Sauropus, have disappeared, and this is now included in Breynia. For comments on Phyllanthus and its classification, see Muñoz-Rodríguez et al. (2023); they note that Phyllanthus s.l. would have ca 850 species, but the nine genera that Bouman et al. (2022) recognized, albeit monophyletic, were not diagnosed morphologically, while Moonlight et al. (2024) continued to recognize a Phyllanthus s.l. with 1,025 species in their list of hyperdiverse angiosperm genera... See Vorontsova and Hoffmann (2008) for genera in Phyllanthoideae-Poranthereae. The limits of Cleistanthus (Bridelieae) will have to be adjusted, and most species may well need a new name (Li et al. 2009).
Previous Relationships. Phyllanthaceae include most of the old Euphorbiaceae-Phyllanthoideae, minus Drypetes and relatives, for which see Putranjivaceae.
Botanical Trivia. Pollen grains of Phyllanthus that remained on the proboscides of Deltophora moths (Gelechiidae), their pollinators, for some time were chemically dissolved, the moths being the only animals known that can break down sporopollenin (Luo et al. 2011).
PICRODENDRACEAE Small, nom. cons. (the old Euphorbiaceae-Oldfieldioideae) - Back to Malpighiales
Trees or shrubs; picrotoxanes +, otherwise chemistry?; cork?; mucilage cells [epidermis] + (0); subsidiary cells piggy back [on top of guard cells]; hairs unicellular or unbranched-uniseriate; leaves spiral, stipules petiolar, cauline, with axillary colleters, or 0; plant dioecious; staminate flowers: P 4; A 4; pollen ± spherical, 3-8 stephanoporate [= pores ± equatorial], echinate, perforate, smooth/(nano)gemmate/granulate; nectary between or inside A; pistillode +/0; carpelate flowers: P (3-)4-8; ?staminodes; G (2-)3(-7), (style +), stigmas stout, entire, sometimes swollen, dry (wet); ovule with outer integument 5-6 cells across, inner integument 3-6 cells across, nucellar cap +, (nucellar beak 0), hypostase +, funicular obturator +, with hairs; seeds carunculate (caruncle 0), vascular bundle branching in chalaza; exotegmic cells cuboid or fibrous; x = 7 (?6).
25 [list, to tribes]/96 - three groups below. Mostly tropical. Map: from Webster 1994a; van Welzen & Forster 2011; FloraBase i.2012 - approximate.
Age. Estimates for the age of crown-group Picrodendraceae are (92.9-)78(-72) Ma (Xi et al. 2012b; Table S7) and 69.2-59.9 Ma (Grímsson et al. 2019).
Fossil pollen identified as that of Picrodendraceae has been found from the Palaeocene onwards; see Grímsson et al. (2019) for fossils of the family.
1. Podocalyceae G. L. Webster
Vessel element perforation plates various; petiole thickened at both ends; stomata anomocytic; staminate flowers sessile, in clusters; n = ?
2/6: Tetracoccus (5). Amazonia.
[Caletieae + Picrodendreae]: vessel elements with simple perforation plates; ?nodes; (leaves opposite), (2-ranked); P to 13; (styles branched); seeds often carunculate.
2. Caletieae Müller Argoviensis —— Synonymy: Androstachyaceae Airy Shaw, Paivaeusaceae A. Meeuse
Often monoecious; staminate flowers: A to many [>50], (connate basally); pollen ± spherical, zono- to pantoporate; (ovule with endostomal micropyle - Austrobuxus).
14/68: Austrobuxus (20). Largely Australia and New Caledonia, few Malesian.
3. Picrodendreae (Small) G. L. Webster —— Synonymy: Micrantheaceae J. Agardh, Pseudanthaceae Endlicher
Lamina (palmate, or with secondary venation ± palmate), (margins toothed [teeth with deciduous apex]), (stipules large, decurrent); (nectary 0 - Picrodendron); staminate flower: A to 50; pollen (pores (5-)10-50 - pantoporate), (nanoechinate); fruit (1-seeded - Pseudanthus); (exotegmen palisade, subprocumbent, mesotegmen ± thickened, endotegmen 2-layered, with banded thickenings - Oldfieldia); (nucellus/perisperm[?] ruminate), endosperm 0, cotyledons plicate - Picrodendron); n = 12.
9/27: Pseudanthus (9), Aristogeitonia (7). Africa, America, S. India and Sri Lanka, eastern Australia; tropical and warm temperate. Photo: Picrodendron Fruit © A. Gentry.
Evolution: Divergence & Distribution. Grímsson et al. (2019) suggest that the family originated in the Americas, and outline its possible movements since. Thus in Hyenanche (Caletieae), with its distinctive pollen, Grimsson et al. (2024: fossil pollen) suggest an itinerary - North Amarica → Eocene Europe → late Eocene Kenya (lowland, wetland) → South Africa (much drier conditions) - another case where the current distribution and ecology of a genus does not reflect its past.
Ecology & Physiology. The New World Piranhea (Picrodendreae) may dominate flooded forests (Connell & Lowman 1989). Soils of communities dominated by P. mexicanum, with arbuscular mycorrhizae, were very like adjacent soils supporting mixed forests, germination of Piranhea seeds on soils of the two types was indistinguishable, etc. (Martijena 1998: as Celaenodendron).
Seed Dispersal. Myrmecochory occurs in most of the species of the family (Lengyel et al. 2009, 2010).
Vegetative Variation. Leaves are very variable here. Genera like Oldfieldia have opposite, palmately-compound leaves, and in Podocalyx, although the leaves are simple, the petioles are swollen at both ends. The lamina margin is usually entire, but the lamina margins of Austobuxus have minute glands while those of are strongly serrate. A number of taxa have very small leaves ca 1 cm long or less, and these are sessile and trifoliolate in Micranthemum. Stipules are often minute, but in Androstachys they are large, intrapetiolar and enclose the bud.
Chemistry, Morphology, etc.. Podocalyx probably has trilacunar nodes (e.g. leaf scars on Aracá Inventory TF-2-20).
Picrodendron may have a perianth of two whorls (Hakki 1985), or perhaps it is modified quincuncial, or perhaps there is no perianth at all, at least in staminate flowers... (Hayden et al. 1984). Grímsson et al. (2019) discussed pollen morphology in some detail, noting i.a. that finely striate markings might be evident on the spines and their immediate surrounds of fossil pollen placed in this family, and sometimes in pollen of extant taxa. For more on pollen morphology with a focus on Pseudanthus, see Till et al. (2025: see esp. Table 3, 2026: Table 3 again, more elaborate).
Additional information is taken from Radcliffe-Smith and Esser (2001: genera, etc.), Hayden et al. (1984: Picrodendron), Wurdack et al. (2004), van Welzen & Forster (2010), and Webster (2013), all general, Westra and Koek-Noorman (2004) and Hayden (1977, 1994), all wood anatomy, Merino Sutter et al. (2006: morphology of carpelate flowers), Levin and Simpson (1994: pollen), and Huber (1991) and Tokuoka and Tobe (1999), both seed anatomy.
The family is poorly known.
Phylogeny. See Wurdack et al. (2004) and Xi et al. (2012b) for outlines of the phylogeny; the relationships are well supported. Grímsson et al. (2019) suggested the relationships [[Podocalyx + Caletieae] [Tetracoccus + Picradendreae]]; Hyaenanche was nestled within Picrodendreae.
Stuppy (1996) noted that both Picrodendron and Oldfieldia were rather different from other taxa included in his Oldfieldioideae, and he compared the latter with Meliaceae because of similarity in seed characters. However, both genera are firmly in Picrodendraceae-Picrodendreae. Two other genera have been associated with Picrodendraceae in the past. Paradrypetes is probably to be included in Rhizophoraceae, although it is like Podocalyx (see above) in particular in wood anatomy and pollen morphology (Levin 1992). Croizatia (see Levin 1992) is also odd, with 5 petals, an extrastaminal nectary and style with distinct branches, and it lacks the distinctive pollen of the family; it is here placed in Phyllanthaceae-Phyllanthoideae-Bridelieae.
Classification. For genera, see Euphorbiaceae-Oldfieldioideae (Webster 1994b). Govaerts et al. (2000) provides a checklist and bibliography (as Euphorbiaceae).
Previous Relationships. Picrodendraceae are the old Euphorbiaceae-Oldfieldioideae (see Webster 1994b).
[Ixonanthaceae + Linaceae] / Linoids: cristarque cells +; lamina vernation involute; flowers perfect; C contorted [direction not fixed]; ovary loculi divided [?]; ovule endothelium +, parietal tissue 2-5 cells across, hypostase +; endosperm scanty, cotyledons large.
Age. Linaceae may have diverged from other Malpighiales in the Cretaceous-Albian 111-100 Ma (Davis et al. 2005a); (103.4-)90(-73.6) Ma is the spread in Xi et al. (2012b; Table S7) and (87.9-)78.6(-73.6) Ma in Schneider et al. (2016a).
Phylogeny. This family pair has strong support (Xi et al. 2012b).
Chemistry, Morphology, etc.. For foliar anatomy, see van Welzen and Baas (1984).
IXONANTHACEAE Miquel, nom. cons. - Back to Malpighiales
Trees; ellagic acid, myricetin 0; vessel elements with simple perforation plates; mucilage cells 0; cuticle waxes as variously arranged platelets; petiole bundle annular (with medullary bundle) or arcuate; (foliar sclereids +); branching from previous flush; leaves spiral, lamina (margins entire), stipules cauline; inflorescences determinate, corymbose or, axillary; (pedicels articulated); K usu. basally connate, (C imbricate); A [and style] folded in bud, 5, opposite K [no staminodes]/10, obdiplostemonous/-20 [in triplets opposite K]; pollen with supratectal spines; nectary between bases of and adnate to filaments, unvascularized, or ± adaxial, vascular tissue from A traces; G [(2) 5] (?fale septae 0), style unbranched, slender, stigma capitate or discoid; ovule micropyle bistomal, (outer integument very long, expanded), suprachalazal zone massive, (obturator +, hairy); fruit valves opening adaxially as well, K and C persistent; seeds basally winged, or aril arising between the hilum and micropyle, (= wing - Ochthocosmus); endotegmen with sinuous anticlinal cell walls; (endosperm 0); n = 14, chromosomes 0.4-1.1 µm long.
3 [list]/13: Ochthocosmus (8). Pantropical. Map: from Aubréville (1974), Kool (1988) and Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003).
Age. Estimates of the crown group age of Ixonanthaceae are (75.4-)51.9(-26.6) Ma (Xi et al. 2012b; Table S7) and (74.3-)48.7-)17.5 Ma (A. C. Schneider et al. 2016).
Evolution: Divergence & Distribution. Ixonanthaceae may have diverged in the Cretaceous-Albian 111-100 Ma (Davis et al. 2005a: c.f. topology).
Chemistry, Morphology, etc.. The stamens opposite the petals in Ixonanthes are paired, but arise from a single trace (L. L. Narayana & Rao 1966). Narayana (1970) depicted a tegmen ca 4 cells thick the innermost layer of which is an endothelium.
See also Forman (1965, but not Allantospermum), Kool (1988) and Kubitzki (2013b), all general, Nooteboom (1967: esp. chemistry), Weberling et al. (1980: stipules), Link (1992c: nectary), and Rao and Narayana (1965) and L. L. Narayana and Rao (1978a), both embryology, etc., for more information. Kool (1980) revised Ixonanthes.
Much remains unknown in this family.
Phylogeny. Relationships in Xi et al. (2012b) are [Ixonanthes [Cyrillopsis + Ochthocosmos]] - support strong (see also Byng et al. 2016; Sun et al. 2016). A. C. Schneider et al. (2016) found that Ixonanthes was paraphyletic, I. icosandra being sister to the rest of the family, although other relationships were as just mentioned.
Previous Relationships. The circumscription and relationships of Ixonanthaceae have been particularly unclear (see below). Allantospermum (now Irvingiaceae) was included here prior to xii.2015.
LINACEAE Perleb, nom. cons. - Back to Malpighiales
Cork?; vessel elements with simple or scalariform perforation plates; true tracheids +; petiole bundle(s) arcuate; epidermal wax crystals as parallel platelets; branching from previous innovation; lamina tooth ?type, petiole short; pedicels articulated; (flowers distylous); K quincuncial, C (trace single), postgenitally connate above base, caducous; nectary outside A; A basally connate, basally adnate to C, anthers basifixed; tapetum binucleate; pollen grains tricellular, starchy; G [2-5], opposite C, or median member adaxial, style more or less divided, stigmas ± capitate; ovule (1/carpel), micropyle endostomal (bistomal), endothelium +, obturator +, with papillae; (megaspore mother cells several); fruit often septicidal, K persistent; tegmen strongly multiplicative; endosperm with chalazal haustorium, variable, (embryo slightly curved), cotyledons large; x = 6 (?9, ?7), nuclear genome [1 C] (0.049-)1.217(-29.926) pg.
Ca 7 [list: to subfamilies]/300 (235)- two subfamilies below. World-wide.
Age. Crown Linaceae have been dated to (47-)36, 35(-23) Ma (Bell et al. 2010); McDill and Simpson (2011, q.v. for more details, as in Bell et al. 2010: Irvingiaceae sister to Linaceae) suggest that crown group divergence was in the early Caenozoic (range 82-43 My); (54.8-)39.5(-28.9) Ma is the age in Xi et al. (2012b; Table S7) and (51.6-)43.9(-37.7) Ma in A. C. Schneider et al. (2016a).
1. Linoideae Arnott
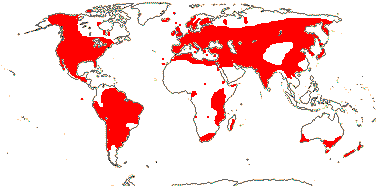
Annual to perennial herbs (shrubs); ellagic acid 0; vessel elements with simple perforation plates; rays uniseriate; (nodes 1:1 - Linum); cristarque cells uncommon; epidermis mucilaginous; leaves opposite or spiral, (lamina margins entire), (stipules 0); K ± equal, C clawed, protective in late bud; (nectary at base of C or A); A 5, opposite K, alternating with staminodes; pollen 3-many colpate, many colporate, or pantoporate, surface ± intectate, verrucate or echinate; G ([2-4]), loculi usu. divided, stigma unifacial, wet or dry; ovules with outer integument 2(-3) cells across, inner integument 3-12 cells across, parietal tissue 0-1 cell across, (nucellar caps +), hypostase?, (lateral tissue scanty), (obturator papillae 0); (2-seeded mericarps also splitting along false septae, units opening adaxially); exotesta with outer walls massively thickened, mucilaginous, tegmen multiplicative, cross cells beneath exotegmen, endotegmen tanniniferous; endosperm with xyloglucans, (helobial), embryo chlorophyllous [Linum]; n = 6, (8), 9, (11-18, etc.); nuclear genome [1C] ca 373 Mb; plastid transmission biparental [Linum].
4/240: Linum (200). Worldwide, but esp. N. temperate and subtropical. Map: from Hultén & Fries (1986), Trop. Afr. Fl. Pl. Ecol. Distr. 1. (2003), Diderichsen and; Richards (2003), Flora of China vol. 11 (2008) and McDill et al. (2009). [Photo - Flower.]
Age. McDill and Simspon (2011) suggested that divergence in the crown group of Linoideae is Eocene in age, while (41-)39.2(-37) Ma is a similar age suggested by A. C. Schneider et al. (2016a).
2. Hugonioideae Hooren & Nooteboom —— Synonymy: Hugoniaceae Arnott
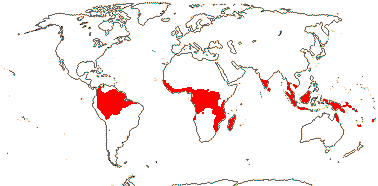
Trees or shrubs, often lianas with branch grapnels; ellagic acid?; vessel elements with scalariform perforation plates; sclereids +; stomatal accessory cells usu. lignified (not Indorouchera), lobed beneath the guard cells; leaves spiral or two-ranked, (stipules pectinate); (flowers tristylous); K often unequal, C at most slightly clawed, often yellow, (0); A 10, of two lengths, (obdiplostemonous); (pollen inaperturate); G (2-)3-6, (opposite K), styles impressed, (stigma bilobed); ovule with micropyle endostomal [Roucheria], outer integument 2-3 cells thick, inner integument 3-12 cells thick; fruit a drupe or with mericarps; seed with an at most slight arillode, testa multiplicative, mesotesta with sclerotic cells, endotesta lignified, exotegmen barely lignified or tegmen obliterated; (endosperm copious); n = 6, 12, 13.
3/61: Hugonia (40). Pantropical. Map: from van Hooren and Nooteboom (1984a), Jardim (1999), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003) and McDill et al. (2009). [Photo - Flower.]
Age. McDill and Simspon (2011) suggested that crown-group Hugonioideae are Oligocene-Miocene in age, although A. C. Schneider et al. (2016a) suggested a late Miocene age of a mere (13.5-)7.6(-2.9) Ma.
Evolution: Ecology & Physiology. Hesperolinon (= Linum s.l.) is notably diverse on the serpentine soils of the western United States (A. C. Schneider et al. 2016a).
Pollination Biology & Seed Dispersal. Heterostylous flowers are scattered in Linoideae, and tristyly is reported from at least some Hugonioideae (Barrett & Shore 2008; Meeuse et al. 2011; Cohen 2019), but whether heterostyly is an apomorphy for Linaceae is unclear; breeding systems have certainly been labile (Thompson et al. 1996; McDill et al. 2008, 2009). In Linum suffruticosum, at least, heterostyly involves positioning of anthers and stigmas so that pollen is placed on the insect in quite different places (dorsally, ventrally) by the anthers of the flowers of the different morphs (Armbruster et al. 2006).
A number of Linoideae have myxospermous seeds (references in Western 2012), and Kreitschitz and Gorb (2017) provide a microstructural analysis of the pectic seed mucilage of Linum.
Genes & Genomes. Two genome duplications in the Linum clade have been reported (Cai et al. 2017/18: no other Linaceae examined); Z. Wang et al. (2012) dated a duplication that was shared with L. bienne to 9-5 Ma, the other one detected by Cai et al. is much older. For variation in genome size in Linum see Wang et al. (2012).
There has been extensive rearrangement of the plastome in Linum usitatissimum and numerous RNA editing sites not found elsewhere have been reported (de Santana Lopes et al. 2017).
Economic Importance. Seeds of flax (Linum usitatissimum) have been used for oil, etc., for about 10,000 years (Vaisey-Genser & Morris 2003; Vollmann & Rajcan 2009), while linen is made from its stem fibres.
Chemistry, Morphology, etc.. Ellagic acid is not known from Linoideae, but members of this subfamily are largely herbaceous. Flat vernation is reported from Linum narbonense by Cullen (1978), other taxa may be conduplicate. Bracts and bracteoles have a single vascular trace ().
Tirpitzia bilocularis has a corolla tube over 2 cm long. The "staminodes" may lack any vascularization (Al-Nowaihi & Khalifa 1973). Tobe and Raven (2011) suggest that the inner integument is multiplicative. In Hugonioideae, only half of the ovules may develop and produce seeds. Guignard (1893) drew the ovules of Linum as having a single parietal layer of cells; at the base of the embryo sac there was a persistent narrow column of cells, while Boesewinkel (1980a) also suggested there might on occasion be a single layer of parietal cells and a nucellar cap two cells across. Note that there may be septicidal dehiscence, presumably liberating pyrenes,in Linoideae (Spichiger et al. 2002).
For general information on the family and its possible segregates see Hallier (1923), Robertson (1971: Linoideae), van Hooren and Nooteboom (1984, 1988a), Jardim (1999: New World Hugonioideae) and Dressler et al. (2013); see also Hegnauer (1966, 1989: chemistry), Schmidt et al. (2010: lignans, in most Linum alone), van Welzen and Baas (1984) and Kumar (1977), both anatomy, Cremers (1974: growth in Hugonia), L. L. Narayana (1964, 1970), D. Rao and Narayana (1965), Rao (1968) and Narayana and Rao (1966, 1969, 1973: Philbornea magnifica, 1978b), all embryology, floral anatomy, etc., and Schewe et al. (2011: floral development in Linum).
Phylogeny. McDill et al. (2009: focus on Linoideae) outlined phylogenetic relationships in the family; Linoideae may be monophyletic, but support is from posterior probabilities only; the status of Hugonioideae is unclear. However, in the more extended sudy of McDill and Simpson (2011) Linoideae are well supported, although Linum is paraphyletic and its current sectional limits need adjusting, Hugonioideae are monophyletic, but still lacking strong support. A. C. Schneider et al. (2016) recovered both subfamilies with good support, but c.f. Sun et al. (2016). Within Hugonioideae, Hugonia was paraphyletic and included Philbornea and Indorouchera; the Australian H. jenkinsii was sister to the combined clade (Schneider et al. 2016a).
Classification. The topology suggested by McDill and Simpson (2011) and Schneider et al. (2016a) necessitates nomenclatural adjustments. It is best to expand Linum; certainly, Cliococca must go, and if there is a desire to maintain Hesperolinon (it does have a sectional name in Linum), then another four or five genera will be needed. Generic limits around Hugonia are also difficult.
Previous relationships. For an early detailed discussion on relationships of Linaceae, then thought to be a "central" family, see Hallier (1923). Linaceae have been linked with Erythroxylaceae (the two have even been included in the same family) and Humiriaceae (Prance 2021 notes the similarity in wood anatomy to that of Linaceae), and thence to Geraniales (e.g. L. L. Narayana & Rao 1978b), or the three families together are placed in Linales (Cronquist 1981); see also Boesewinkel and Geenen (1980)
[[Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]], [Irvingiaceae + Pandaceae], [Ochnaceae [[Bonnetiaceae + Clusiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]] / Clade 2 of Xi et al. (2012b) / [Rhizophoroids, Pandoids [Ochnoids + Clusioids]]: cristarque cells +; anthers basifixed.
Age. The crown age of this clade is (110.9-)106.6, 105.2(-99.9) Ma (Xi et al. 2012b: Table S7) or around 101.1/100.5 Ma (Tank et al. 2015: table S1, S2).
Chemistry, Morphology, etc.. The ovule here is weakly crassinucellate, or with a tendency to be incompletely tenuinucellate (Endress ).
Phylogeny. For discussion of relationships in this clade, see above.
[Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]] / Rhizophoroids: leaves opposite, stipules enclosing the terminal bud, interpetiolar; pedicels articulated; K with <3 traces; nectary outside of A; A 2 x C [antepetalous A longer than antesepalous], connate basally, (minute corona +); G postgenitally united, placentation apical, septae often thin, stigmas capitate/lobed, papillate; ovules 2/carpel, collateral, epitropous, micropyle zig-zag, outer integument thinner than the inner [inner integument 5< cells across], nucellus laterally thin, disintegrates, endothelium +, placental obturator +; K persistent in fruit; seeds arillate, exotestal; endosperm +.
Age. The crown age of this clade is around (68.6-)66.7(-65.4) Ma (Xi et al. 2012b: Table S7).
Chemistry, Morphology, etc.. Matthews and Endress (2011) provided many details of the floral morphology of these three families. Tobe and Raven (2011) suggested that all have a multiplicative inner integument, rather, at least sometimes it is very thick by the time of fertilization.
Phylogeny. For relationships in this clade, all well supported, see Xi et al. (2012b).
CTENOLOPHONACEAE Exell & Mendonça - Ctenolophon Oliver - Back to Malpighiales
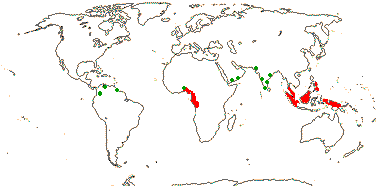
Trees; ellagic acid?; vessel elements with scalariform perforation plates; calcium oxalate as single crystals; cuticle waxes 0; stomata anomo- or anisocytic; petiole bundle arcuate; hairs tufted/stellate; buds perulate; lamina margins entire; inflorescence terminal, ?thyrsoid; K quincuncial, basally connate, (with 1 trace), C protective in bud, contorted, [direction ?not fixed], caducous; nectary annular, 10 lobes alternating with A; A adnate to base of nectary; pollen 3-9 equatorially colporate [stephanocolporate]; G [2], septae thin, style +, branches short; ovules apical [?level], with zig-zag micropyle, integuments lobed, outer integument ca 5 cells across, inner integument ca 11 cells across; fruit a [?kind] capsule, K swollen; seed single, persisting on columella, columella very thin; aril ± hairy [when dry!], exotestal cells large, subpalisade, the outer wall alone thickened, exotegmic cells laterally flattened, tracheidal; endosperm copious, cotyledons very large, folded; n = ?.
1 [list]/3. W. Africa, Malesia. Map: from van Hooren and Nooteboom (1988b), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003) and fossils [green] from Krutzsch (1989).
Age. The distinctive pollen of Ctenolophon is known fossil from South America and India, appearing more or less simultaneously on both sides of the Atlantic 84-74 Ma (Muller 1981; Krutzsch 1989; Morley 2003).
Evolution: Divergence & Distribution. Morley (2003) suggested movement via Africa to India and tropical Southeast Asia, also to South America (see also Arecaceae, Dipterocarpaceae, Ebenaceae and Euphorbiaceae-Crotonoideae).
The diversification rate in this clade may have decreased (Xi et al. 2012b).
Chemistry, Morphology, etc.. Like Humiriaceae, there are "marginal" stomata on the nectary and the anthers have a broad connective (Link 1992b); their wood anatomy is also similar. Takhtajan (1999) perhaps implies that there may be an endothelium, but embryology, etc., are largely unknown.
Some general information is taken from van Hooren and Nooteboom (1984, 1988b) and Kubitzki (2013b); for embryology, see D. Rao and Narayana (1968) and for seed anatomy, see Huber (1991).
Previous Relationships. "Ctenolophon was almost universally recognized as belonging to the Linaceous alliance" (van Hooren & Noteboom 1988: p. 629).
[Erythroxylaceae + Rhizophoraceae]: tropane [hygroline] alkaloids, pyrrolidine alkaloids, non-hydrolysable tannins +; sieve tube plastids with protein crystalloids; mucilage cells common; stomata paracytic; lamina vernation involute, colleters +; inflorescence cymose; K valvate, with large idioblasts in the mesophyll, postgenitally united, C ± clawed, basally valvate, conduplicate, each enclosing 1-several A; median G adaxial, style somewhat impressed; (micropyle endostomal); fruit a septicidal capsule; exotestal cells enlarged, thick-walled, ± tanniniferous; endosperm starchy, embryo chlorophyllous.
Age. Estimates for the age of this node are (58-)54, 49(-45) Ma (Wikström et al. 2001), (63.1-)54.6(-38.4) Ma (Xi et al. 2012b: Table S7), (79-)63, 60(-44) Ma old (Bell et al. 2010), or around 74.7 Ma (Tank et al. 2015: table S2) - Davis et al. (2005a) would put the age at (119-)114(-110/(106-)102 Ma, but in the context of a different topology, while L. Cai et al. (2025b) suggested an age of ca 98.4 Ma.
Evolution: Divergence & Distribution. Although [Erythroxylaceae + Rhizophoraceae] are a somewhat unexpected family pair when contrasting Erythroxylum with mangrove Rhizophoraceae (= Rhizophoreae), the latter are very derived morphologically, so when comparing Aneulophus (Erythroxylaceae) with non-mangrove Rhizophoraceae, the differences are less stark. Furthermore, as noted above, the two families are united by several synapomorphies; for tropane alkaloids, see Y.-J. Wang et al. (2023), but note that how these alkaloids are handled in the characterizations here may be incorrect.
Chemistry, Morphology, etc.. For floral development, see Matthews and Endress (2007).
ERYTHROXYLACEAE Kunth, nom. cons. —— Synonymy: Nectaropetalaceae Exell & Mendonça - Back to Malpighiales
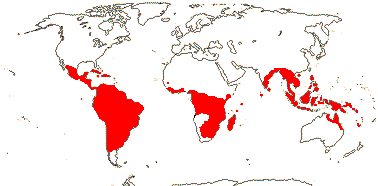
Smallish trees and shrubs (deciduous); mycorrhizae 0; tropane alkaloids ecgonone/cocaine type , ellagic acid 0; vessel elements with simple perforation plates; wood commonly with SiO2 grains; nodes with lateral bundles originating well before the central, forming cortical bundles; sclereids +; petiole bundle arcuate to annular with medullary and adaxial bundles; stomata also parallelocytic; branching from previous flush; buds perulate; leaves two-ranked (spiral; opposite), lamina margins entire, stipules intrapetiolar and hooded or interpetiolar; inflorescence often fasciculate; (pedicel not articulated - Aneulophus?), heterostyly common; (hypanthium + - Nectaropetalum); K connate basally, C protective in bud, with fringed bilobed ligule (0); nectary glands just below ligule; A obdiplostemonous, latrorse, (connective not thickened); pollen grains tricellular; G [(2-)3(-4)], adaxial only fertile [Erythroxylum], (short), (stylar canal +), style branches ± well developed, stigma ± capitate; ovule also 1/carpel, outer integument 2-5 cells across, inner integument (ca 3?-)5-9 cells across, endothelium +, parietal tissue 2-4 cells across, suprachalazal zone extensive, hypostase 0, raphe with 2 vascular bundles; fruit a drupe, 1-seeded, longitudinally ± ridged/capsule [Aneulophus], A also persistent; (aril 0); testa weakly multiplicative, exo- and endotesta tanniniferous, tegmen strongly multiplicative or not, exotegmen with reticulate thickenings [?all], innermost cuticle well developed; (endosperm 0); n = 12, x = ?7 (?6).
4 [list]/240: Erythroxylum (230). Pantropical, esp. American (map: from van Steenis and van Balgooy 1966; Heywood 1978; Trop. Afr. Fl. Pl. Ecol. Distr. 1. 2003). [Photo - Flower, Fruit.]
Age. The crown age of Erythroxylaceae is (42.5-)19.3(-12.3) Ma (Xi et al. 2012b: Table S7, An. + Ery.).
Evolution: Divergence & Distribution. The rate of diversification may have increased in Erythroxylaceae (Xi et al. 2012b).
Y.-J. Wang et al. (2023) discuss details of the synthesis of the distinctive tropane alkaloids found in both Erythroxylaceae and Solanaceae. Although they share a common intermediate, ecgonone, which has the characteristic tropane ring, the end products, for example hyoscyamine (from the tropinone pathway) and cocaine (from the ecgonone pathway), are synthesized via different pathways. Comparing Erythroxylum novogranatense with Anisodus acutangulus there is also substantial variation in how ecgonone is synthesised, furthermore, even when a step in the synthesis involves the same substrate and product in both taxa, different enzymes may be involved in the two, an example being the conversion of malonyl-CoA to 3-oxo-glutaric acid, three steps before the production of ecgonone itself (Wang et al. 2023: Fig. 1). Somewhat complicating the story, Wang et al. (2023) noted that the tropinone pathway occurs in Rhizophoraceae, but I do not know exactly what goes on there.
Plant-Animal Interactions. Cocaine is sequestered by the larvae of Eloria noyesi (most is excreted), a lymanitrid moth that feeds on Erythroxylum and are something of a pest in plantations of E. coca; cocaine is also found in the adult moth (Blum et al. 1981).
Genes & Genomes. Erythroxylum (Rhizophora sister) has a genome duplication (Cai et al. 2017/18).
Chemistry, Morphology, etc.. The nodes were described as being unilacunar by Sinnott (1914), however, there are lateral traces although their gaps may be inconspicuous and the traces themselves may depart from the vascular cylinder well before the central trace (Rury 1982). Erythroxylum sometimes has milky exudate. Are the lamina teeth theoid? The leaves of Erythroxylum coca were described as being revolute by Cullen (1978); they are involute (e.g. Peyritsch 1878; Weberling et al. 1980; Rury 1982; Keller 1996).
Matthews and Endress (2011) described the complexity of the postgenital fusion of the petals.
For general information, see van Tieghem (1903d: inc. some anatomy) and Bittrich (2013), for chemistry, see Hegnauer (1966, 1989) and Aniszewski (2007), for foliar anatomy, see van Welzen and Baas (1984), and for ovule and seed, see D. Rao (1968) and Boesewinkel (1980).
Phylogeny. Aneulophus is sister to the rest of the family (M. Sun et al. 2016).
Previous Relationships. In the past, Erythroxylaceae have been associated with Linaceae and Humiriaceae, and thence linked with Geraniales (L. L. Narayana & Rao 1978b), or the three families together are placed in Linales (Cronquist 1981); some have even toyed with the idea of including Erythroxylaceae and Linaceae in the one family (Boesewinkel & Geenen 1980 and references).
RHIZOPHORACEAE Persoon, nom. cons. - Back to Malpighiales
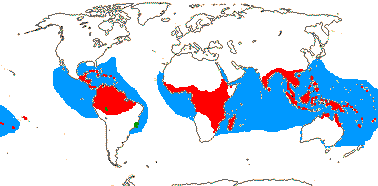
Trees; tropane alkaloids tropinone type, ellagic acid +; root hairs 0; vessel elements with simple and/or scalariform perforation plates; true tracheids +; pits vestured; cristarque cells 0; subepidermal laticifers in flower [in K, G]; branching from current flush; inflorescence axis often evident; K (3-)4-5(-16), C often hairy, variously lobed, fringed, with filiform appendages or aristate, reflexed; anthers ± dorsifixed, (fasciculate), (free); nectary inside A [on ovary or hypanthium]; G opposite K when 5, when 2, collateral, septae often thin/disintegrating, style +, stigma also ± punctate, ?type; (micropyle also zig-zag), outer integument 3-6 cells across, inner integument 4-8(-20?) cells across, (endothelium 0), parietal tissue 1-3 cells across; megaspore mother cells several; (endotesta crystalliferous); endosperm with micropylar and chalazal haustoria [?distribution], embryo (short), chlorophyllous; x = 7 (?6), chromosomes ca 1 µm long; germination epigeal, cotyledonary node unilacunar.
16 [list, to tribes]/149. Four groups below. Pantropical. Map: from Ding Hou (1958), van Steenis (1963), Fl. Austral. vol. 8 (1984), Tomlinson (1986), Juncosa and Tomlinson (1988a), Levin (1992), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003). [Photo - Flower, Flower, Fruit.]
Age. The crown age of this clade is (54.5-)41.7(-30.4) Ma (Xi et al. 2012b: Table S7, c.f. topology).
[Macariseae + Paradrypetes]: ?
1. Macarisieae Baillon —— Synonymy: Cassipoureaceae J. Agardh, Legnotidaceae, nom. illeg., Macarisiaceae J. Agardh
Nodes 1:1 + split laterals [?all]; calcium oxalate crystals solitary; (leaves bijugate - Cassipourea), ("alternate"), stipules valvate; (hypanthium +); K open; A (many), anthers latrorse; G [2-6], stigma not lobed; seeds winged at micropylar end/arillate; n = 18, 21, 32; (seedlings with root hairs - Cassipourea).
7/94: Cassipourea (62), Dactylopetalum (15). Tropical America and Africa, also peninsula India and Sri Lanka.
2. Paradrypetes Kuhlmann
Raphides +; plant glabrous; lamina with long, zig-zag intersecondary veins; plant dioecious; inflorescence epiphyllous, on petiole; flowers small; P uniseriate, 3-4, imbricate; staminate flowers: anthers extrose, filaments ± 0; pollen 4-colporate, surface spiny; nectary 0; pistillode 0; carpelate flowers: staminodes 0[?]; G [3], style 0, stigmas broad; ovule with placental obturator; fruit a drupe, 1-seeded; seed coat vascularized; endosperm starchy, abundant, cotyledons plicate, broad; n = ?
1/2. Upper Amazon and Atlantic Forest, Brasil. Map above: green.
[Gynotrocheae + Rhizophoreae]: stilt roots +, root hairs 0; laticifers radially expanded; leaves bijugate (not - Pellacalyx), stipules imbricate; hypanthium +; ovary ± inferior; obturator 0; fruit indehiscent; aril 0, testa vascularized.
Age. This node has been dated to (15-)13, 9(-7) Ma (Wikström et al. 2001) or (15-)9, 8(-2) Ma (Bell et al. 2010), very unlikely estimates (see below), and ca 54.6 Ma (S. Xu et al. 2017b).
3. Gynotrocheae Engler
(Stilt roots 0 - Pellacalyx [P.]); (laticifers 0, idioblastic secretory cells - Gynotroches [G., P.), (lamina margins entire), (stipules not sheathing terminal bud); (plant dioecious - G.); (petals entire); A (5, opposite K - Carallia), (antepetalous A initiated first), (many - Kandelia); G [3-28], septae ± developed or not, loculus partly schizogenous when G many [P., Crossostylis], (style branched - G.); ovules (to 8/carpel), outer integument 2-3 cells across, inner integument 2-6 cells across, (parietal tissue 0), endothelium +; (megaspore mother cell 1); fruit a berry; (seed arillate - Crossostylis); exotesta mucilaginous, tanniniferous, other testal cells crystalliferous, tegmen 0, or fibrous to palisade, meso- and endotegmen persist; cotyledons short, or large, involute [Carallia, P.]; n = 14.
4/30: Crossostylis (10). Indo-Malesia, Madagascar.
4. Rhizophoreae Bartling —— Synonymy: Mangiaceae Rafinesque
Plants salt tolerant; root cortical phi [φ] cell wall thickenings radially oriented [?all]; nodes 5:5, 7:7, + split-laterals; cortical, etc., fibres +; stomata cyclocytic; abaxial hypodermis +; sclerenchymatous sheath of midrib at most weakly developed; colleters +. numerous [?level]; lamina vernation supervolute, margins entire; flowers 4-16-merous; (C postgenitally united above base), (with hydathodes); A (many - Kandelia), (anthers locellate - Rhizophora [R.]); G [2-3]; outer integument ca 25 cells across, inner integument 5-6 cells across, parietal tissue ca 2 cells across, endothelium ?+; fruit indehiscent, 1-seeded; seeds large, coat undifferentiated, integument vascularized, tegmen not persisting; endosperm overflows from seed; (cotyledons connate - R.; convolute - R., Bruguiera); n = 18; nuclear genome [1C] 291-186 Mb; seeds viviparous/epigeal, cotyledonary node tri- (multi- lacunar, radicle 0 [R.].
4/17: Rhizophora (?9). Pantropical, centred on the eastern Indian Ocean, introduced into the central Pacific and Hawai'i. Map above: blue, also Spalding et al. (2010).
Age. Ricklefs et al. (2006) dated crown Rhizophoreae to ca 50 Ma while ca 40.7 Ma is the age in S. Xu et al. (2017b) and ca 49.8 Ma that in Pootakham et al. (2022).
Evolution: Divergence & Distribution. The fossil record of Rhizophoreae was evaluated by Graham (2006); see also Lo et al. (2014) for fossils of Rhizophora. For more on mangroves, their physiology and evolution, see the next section,
Juncosa (1988: esp. Fig. 1) discussed the evolution of 34 floral and vegetative characters in Rhizophoreae; both Macarisieae and Cynotrocheae as shown there are paraphyletic. Crossostylis, with dehiscent fruits and arillate seeds, is embedded in Gynotrocheae, which otherwise have fleshy, indehiscent fruits and seeds without arils. Dehiscent fruits may have evolved in parallel in Crossostylis - Schwarzbach (2013: p. 291) describes dehiscence there as "tardy with distal slits or an operculum", and this seems rather different from the septicidal fruits of Macarisieae, but both may have arillate seeds. At least some Macarisieae have stamens of two lengths and well-developed anther connectives (D. Kenfack, pers. comm.), probably plesiomorphic features.
The evolution of Rhizophoreae has been linked with the Palaeocene-Eocene thermal maximum (PETM) of about 55 Ma (50 Ma in S. Xu et al. 2017b: crown-group Rhizophoreae estimated to be ca 40.7 My). Mangrove taxa are derived within Rhizophoraceae (e.g. Schwarzbach & Ricklefs 2000) and are most diverse in the Southeast Asia-Malesian area (see below). Lo et al. (2014, see also Friess et al. 2019) discuss the biogeography of Rhizophora and suggest that it initially evolved along the shores of the Tethys Sea in the Late Cretaceous. They note a number of long distance dispersal events, including, for example, the movement of R. mucronata from Australia to East Africa, Central America to Fiji (see R. samoensis!), across the Atlantic, etc. - oceanic islands may have served as way stations for such events (Lo et al. 2014). Indeed, Rhizophoreae, and Rhizophora in particular, may be most effectively dispersed by ocean currents (e.g. Lo et al. 2014; Van der Stocken et al. 2018); see also below.
Heads (2019a) suggested that mangrove associates (see also below) had played a key role in evolution, both of angiosperms in general and of Meliaceae in particular, as well as in groups like Malesian Ericaceae (Heads 2003). Such a central role seems rather unlikely.
Ecology & Physiology. The term "mangrove" refers both to members of Rhizophoraceae-Rhizophoreae in particular and to mangrove vegetation and the plants that grow in it in general. The focus in the discussion below is on mangrove vegetation - for general accounts, see Tomlinson (1986, 2017), Spalding et al. (2010), Faridan-Hanum et al. (2014: Asian mangroves), Hogarth (2015), Friess et al. (2019) and Suvarna Raju (2021: pollination). Rhizophoreae are indeed a prominent component of mangrove vegetation, but the latter also includes a few palms and members of several other families. For the evolution of the mangrove ecosystem as a whole, which includees diversification of clades of molluscs, etc., that are found there (Reid et al. 2008), see e.g. Ellison et al. (1999), especially Plaziat et al. (2001 and references), Ricklefs et al. (2006) and He et al. (2022).
The true mangroves, a mere 34 species in nine genera and five families, dominate mangrove vegetation worldwide, and of these plants, half are Rhizophoraceae-Rhizophoreae (4 genera, 17 species), otherwise the taxa are largely unrelated. There are another around 20 species in 11 genera and ten families (only one also including true mangroves) that are quite common, and these are the mangrove associates (Tomlinson 1986); estimates in Spalding et al. (2010) are 38 "core" species, 73 species of "true" mangroves, while Saenger et al. (2019) estimated that there were 81 species of mangroves of any sort (trees and shrubs) in 30 genera and 17 families. Other families that include true mangroves are Primulaceae-Myrsinoideae (Aegiceras), Lythraceae (Sonneratia, ca 7 spp.), Acanthaceae (Acanthus ilicifolius and the seven species of Avicennia - the two genera are not immediately related), Tetrameristaceae (Pelliciera) and Combretaceae (Lumnitzera, Laguncularia, all told 3/8 species of Laguncularieae). The palm Nypa fruticans in particular forms extensive monospecific stands growing along rivers to the upper limits of tidal influence in South East Asia—Malesia.
There are two geographical groups of mangroves. The eastern group, from east Africa to the western Pacific (perhaps most diverse in the Makassar Straits area - Saenger et al. 2019), is much more speciose and includes ca 40 species of true mangroves and mangrove associates, ca 14 of which are Rhizophoreae, while the western group, from west Africa to the Americas, is made up of only eight species, including three Rhizophoreae, Avicennia germinans, etc.. Depending on how species limits are drawn, no species of true mangroves are common to the two areas (Tomlinson 1986, 2016), and no true mangrove has a non-mangrove sister taxon restricted to the New World (Ricklefs et al. 2006). This division of mangroves into the Indo-West Pacific and the Caribbean-West Atlantic areas is variously estimated as being in place by ca 20 Ma (Plaziat et al. 2001) or (50.7-)47.6(-44.5) Ma (Lo et al. 2014), However, note that pollen of Nypa is reported from the Caribbean area, but only before the Eocene-Oligocene transition ca 24 Ma, Pelliciera becoming much less common then and Rhizophora much more common (Rull 2023b: Fig. 3). H.-F. Yan et al. (2024) suggested that the age of the Avicennia clade was around 13.5 Ma. He et al. (2018) discussed speciation in eastern mangroves, emphasizing the role of the Straits of Malacca in the isolation and speciation of mangroves there, but there was also migration and gene flow. There can be considerable genetic differentiation within Atlantic populations of mangrove species (Takayama et al. 2008a, b). Overall, Ricklefs et al. (2006) thought that diversity in the mangrove ecosystem seemed to have increased regularly over time, with little extinction. However, Z. He et al. (2022) looked at the genomes of the majority of obligate mangroves and some of their relatives and suggested that changes in sea level may well have caused extinctions, and another result of these changes was that population contractions were common and within-species heterozygosity was low; speciation rates also seemed to be on the low side.
Mangrove species can have quite wide distributions, and many of them have floating propagules; the movement of such propagules around the globe has been modelled by Van der Stocken et al. (2019) and not surprisingly depends on how long viable propagules can float. Simulations suggest that there is currently no connection between mangrove populations on the two sides of both the African and American continents, although of course the Isthmus of Panama, fairly recent, is reponsible for the latter barrier. There are no native mangroves on the Hawai'an islands (Van der Stocken et al. 2019). However, Avicennia (Nettel & Dodd 2007; Lo et al. 2014) and in particular Rhizophora (Mori et al. 2015) have moved across the Atlantic and long distance dispersal events occured elsewhere, too; see also Rabinowitz (1978), Clarke et al. (2001) and Tomlinson (2017) for mangrove propagules, germination, and so on.
Note that broad-scale fossil and current distributions of some true mangroves seem to have little to do with each other, and the histories of individual mangrove species are complicated. It has been suggested that mangroves, perhaps even taxa that could be placed in extant genera, migrated along the shores of the Tethys 100-80 Ma ago (e.g. Nettel & Dodd 2007); subsequent vicariance and also long distance dispersal events help explain current distributions and patterns of genetic relationships (Lo et al. 2014). Nypa (Arecaceae, q.v. for fossils), today found only in the Indo-Malesian area, is first known from the Upper Cretaceous ca 70 Ma (estimated ages for the Nypa clade are 107-14 Ma - see Arecaceae) and by the Palaeocene ca 55 Ma it was growing in both the Old and New Worlds (e.g. Plaziat et al. 2001), and peat formed in Palaeocene Nypa-dominated mangrove habitats in Guyana (Leidelmeyer 1966). By the Eocene, ca 50 Ma, many mangrove genera are known from the fossil record, and several are known from both the Old and the New Worlds (Plaziat et al. 2001, but see below for Pelliciera). Avicennia-dominated stands have been found in early-Eocene Siberia, well to the north of other mangrove-type vegetation (Popescu et al. 2021); He et al. (2022) date Avicennia to just under 40 Ma. Fossil hypocotyls identified as Ceriops and preserved with good anatomical detail are known from the Lower Eocene London Clay (Wilkinson 1981), although Collinson and van Bergen (2004) noted that they did not show the distinctive curvature of seedlings of extant Rhizophoreae; He et al. (2022) date Rhizophoreae to somewhat under 50 Ma. Sonneratia may be Oligocene in age (Muller 1984); He et al. (2022) date it to over 40 Ma. At the other geographical extreme, Rhizophoreae are known from the Early Eocene 55-48.5 Ma in western Tasmania, Australia (Pole 2007), and this would seem to be the earliest record of the clade - and pretty much in conflict with the family age in Xie et al. (2012b). Rhizophora itself is known from the Caribbean in the late Eocene ca 50 Ma, but the common ancestor of the existing Caribbean populations probably arrived in the New World only ca 11 Ma (Graham 2006: much detail).
Pelliciera is an odd genus. He et al. (2022: Fig. 2b) thought that it had evolved over 60 Ma, although as a mangrove plant it is perhaps slightly less than 50 Ma, while mid-Eocene is the suggestion in Rull (2023a, b). Records of fossil pollen (see Lanagiopollis crassa, = Psilatricolproyes crassus) show that Pelliciera was once more widespread in the New World than it is now (A. Graham 1977; Rull 2023a), and it may even have grown in the Old World. However, the history of the genus is uncertain (see Martínez-Millán 2010: New World records, based on pollen, rejected - but Old World records are also based on pollen, see also Ellison et al. 1999; Plaziat et al. 2001; Manchester et al. 2015: potential confusion with pollen of Alangium-Cornaceae; Duke 2020: confusion with Hura-Euphorbiaceae). However, Rull (2023a) based his detailed scenario of changes in the range and abundance of Pelliciera in the New World from the Eocene onwards on the palynological literature. Rull (esp. 2022, 2023b) thought that the genus predated other mangroves there, and in the Eocene it was notably abundant where it grew. However, Rhizophora in particular became widespread and abundant in the Americas after the Eocene-Oligocene climate transition (cooling, pCO2 decrease, etc.) of ca 34 Ma while Pelliciera became very much less abundant, if somewhat more widespread than it had been, but then its range contracted and shifted (see especially Rull 2023a; also 2023b).
Changes in distribution continue. Not only has the extent of mangroves been severely recuced by human activiities, with reulant negataive effects on coastal protection, but mangroves have been expanding polewards in both hemispheres over the last 50 years or so, replacing salt marshes as they go - a response to climate change. This movement has been particularly pronounced in Avicennia germinans, the black mangrove (Saintilan et al. 2013). The biotic interactions between mangrove and marsh are quite complex and depend i.a. on whether the plant is a seedling or not and its elevation in the marsh (H. Guo et al. 2013).
Mangrove taxa have evolved parallel physiological and anatomical adaptations to the saline, tidal habitats in which they grow (e.g. Ball 1988; Parida & Jha 2010; Reef & Lovelock 2015 and other papers in Ann. Bot. 115(3). 2015), conditions in which are pretty extreme for land plants to live in. However, studies on New Guinea mangroves found rather greater variation in structural diversity (venation density, leaf mass/area, etc.) and functional performance (e.g. photosynthetic carbon assimilation rate) than might be expected (Balun 2011). Interestingly, above-ground growth of mangroves like Avicennia marina increased when non-saline water was available, e.g. from runoff, while below-ground growth was only indirectly affected, nutrient availability being important here (Hayes et al. 2018). Similarly, Rhizophora mangle grows well under moderately saline conditions (10 ppt NaCL) but not if the salinity is that of sea water (ca 35 ppt) or higher; from this point of view it behaves as a facultative halophyte (B. P. Silva et al. 2022). For salt and water balance, see Reef and Lovelock (2015) and other papers in Ann. Bot. 115(3). 2015. There has been convergent evolution in genes involved in stress response and embryo development in the mangroves R. apiculata, A. marina and Sonneratia alba, and in R. apiculata in particular there are expanded gene families involved in e.g. plant-pathogen interaction and the biosynthesis of secondary metabolites (S. Xu et al. 2017a, b). In both Rhizophora mangle and Avicennia root cortical phi [φ] cell wall thickenings are radially oriented (Collings et al. 2020) - extend the observations! Robert et al. (2009) discuss the hydraulic architecture of the wood of Rhizophora.
Toledo et al. (1995) found that nitrogen-fixing cyanobacteria were abundant on the roots of black mangroves, Avicennia nitida, in Mexico.
Mangroves tend to have large seeds/embryos, often considerably larger than those of their non-mangrove relatives (e.g. Moles et al. 2005a), that lack dormancy and are not infrequently viviparous, and this is connected with the need for fast establishment in the mangrove habitat - small seedlings could easily be washed away by the tides. In general, quick seedling establishment is facilitated by very fast germination, i.e. germination within one day of the start of imbibition (vivipary is simply an extreme), and this is especially common in Chenopodiaceae, many of which are other plants of saline habitats (see also Parsons 2012; Parsons et al. 2014; Kadereit et al. 2017).
The mangrove ecosystem is very productive and has high carbon flux rates, and it also stores much carbon, especially below ground - at about 1,000 Mg C ha-1, storage is about three times as much as in temperate or boreal forests or tropical upland forests, and if ca 900 Mg C ha-1 about half that of peatlands but more than other wetland vegetation; as with salt marshes in particular, a substantial amount of allochthonous C is trapped - ca 21 mm year -1 (Temmink et al. 2022). Pan et al. (2013) suggest figures of 218±173 Mg C ha-1, similar to peat swamps and much greater than in boreal forests (c.f Crowther et al. 2019). Mangroves occupy 13.7-15.2 million hectares (however, they have lost around 35% or more of their area in recent centuries - see Temmink et al. 2022), and they may store 4-20 PgC globally (Bouillon et al. 2008; Donato et al. 2011 and references). Other estimates are that they bury 17.0-23.6 TgCy-1, their gross primary productivity is 2087 gCm2y-1, global primary productivity is 417 TgCy-1, but with a rather lower net ecosystem production (221 gCm2y-1> and globally 44 TgCy-1) because of a relatively high respiration rate, at least when compared with sea grasses (Duarte et al. 2005: area estimated at 20 million hectares). The carbon in mangrove peat is usually very much older than that in adjacent vegetation, and that in the oldest deposists in a core from Kalimantan, Borneo, may be some 24,000 y.o. (Page et al. 2004). Note, however, that comments such as are made here belie the fact that the phrase "mangrove ecosystem" covers a multitude of sins. At the local scale there is variation not only in species composition, but tidal regime, the sediment composition and salinity of the seawater, etc.. Indeed, net and gross primary productivity and the like vary extensively between different kinds of mangrove habitat. Thus arheic (= no drainage channels) and karst/carbonate coasts have no or little submarine groundwater discharge respectively (Dürr et al. 2011), and although the latter in particular include less than 5% of the world’s total mangrove area, it stores almost 10% (213 Tg) of the global mangrove soil organic carbon (53.9 ± 1.6 mg cm3 - Twilley et al. 2018). And what about mycorrhizae? In an experiment, it was found that Bruguiera gymnorhiza formed associations with arbuscular mycorrhizal fungi if the water were fresh, rarely if it were brackish, but Rhizophora stylosa never did, and the latter is often found in more saline conditions than the former (Akaji et al. 2024, also references).
The rate of carbon accumulation and longer-term sequestration in mangroves and the extent of mangrove vegetation is likely to have been affected by recent changes in the sea level. Under stable sea levels, carbon accumulation is low, as it rises, accumulation increases in coastal wetlands in general, also, some C may be buried by fluvial and coastal processes and so sequestered, if sea levels fall, new areas become available for the establishment of such communities; if the rate of rise is too great, over 6 mm/year-1, then trees along the seaward side of mangrove vegetation may die, the mangrove community migrating inland (Rogers et al. 2019; Treat et al. 2019; Santilan et al. 2020). For the developing risk of CO2 emissions from mangroves, see Lovelock et al. (2017). There is other information about mangroves under Clade Asymmetries; see also other major peat-producing ecosystems like Sphagnum bogs, seagrasses and the somewhat less extensive dipterocarp forests.
Thinking about Rhizophoreae in particular, all members of which are mangroves and make up overall the most important component of mangrove vegetation, their seeds have little endosperm and are viviparous (aquatic/marine/mangrove plants commonly have large embryos, and in some the seed starts to germinate before it falls off the plant). In all genera except Bruguiera the endosperm overflows from the seed, pushing open the micropyle as it does so, and the hypocotyl/radicle of the germinating embryo dangles from the seed; the embryo minus cotyledons is the unit of dispersal. After the embryo falls from the tree it floats in the water, and after grounding the hypocotyl straightens, lateral roots develop, and the seeding becomes established (Juncosa & Tomlinson 1988b); for more on seeds, seedlings and dispersal, see Rainowitz 1978; Clarke et al. 2001; van der Stocken 2018). Depending on the genus, there are either stilt roots, plank roots, or pneumatophores (Gill & Tomlinson 1975). Axillary buds along the branches soon die so the plants cannot regenerate when cut or if the twigs are killed by frost, etc. (see Tomlinson 1986, 2017 for much useful information). Rhizophoreae are an example of the relatively uncommon situation in flowering plants where salt tolerance was acquired quite deep in the phylogeny, being retained since (Moray et al. 2015: ?6 origins).
Pollination Biology & Seed Dispersal. Pollen in Rhizophoreae is deposited on to the hairy petals, so there may be secondary pollen presentation. Pollination in Bruguiera and Ceriops tagal is explosive (Tomlinson et al. 1979; Juncosa 1988), the stamens being held in groups by the petals until the flower is tripped by the pollinator. Pollinators in this small tribe include birds and a variety of insects, and pollination mechanisms are diverse. The petals often have an arista or other appendages and are shaped like a tiny bivalve mollusc (Endress & Matthews 2006b). The pollen grains are very small, and in Rhizophora in particular pollination may be by wind (Juncosa & Tomlinson 1988b).
Genes and Genomes. S. Xu et al. (2017b) date a whole genome duplication in the family to ca 69 Ma and suggest that it is implicated in the origin/diversification of Rhizophoreae - note that 69 Ma is older that some estimates of the divergence of Erythroxylaceae and Rhizophoraceae. Cai et al. (2017/18) found that Rhizophora (Erythroxylum sister) had a genome duplication, while Pootakham et al. (2022) suggested Rhizophoreae as a whole had a genome duplication.
Interestingly, the genome size of true mangroves may be much smaller - about one third the size - than that of mangrove-associated and non-mangrove species, apparently because that part of the genome occupied by repetitive DNA, particularly by long terminal repeat retrotransposons, was smaller, in part because of a reduction in the rate of their origination (Lyu et al. 2017: focus on Rhizophora, Sonneratia and Avicennia, no comparison for the first given, see also Costa et al. 2024). They discuss this size reduction as occuring within the last ca 9 Ma as the plants invaded a stressful environment, although tese dates seem very recent compared with the age of the mangrove habitat.
In a survey of 14 mangrove plant plastomes, Su et al. (2019) found them to be largely unremarkable, although the authors noted a number of simple sequence repeats (61-130).
Chemistry, Morphology, etc.. Growth in a number of Rhizophoraceae may be continuous, although growth patterns in Macarisieae are unknown. Cork initation in the root is superficial in at least some taxa, perhaps just those with stilt roots (see von Guttenberg 1968 for Carallia), and their aerial roots are polyarch (Gill & Tomlinson 1975). The leaf teeth are theoid. The colleters of Rhizophoreae, at up to 1.5 mm long, are notably larger than those of other members of the family (Sheue et al. 2013: Paradrypetes not studied).
There is considerable variation in floral merosity in the family, both carpel and stamen number varying considerably (Matthews & Endress 2011). The stamens in polystemonous flowers arise from ring primordia (Ronse de Craene & Smets 1992b). Rhizophora has transversely arranged carpels (Eichler 1876). Variation in testal morphology in Gynotrocheae in particular is considerable, Gynotroches and Pellacalyx, with strongly exotegmic seeds, differing so much from Carallia, which lacks an exotegmen, that Corner (1976) preferred to segregate the former as Legnotidaceae - a comprehensive survey of seed anatomy in the family is desirable.
The morphology of the embryo of the mangrove species is interesting. In Rhizophora, at least, the radicle is deep-seated in origin, and in that genus and Ceriops it seems to be non-functional, the root system of the seedling being developed from axillary roots; Bruguiera does have a functional radicle (Kipp-Goller 1939; Juncosa 1982). The cotyledons of Rhizophora are connate when initiated (Juncosa 1982).
See also Juncosa and Tomlinson (1988) and Schwarzbach (2013), both general, Howard (1970: nodal anatomy), Baranova and Jeffrey (2006: leaf anatomy), Juncosa and Tomlinson (1987) and Juncosa (1988), both floral development, Endress and Matthews (2006b: petal morphology), Carey (1934) and Mauritzon (1939a), both embryology, Tobe and Raven (1987e, 1988b: seed coat anatomy) and Souto and Oliveira (2012: Banisteriopsis and Diplopterys, fruit anatomy); for information on Paradrypetes, see also Levin (1986, 1992) and Radcliffe Smith (2001 - as Euphorbiaceae).
Phylogeny. Schwarzbach and Ricklefs (2000: Paradrypetes not included) found strong phylogenetic structure in the family, with three major clades. The phylogenetic structure there is basically the same as that in later studies when Paradrypetes was included (e.g. M. Sun et al. 2016), indeed, molecular data have rather surprisingly placed Paradrypetes (ex Euphorbiaceae) here (e.g. Davis et al. 2005a), and it is strongly supported as sister to Cassipourea (Wurdack & Davies 2008: only one species from each tribe included). Paradrypetes has a rather unexpected combination of characters and is highly apomorphic (see above).
Within Rhizophoreae, relationships are [Bruguiera [Ceriops [Kandelia + Rhizophora]]]] (e.g. Schwarzbach & Ricklefs 2000; Lakshmi et al. 2002; S. Xu et al. 2017b), although Ceriops switched positions with Rhizophora in Pootakham et al. (2022).
Classification. Schwarzbach and Ricklefs (2000) suggested that three tribes be recognized for the three major clades that were apparent in their phylogeny of the family.
Previous Relationships. Rhizophoraceae have often been associated with Myrtales (Cronquist 1981) or Myrtanae (Takhtajan 1997), largely because of their vestured pits, opposite leaves, and inferior ovary, and they have sometimes also included or been closely associated with (Takhtajan 1997) Anisophylleaceae, here in Cucurbitales.
[Irvingiaceae + Pandaceae] / Pandoids: leaves on plagiotropic axes two-ranked; lamina vernation involute; flowers small; K connate basally; ovule 1/carpel, apical, pendulous, epitropous; fruit indehiscent; exotesta and endotegmen tanniniferous.
Age. The divergence of these families is estimated at (107.1-)91.2(-70.5) Ma in Xi et al. (2012b: Table S7). Although Davis et al. (2005a) did not recover this clade, the two families were both in isolated positions in his analysis, and he dated their stem-group ages to somewhere between 119-97.5 Ma.
Evolution: Divergence & Distribution. The rate of diversification of this clade - it contains ca 25 species - may have decreased (Xi et al. 2012b).
Economic Importance. Embryos of both Panda and Irvingia are rich in fats and are much used locally.
IRVINGIACEAE Exell & Mendonça - Back to Malpighiales
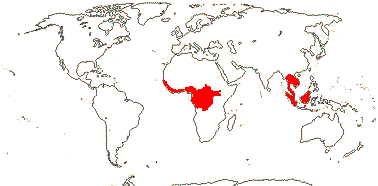
Trees; ellagic acid, myricetin +; vessel elements with simple perforation plates; nodes ?multilacunar; (sclereids +); stomata paracytic, veins vertically transcurrent; lamina margins entire, secondary veins strong, rather close and subparallel, tertiary veins also ± parallel and at right angles to the secondary veins, stipules intrapetiolar, deciduous; inflorescences racemose, branched, axillary or terminal; pedicels basally articulated; K cochlear; C protective in bud, cochlear or quincuncial, with 3 traces; A (9) 10, latrorse, filaments [and style] folded in bud; pollen ± triangular [polar view]; nectary massive, annular, vascularized from staminal traces; G [(2) 5], G median (when 2) or opposite sepals, style single, stigma subcapitate-papillate, ?type; cotyledons large, cordate; x = 7 (?6).
3 [list]/12: Irvingia (7) - two groups below. Africa, Madagascar; South East Asia to W. Malesia. Map: from Harris (1996). [Photo - Fruit]
Vessel/tracheid pits minute, half bordered; petiole bundles arcuate (plus adaxial-annular); fruit a septicidal/part loculicidal capsule with columella.
1/2. Madagascar, West Malesia.
2. Klainedoxa Engler + Irvingia J. D. Hooker
Secretory canals +; cristarque cells +; epidermal mucilage cells +; petiole bundle annular, (with inverted adaxial bundles in sheath); stipules very long and ensheathing terminal bud; ovules ± sessile, attachment broad, funicle thick, micropyle bistomal, outer integument 2-3 cells across, inner integument 3-4 cells across, parietal tissue 3-4 cells across, (nucellar cap +, weak), epidermis at nucellar apex with radially elongated cells, suprachalazal zone massive, placental obturator +, hypostase 0; embryo sac becomes long; fruit a 1-seeded berry, 1- or 5-seeded drupe with radial mesocarp fibres, or samara, K deciduous or not; seed with long hilum [Irvingia]; outer (esp.) and inner integuments multiplicative, testa vascularized, exotegmen fibrous/tracheidal, the rest ± collapsed; endosperm copious to 0; n = 13, 14, chromosomes 0.7-1.4 µm long; germination epigeal, phanerocotylar.
2/9: Irvingia (7). Africa; South East Asia to W. Malesia.
Evolution: Divergence & Distribution. Klainedoxa and Irvingia diverged (22.6-)11.8(-4.1) Ma (Xi et al. 2012b: Table S7).
Ecology & Physiology. Two species of Irvingiaceae (as Klainedoxa, Desbordesia [= Irvingia]) together made up over 6% of the above-ground biomass (a.g.b.) in eight Central African rainforests examined; they are two of the eighteen species that together make up half of the a.g.b., occupy numbers 2 and 4 on the list, with 3.6% and 2.7% a.g.b. and found at 7 and 3 sites respectively (Bastin et al. 2015).
Plant-Bacterial/Fungal Associations. For the mycorrhizal status of Irvingiaceae, see Bechem et al. (2014).
Chemistry, Morphology, etc.. Keller (1996) suggests that the leaves are involute in bud; this should be confirmed.
Forman (1965) described the seeds of Allantospermum as pulling away from a basal arilloid process. Netolitzky (1926) is unclear about exactly where the fibrous layer is in the seeds of Desbordea and Klainedoxa, suggesting that the seeds are exotestal, although Boesewinkel (1994) calls the fibrous layer exotegmic, which seems more likely.
See also Harris (1996: monograph) and Kubitzki (2013b) for general information, Forman (1965: Allantospermum), also Nooteboom (1967: esp. chemistry), Jadin (1901), Rojo (1968), van Tieghem (1905a: stomata anomocytic?), van Welzen and Baas (1986), all anatomy, Weberling et al. (1980: stipules), Link (1992c: nectary), and Tobe and Raven (2011: stamens, ovules and seed - Irvingia); details of floral orientation are taken from Eckert (1966) and fruit anatomy from Fernando and Quinn (1992).
Phylogeny. Relationships are [Allantospermum [Klainedoxa + Irvingia]] (Byng et al. 2016: support good).
Previous Relationships. All over the place, both in early molecular studies and in morphological studies - see introduction to the order. Prior to xii.2015, Allantospermum was in Ixonanthaceae...
PANDACEAE Engler & Gilg, nom. cons. - Back to Malpighiales
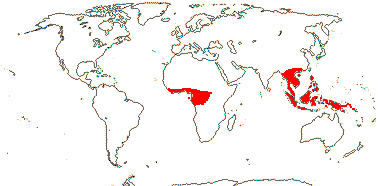
Trees to shrubs; chemistry?; cork?; vessels in radial multiples, vessel elements with scalariform (and simple - Galearia) perforation plates; rays 2-9 cells wide; sieve tubes with non-dispersive protein bodies; pericycle also with sclereids; druses and crystals +; petiole bundles D-shaped to (incurved-)arcuate; stomata various, cuticle waxes 0; leaves on orthotropic axes spiral, reduced, lamina with a single vein running into the opaque persistent tooth apex, one stipule higher than the other on the stem; inflorescences various; plant dioecious; K free to ± connate, C valvate or imbricate, usu. thick, hooded to flat; nectary 0; staminate flowers: A = and opposite K, 10, or 15, in one or two series, connective produced or not; pistillode +; carpelate flowers: staminodes 0; G [2-6], style 0, stigmas spreading, laciniate or entire; ovule (straight - Panda), outer integument 3-5 cells across, inner integument 3-5 cells across, nucellar cap ca 6 cells across, obturator 0/+; fruit a 2-5-seeded drupe, stone surface often irregular; exotegmen tracheoidal, (many layered - Panda); endosperm ?development, +, cotyledons incumbent, thin and flat, oily; n = 15, x = 7 (?6).
3 [list]/15: Microdesmis (10). Tropics, Africa to New Guinea (map: in part from Léonard 1961; van Welzen 2011; Trop. Afr. Fl. Pl. Ecol. Distr. 2. 2006). Photo: habit, fruit.
Age. Crown-group Pandaceae are (72.7-)47(-23.3) Ma (Xi et al. 2012b: Table S7).
Chemistry, Morphology, etc.. Panda smells like onions. Microdesmis has punctate leaves. The plagiotropic branches have been confused with compound leaves, especially in the derived Galearia and Panda; the stipules may be asymmetrically placed, as in Panda.
If the pedicels are articulated, they are articulated only at the very base.
For general information, see Forman (1966b), Radcliffe-Smith (2001), van Welzen (2011) and Kubitzki (2013b), also Hegnauer (1969: chemistry), Nowicke (1984) and Nowicke et al. (1998: pollen), Stuppy (1996) and Vaughan and Rest (1969), both seed anatomy, Tokuoka and Tobe (2003: ovules and seeds) and Hill (1937: germination of Panda) - mostly as Euphorbiaceae.
The embryology, etc., of the family are little known.
Phylogeny. What is known about wood anatomy suggests that Galearia and Panda are close, while pollen suggests that Galearia and Microdesmis are close (van Welzen 2011). The relationships [Microdesmis [Galearia + Panda]] are strongly supported by molecular data (see Xi et al. 2012b; M. Sun et al. 2016), in line with wood anatomical variation.
Previous Relationships. Pandaceae were included in Euphorbiaceae until quite recently, e.g. Govaerts et al. (2000) and Radcliffe-Smith (2001), but they differ from even the uniovulate taxa, Euphorbiaceae s. str., in several respects, including their indehiscent fruits. Rays of Euphorbiaceae are only 1-5 cells wide (Hayden & Hayden 2000); Pandaceae usually lack obturators, while Euphorbiaceae have them - another difference. Dicoelia (Euphorbiaceae - Dicoelieae) and Galearia both have stamens in depressions in the petals, however, Dicoelia has a low, thin-walled testa, a massive exotegmen, and a moderately thickened mesotegmen (Stuppy 1996), and belongs in Phyllanthaceae-Phyllanthoideae-Wielandieae (Kathriarachchi et al. 2005).
Engler had trouble with Panda, mistaking its strongly plagiotropic branches for compound leaves, so he described it first as a species of Burseraceae, then as a species in Sapindaceae, and after he recognized his mistake, he was still unclear as to its relationships and placed it in a monotypic Pandales (Forman 1966).
[Ochnaceae [[Bonnetiaceae + Clusiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]] / [Ochnoids + Clusioids]: biflavones +; indumentum poorly developed; stomata paracytic; leaves with colleters; C protective in bud, becoming widely spreading/reflexed, contorted [direction not fixed]; A many, centrifugal; pollen grains usu. small [<30µm in diameter]; nectary 0; (G [5+]); ovules many/carpel, parietal tissue 0, endothelium +; fruit a septicidal or -fragal capsule; endosperm at most slight.
Age. The crown age of this clade is (106.7-)101.5(-96.4) Ma (Xi et al. 2012b: Table S7), ca 100.9 Ma (Tank et al. 2015: table S1, S2), around 150, or even 185 Ma (Bissiengou et al. 2015b); (124.9-)120.6, 115.4(-104.2) Ma (Ruhfel et al. 2016: app. S9) and ca 166.7 Ma (L. Cai et al. 2025a) are other estimates.
Evolution: Divergence & Distribution. Several of the characters above are suggested as possible synaporphies for Ochnaceae by J. V. Schneider et al. (2014a). Optimization of characters like ovule number and styles free (= styluli)/fused is difficult. Xi et al. (2012b) describe the ovules of this clade as being "often incompletely tenuinucellate".
Ecology & Physiology. Lamina venation in this whole clade is interesting. Schneider et al. (2016), in their study of the venation of Ochnaceae, a family often with very close secondary veins and not much else or a very well developed and close reticulum, noted that the normal vein order/leaf size scaling relationships (for which, see Sack et al. 2012) in eudicots had in part broken down. Calophyllum (as C. longifolium, Calophyllaceae) also in this clade, is similar (Sack et al. 2012). Indeed, taxa with rather odd venation like closely parallel secondary venation are scattered throughout this family group, and include Neblinaria (= Bonnetia), no midrib at all, and other Bonnetiaceae, Endodesmia and Calophyllum (Calophyllaceae), some species of Clusia and Garcinia (Clusiaceae), and so on. Podostemaceae are of course vegetatively a law unto themselves.
Genes & Genomes. 7/23 genome duplications that Cai et al. (2017/18) recorded for Malpighiales as a whole are in this clade.
Chemistry, Morphology, etc.. It is quite common for the calyx to be small relative to the corolla even in bud (e.g. Matthews et al. 2012 for Ochnaceae), and here the corolla has taken over the protective function of the bud, although in taxa like Calophyllum and Clusia the bud is at first completely enclosed by the sepals. Pollen grains in general are quite often small, but there is substantial variation in the [[Bonnetiaceae + Clusiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]] clade (Furness et al. 2013b, c.f. 2012).
Classification. Van Tieghem (1902) thought that on balance Clusiaceae s.l. and Ochnaceae might be close, largely because polystemony occured in the former and in some of the latter.
OCHNACEAE Candolle, nom. cons. (Ochnoids in Xi et al. 2011b) - Back to Malpighiales
Pits vestured; mucilage cells/canals +; branching from previous flush; lamina with secondary and tertiary venation well developed; pedicels articulated; K with ≥3 and C with 3 vascular traces; (pollen with endexine thickened around apertures ["costate"]); gynophore +; micropyle bistomal, often zig-zag; K persistent in fruit; endosperm +; x = 14 (?13), nuclear genome [1C] (0.013-)0.706(-39.5) pg.
36 [list - to tribes]/650 - seven groups below. Tropical, esp. South America.
Age. Bell et al. (2010) date crown group Ochnaceae at (60-)45(-28) Ma; (43-)39, 36(-32) Ma is the estimate in Wikström et al. (2001: note topology) and (90.5-)77.8(-83.5) Ma in Xi et al. (2012b: Table S7). The age of the [Ochnoideae [Medusagynoideae + Quiinoideae]] clade is estimated to be (83.7-)71.4(-58.3) Ma (J. V. Schneider & Zizka 2017) or (81.1-)68.9(-57.2) Ma (Schneider et al. 2022).
1. Ochnoideae Burnett
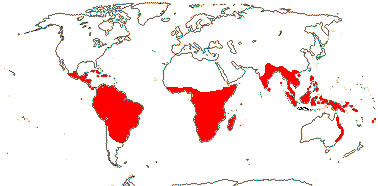
Isoflavonoids +; (vessel elements with scalariform perforation plates); vessel/parenchyma pitting unilaterally compound; nodes also multilacunar; cortical vascular bundles +; (sclereids +); pericycle of small isolated fibre bundles; petiole bundle annular, (several, arcuate); leaves 2-ranked; lamina with secondary veins strong and close, and/or with parallel tertiary veins, stipules fimbriate or not, (intrapetiolar); flowers 5-merous, monosymmetric in bud; K almost scarious; A (5-many), anthers (locellate), porose (not), filaments abruptly narrowed at anther junction/not, shorter than the anthers; androgynophore +, (short), G (1-)5(-15), opposite K, when 3 median member adaxial, style not branched, stigma ± punctate; ovule micropyle endostomal; antipodals persistent; filaments persistent in fruit, anthers abscising; ovules many/carpel; seeds winged; endotesta with small crystalliferous cells; (embryo curved).
28/495. Tropical, esp. Venezuelan Guayana (ca 1/4 the species) and Brasil. Map: from Kanis (1968, 1971), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003), Australia's Virtual Herbarium (consulted xii.2012) and Brummitt (2007: America).
Age. Crown-group Ochnoideae are estimated to be (74.6-)69.5(-63.5) Ma (Bissiengou et al. 2014a); the Testulea clade separated from other Ochnoideae ca 60 Ma (J. V. Schneider et al. 2022: Fig. 1).
1A. Testuleeae Horaninow - Testulea gabonensis Pellegrin
Lamina with secondary veins distant, brochidodromous, margin entire; bracteole 1; flowers 4-merous, monosymmetric; androecium adaxial, A 1, staminodes +, two thirds connate; G [2], placentation parietal; capsule inflated; n = ?
1/1. West tropical Africa.
[Luxemburgieae [Ochneae + Sauvagesieae]]: lamina with secondary veins closely parallel.
Age. The crown age of this clade is (72.4-)53(-38.9) Ma (Xi et al. 2012b: Table S7) or (57.4–)47.1(-36.7) Ma (J. V. Schneider et al. 2022).
1B. Luxemburgieae Horaninow —— Synonymy: Luxemburgiaceae van Tieghem
Leaves (sessile), venation closely parallel; monosymmetry evident in bud, oblique; A adaxial, anthers connate or not, porose, deciduous after anthesis, filaments ± connate, (staminodes +, small, outside fertile A); pollen exine with small perforations, ± smooth; placentation ± parietal, stigma commissural; (carpels pulling away acropetally and opening adaxially); G [3]; n = ?
2/22: Luxemburgia (18). Venezuela and Brazil.
Age. Crown-group Luxemburgieae are estimated to be (29.6-)20.1(-11.1) Ma (Bissiengou et al. 2014b).
[Ochneae + Sauvagesieae]: (pith chambered); (lamina with secondary veins distint); pollen with striate-rugulate exine; ovules (1-4/carpel).
1C. Ochneae Bartling —— Synonymy: Gomphiaceae Schnizlein, Lophiraceae Loudon
Root phellogen ± superficial [Ochna]; vessel/parenchyma pitting not unilaterally compound; (petiole with inverted medullary bundle and subepidermal fibres); leaves two-ranked, (stipules semi-intrapetiolar - Ouratea); flowers polysymmetric; K quincuncial, C contorted [?level], (inner edge of petal enveloping stamens in pairs); A (10, ob/diplostemonous), stamen development centripetal, (anthers dehiscing by long slits), (filaments longer than anthers); (pollen 3-celled); G [2-]5-10[-15], postgenitally connate [± apocarpous], style gynobasic, hollow or not, branched, transmitting tissue variously arranged, stigmas expanded (not); ovule one/carpel, (campylotropous), apotropous, integument single [= 2 fused, except sometimes at tip], 7-17 cells across, (outer integument 3-4 cells across, inner integument 2-3 cells across - Ochna), pachychalazal, vascularized by raphal bundles, hypostase + (/0?); embryo sac with antipodals enlarged; fruit indehiscent, nut-like (drupelet), (K enlarged, wing-like - Lophira), receptacle enlarged; seeds not winged; testa with vascular bundles, endotesta lacking small crystaliferous cells, fibrous exotegmen 0; endosperm 0; cotyledons massive, variously arranged, (unequal); n = 10, 12-14; germination commonly hypogeal.
9/500: Ouratea (inc. Gomphia: 310), Ochna (85), Campylospermum (41), Philomeda (13). Tropical, especially Brazil. [Photo - Flower, Flower, Fruit.]
Age. Crown-group Ochneae are estimated to be (48.4-)36.3(-22.5) Ma (Bissiengou et al. 2014b) or (45.2–)35.4(-25.5) Ma (J. V. Schneider et al. 2022).
Crown-group Ochninae are estimated to be (27.5–)20.9(-15.0) Ma (J. V. Schneider et al. 2022). Note, however, that fossil fruits identified as Ochninae have been found in North Dakota in deposits of Late Palaeocene age, perhaps around 58 Ma (Ickert-Bond et al. 2015b).
1D. Sauvagesieae de Candolle —— Synonymy: Euthemidaceae van Tieghem, Sauvagesiaceae Dumortier, Wallaceaceae van Tieghem
(Herbs); (medullary vascular bundles +); (colleters +); leaves spiral, (pinnately compound - Rhytidanthera), lamina vernation conduplicate-flat, (venation very closely parallel), base ± decurrent; monosymmetry developing late, involving A and G (flowers polysymmetric - Neckia, Sauvagesia, etc.); (K with outer members smaller than the rest), (C with 3 traces); (androecium with positional monosymmetry at anthesis), A 5 [opposite K], 10, (many, centrifugal), diplostemonous, (anthers deciduous after anthesis), (dehiscence apical or by long slits), staminodes +, ± petal-like [antepetalous staminodes forming a cone, contorted or connate], (0), (also many small spathulate coronal structures); (pollen exine with small perforations); G [2, 3, 5], when 3, median member adaxial, ovary finely ridged, (placentation parietal; laminar), style (0), stigma (porose), (shortly lobed), (lobes commissural); (ovules ≥2/carpel), micropyle bistomal/zig-zag, outer integument ca 2 cells across, inner integument ?3-4 cells across; (fruit a drupe); (seeds not winged); exotesta of large cells, ± detached, entotesta with crystalliferous cells; n = 19 [one count]; germination epigeal.
16/82: Sauvagesia (40). Pantropical, only 2 spp. in Africa, most South American.
Age. Estimates of the age of crown-group Sauvagesieae are similar - (49.4-)41.8(-29.7) Ma (Bissiengou et al. 2014b) and (49.3–)39.9(-30.5) Ma (J. V. Schneider et al. 2022).
[Medusagynoideae + Quiinoideae]: true tracheids +; leaves opposite; flowers often unisexual; anthers relatively short [2> x longer than broad], thecal septum massive, persistent; ovary with longitudinal ridges, styluli +, ovary roof well developed, stigmas expanded ["suction-cup-shaped"]; ovules 2/carpel, superposed, inner integument 3-4 cells across, nucellar endothelium +; K not persistent in fruit.
Age. The age of this clade is (89.1-)72.3(-54.9) Ma (Xi et al. 2012b: Table S7), (77.5-)64.7(-42.6) Ma (Bissiengou et al. 2014b) or early Eocene maybe around 54 Ma (J. V. Schneider et al. 2022).
2. Medusagynoideae Reveal - Medusagyne oppositifolia Baker —— Synonymy: Medusagynaceae Engler & Gilg, nom. cons.
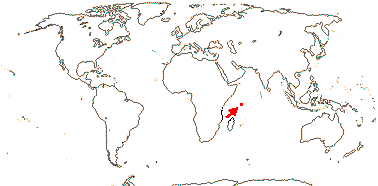
Plant tanniniferous; phloem stratified; nodes 5:5 + 2 phloic bundles; cristarque cells 0; petiole bundles many, arcuate, variously oriented; stomata anomocytic, cuticle waxes 0; plant glabrous; buds perulate; lamina venation very reticulate, stipules 0, colleters +; inflorescence terminal, ?cymose, plant andromonoecious; K basally connate; A spiral; orbicules numerous; pollen porate, exine protruding at the pores, surface finely striate, striae intertwined, onci +, basal layer of exine massive, endexine not lamellate, intine lamellate; G [16-25], adnate to massive central axis, ridged, ridges ± interrupted, without vascular bundles, stigma ?wet; ovules 2-5/carpel, when 2, one ascending, one descending, outer integument 3-4 cells across, inner integument 3-4 cells across, "weakly crassinucellate", funicles long; fruit verrucose, deeply ridged, carpels pulling away acropetally and opening adaxially, columella persistent; seeds winged; wings with several cell layers, exotesta slightly thickened; endosperm ?development, thin; n = ?
1/1. Seychelles, very rare.
3. Quiinoideae Luersson
Trees; mycorrhizae 0; cork?; (vessel elements with scalariform perforation plates); (silica bodies +); petiole bundle annular, often complex; lamina with strong, close secondary venation, tertiary venation closely parallel, at right angles to secondary veins, stipules pubescent; flowers small [5> mm in diameter]; (hypanthium +); K 4-5, pubescent, C 4-5(-8), usu. imbricate; A basally connate or not, (adnate to the base of the C), (subdorsifixed), thecae distinct; pollen exine with small perforations; (androgynophore 0); G (strongly ridged, ridges with vascular bundle), stigma type?; ovules basal, apotropous or epitropous, outer integument 4-7 cells across; fruit striate-somewhat ridged when dry, exocarp with lacunae; seeds 1-4, unwinged; coat?; endosperm development?; n = ?
4/48. Tropical America. Map: from J. V. Schneider et al. (2002).
Age. The crown age of this clade is (38.7-)18.1(-4.3) Ma (Xi et al. 2012b: Table S7), (27.2-)19.1(-11.7) Ma (Bissiengou et al. 2014b) or (57.3-)39.4(-22.7) Ma (J. V. Schneider & Zizka 2017).
3A. Froesia Pires
Plant ± unbranched, leaves in a rosette at end of stem [caulirosulate]; cristarque cells 0; phloem fibres 0; leaves compound, lamina margins entire; flowers perfect; G 3, free, stigmas punctate; ovules collateral; fruit follicular, one seed/fruit; endosperm 0, cotyledons massive.
1/5. Tropical South America.
Age. The crown age of Froesia is (20.6-)9.7(-1.8) Ma (J. V. Schneider & Zizka 2017).
3B. Quiineae Planchon & Triana —— Synonymy: Quiinaceae Engler, nom. cons.
(Lianas); phloem fibres +; (leaves compound), lamina margins entire to deeply lobed, (venation paxillate), stipules also interpetiolar, large, ± persistent; plant (cryptically) dioecious; (C with three traces); G [2-16], stigmas obliquely expanded; (ovules 1-4/carpel); fruit ± berry-like; seeds hairy; (endosperm 0, cotyledons massive).
3/43: Quiina (32). Tropical America. [Photo - Flower, Fruit.]
Age. The crown age of this clade is (42.7-)29(-16.3) Ma (J. V. Schneider & Zizka 2017).
Evolution: Divergence & Distribution. The diversification rate in Medusagynoideae may have slowed down (Xi et al. 2012b). Indeed, there is substantial clade size imbalance in Ochnaceae - [[1 [[4 + 18] [[5 + 77] [2 + 388]]] [1 [5 + 43]]].
The very long phylogenetic stem of Ochnaceae, over 70 Ma by some estimates, could reflect extinction at the K/P boundary. Bissiengou et al. (2014b) suggested that Ochnaceae originated in America (West Gondwana - J. V. Schneider et al. 2022), the ancestors of Medusagyne, now restricted to the Seychelles, perhaps getting where they did via migration over the North Atlantic land bridge via India (the ocean crust separating India and the Seychelles started to form ca 63.4 Ma - Collier et al. 2008). Schneider et al. (2022: see Fig. 2) suggested that most disjunctions in the family reflected Tertiary long-distance dispersal events, mostly west to east and transoceanic (dispersal in Sauvagesieae may have been westwards); this paper should be consulted for discussion about ages, details of distributions, etc..
A number of "features of systematic interest", "possible synapomorphies", etc., listed in bold by Matthews et al. (2012) have been placed tentatively at their appropriate hierarchical levels. J. V. Schneider et al. (2014a) suggest numerous apomorphies throughout the family, which I have tried to follow. However, I have not placed monosymmetric flowers as an apomorphy for Ochnoideae as a whole (with subsequent reversals), mainly because the flowers are monosymmetric in three different ways, so suggesting independent origins of this feature.
Ecology & Physiology. The relationship between vein density and stomatal density reflects an aspect of photosynthetic efficiency; venation construction costs are little affected. Species of Ochnaceae with the distinctive venation systems of Ochnaceae are common on the poor and sandy soils of the Venezuelan Guayana (J. V. Schneider et al. 2016).
Pollination Biology & Seed Dispersal. Buzz pollination is common/prevalent in Ochnaceae, and although the anthers have an endothecium, it is sometimes restricted to the area around the anther pore or pores. In Sauvagesieae a cone formed by petaloid staminodes closely surrounds the fertile stamens (their dehiscence varies from pores to longitudinal slits); pollen comes out of the apex of the cone, which functions as a pore, when the flower is buzzed (Kubitzki & Amaral 1991). There may have been reversal from dehiscence by pores to dehiscence by slits (Amaral 1980; Amaral & Bittrich 2004, 2013).
Vegetative Variation. The often very dense - and beautiful - venation of many Ochnaceae has attracted attention. Unlike the common relationships between the different levels of the venation hierarchy in eudicots (Sack et al. 2012), the secondary veins may be very numerous and close, and sometimes also very fine, or the reticulum in general may be close and well-developed, yet there is also reversal to more conventional venation (J. V. Schneider et al. 2016). The venation of the leaves of Quiinoideae is very distinctive, although not that dissimilar from that of other Ochnaceae, and it was studied in detail by Foster (1952 and references); veinlets ending free in the mesophyll can be few or even absent.
Genes & Genomes. There may be two genome duplications somewhere around here (Cai et al. 2017/18: Ochna the only species examined). Landis et al. (2018) suggested an age of ca 60.6 Ma for the OCMOα event, which involves the whole family.
Chemistry, Morphology, etc.. Sauvagesia lacks vestured pits; two other genera in Ochnoideae are recorded as having them (Jansen et al. 2001). Godoya has stratified phloem. There are mucilage cells or mucilage channels throughout the family that are to be found in various places in the plant, and the plants sometimes have a watery exudate. Rios et al. (2020) discuss leaf leeth and associated colleters in Ochnaceae (Luxemburgia).
There is considerable variation in floral morphology in Ochnaceae-Ochnoideae. Sauvagesia has numerous linear staminodes, five petal-like staminodes opposite the petals, and five stamens opposite the sepals, and the order of initiation of the floral parts is K, C, antesepalous A, G [ring primordium], antepetalous A [petal-like staminodes], numerous spathulate coronal structures (Farrar & Ronse De Craene 2013). The antesepalous primordia of Ochna (Ochnoideae-Ochneae) show centripetal development (Pauzé & Sattler 1978), while the androecia of members of the other tribes develop centrifugally (Amaral & Bittrich 1998). Zygomorphy is largely the result of the unequal development of the androecium rather later on, but in Philacra and Luxemburgia it is evident early in development (Amaral & Bittrich 1998). Lophira has unequally accrescent sepals, two members forming wings (there are only two carpels, each with many ovules, and the testa is thin). Placentation is often axile, but it can be laminar, as in Wallacea, or parietal, as in Schuurmansia (Amaral & Bittrich 2013).
There is also variation in the ovule, etc., of Ochna, alternatively, some reports must be incorrect. Chikkannaiah and Mahalingappa (1974) suggest that there is no endothelium, although the nucellar epidermis seems to take over that function (but see Endress et al. 2012). Batygina et al. (1991, p. 222) show Sauvagesia erecta as having a much enlarged endotesta with thick cell walls.
Matthews et al. (2011, esp. 2012) provide vast amounts of information on floral morphology for the family as a whole; for pollen, see Furness (2013b). For further information on Ochnoideae, see van Tieghem (1902: also embryo, 1904), Amaral (1991), Kanis (1968) and Amaral and Bittrich (2013), all general, Hegnauer (1966, 1989: chemistry), van Tieghem (1903c: root anatomy), Decker (1966: Luxemburgieae) and Dickison (1981), both anatomy, Ronse De Craene and Bull-Hereñu (2016: androecium), Baum (1951b: gynoecium), and L. L. Narayana (1975) and Guédès and Sastre (1981), both embryology.
In Medusagyne K and C initiate in a spiral sequence, the existence of stamen fascicles is debatable, and the ovules are initially parallel/collateral, but they become superposed (Ronse De Craene et al. 2017b); the upper ovules are ascending and epitropous, the lower ovules descending and apotropous (Batygina et al. 1991; Doweld 1998b). For a comparison of the fruit dehiscence of Medusagyne with that of some Ochnaceae, particularly some Sauvagesioideae, see Fay et al. (1997a); the anatomy of the fruits is similar to that of Caryocaraceae (Dickison 1990a).
Additional information on Medusagyne in particular is taken from Dickison and Kubitzki (2013: general), Beauvisage (1920: anatomy), Robinson et al. (1989: morphology), Dickison (1990a, 1990b: morphology and anatomy), Matthews et al. (2012: floral morphology) and Furness (2014: pollen).
The stomata of Quiinoideae are described as being paracytic by J. V. Schneider et al. (2002).
For general information, see Kubitzki (2013b) and J. V. Schneider and Zizka (2014, esp. 2016), for wood anatomy, see Gottwald and Parameswaran (1967), and for many details of floral morphology, see Matthews et al. (2012).
In both Quiinoideae and Medusagyne some of the ovules in each carpel abort, but both are particularly poorly known embryologically, etc..
Phylogeny. There has been good molecular support for a monophyletic Ochnaceae s.l. for some time, e.g. Fay et al. (1997a), Nandi et al. (1998), Savolainen et al. (2000a), Chase et al. (2002) and Korotkova et al. (2009), although relationships between the three main clades that make up the family have been less clear. However, Xi et al. (2012b: as families) found moderate (75% ML bootstrap; 1.00 p.p.) support for a [Medusagynoideae + Quiinoideae] clade, support is weaker in Wurdack and Davies (2009) and not terribly strong in J. V. Schneider et al. (2014a), it was also not that strong in the targeted enrichment study using nuclear genes of Schneider et al. (2020), perhaps a bit better in W. R. Baker et al. (2021, the Seed Plant Tree) and overall quite strong in the various analyses carried out by Shah et al. (2021a). Indeed, Schneider and Zizka (2017) found Medusagyne to be sister to the other two subfamilies, although support was weak; on the other hand, support was strong in the plastome analysis of Schneider et al. (2021).
The tribes of Ochnoideae and their relationships (as spelled out in the characterizations above) are all well supported in the study by Schneider et al. (2014a: 4 plastid loci + ITS, 2020: nucleome, 2021: esp. plastome; see also Bissiengou et al. 2014b; M. Sun et al. 2016, also the relationships recovered by W. J. Baker et al. 2021a, the Seed Plant Tree, sampling good). Shah et al. (2021a) compared the targeted dataset assembled by Schneider et al. (2020) with the Angiosperms353 dataset and also found overall good support. There is a certain amount of conflict between relationships suggested by nuclear and plastome data in Luxemburgia, Ochna and in particular Ouratea (Schneider et al. 2021).
Ochneae: Bissiengou et al. (2014a) examined relationships within Ochneae, and although support was sometimes not very high, Campylospermum may be polyphyletic (yes: e.g. J. V. Schneider et al. 2020, 2021; Shah et al. 2021a). Elvasia is morphologically very distinct: it has congenital carpel connation, an ovary with commissural lobes, terminal, shortly branched style with punctate stigmas, and non-vascularized integument, however, it is clearly embedded in Ochneae, while Lophira is sister to the rest of the tribe (see also Schneider et al. 2014a, 2020, 2021; Shah et al. 2021a). Groupings in Ochna at least in part corresponded to those in earlier work (see also Schneider et al. 2021), although the latter observed extensive incongruence between nuclear and chloroplast phylogenies; Shah et al. (2021b) suggested that there were numerous undescribed species here. Schneider et al. (2020) found some well-supported clades in Ouratea (polyphyletic - 2 species to be excluded), although they did not correspond to previous infrageneric groupings and were less well supported in plastome analyses (Schneider et al. 2021). Using Angiosperms353 and Ochnaceae-specific probe kits, Bissiengou et al. (2025: 59 samples of Ochneae-Ochninae) clarified generic relationships here, in particular, they confirmed the polyphyly of Campylospermum, part of it being sister to Brackenridgea while Campylospermum s. str. is sister to Rhabdophyllum...
Sauvagesieae. The excluded species of Ouratea, which were included in that genus quite recently, used to be in Indovethia and Neckia and indeed formed a small clade along with Euthemis, Indosinia and Schuurmansiella, clearly separate from Sauvagesia (Schneider et al. 2020) - these are in Sauvagesieae, and there are biogeographic implications. Sauvagesia itself is polyphyletic (Schneider et al. 2020; Shah et al. 2021). Reinales and Parra-O (2020: one nuclear and four plastid markers) found the relationships [Rhytidanthera [Godoya [Cespedesia + Krukoviella]]]; this group, along with Fleurydora, sister to the rest, was also evident in Schneider et al. (2020, 2021).
Within Quiinoideae, Froesia is sister to the other genera (Schneider et al. 2006, esp. 2014a, see also Schneider et al. 2002 for a morphological phylogeny; Wurdack & Davies 2009: Schneider & Zizka 2017). Other relationships are [Quiina [Touroulia + Lacunaria]] (Schneider & Zizka 2017).
Classification. Van Tieghem (1904), emphasizing variation in characters of the embryo, etc., recognized numerous genera now synonymized under Ouratea and Ochna. Including Ochnaceae, Medusagynaceae and Quiinaceae in Ochnaceae s.l. is an optional arrangement in A.P.G. II, and they have much in common; Ochnaceae s.l. are recognized in A.P.G. III (2009). The tribal classification above is that of J. V. Schneider et al. (2014a, 2021a, etc.); they also describe subtribes. Rhabdophyllum (Ochneae) is polyphyletic, part now residing in Philomeda (Bissiengou et al. 2025). For sections in Ochna, see Shah et al. (2021b).
Previous Relationships. Diegodendron was included in Ochnaceae by Cronquist (1981), it is here placed in Malvales as Diegodendraceae (see also Amaral 1991).
Medusagyne is morphologically very distinctive. Comments on the species cover of Medusagyne at the Royal Botanical Gardens, Kew, ca 1985: "c.f. Actinidia. - Would be much better placed in Guttiferae or Hypericaceae - !!!!! - this plant allied to Myrtales. - Nonsense! - oh yes it is!" Hardly surprisingly, it was placed in a monotypic Medusagynales (Theanae) by Takhtajan (1997) and generally associated with Theales (e.g. Cronquist 1981); the latter was already such an heterogeneous group that the inclusion of practically anything else made little difference to its description.
Thanks. For discussion, and for comments on relationships within Ochnoideae, I am grateful to Maria Amaral and Volker Bittrich.
[[Bonnetiaceae + Clusiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]] / Clusioids: flavones, flavonols, (ellagic acid), prenylated xanthones and dimeric xanthones, polyisoprenylated benzophenones [benzophenone = (C6H5)CO(C6H5)], acylphloroglucinol derivatives, quinones +; vessel elements with simple perforation plates; schizogenous resin canals or cavities + [exudate colour varies]; nodes 1:1; cristarque cells 0; leaves opposite, lamina margins entire, stipules 0; inflorescence cymose; A fasciculate, fascicles opposite C; G opposite K [check], or median member adaxial; micropyle exostomal; exotegmen with low, lignified, sinuous anticlinal cell walls; embryo ± fusiform.
Age. This node can perhaps be dated to the Cenomanian (104-)94, 89(-87) Ma (Davis et al. 2005a: note topology), (91-)89.5(-88.4) Ma (Xi et al. 2012b: Table S7), around 148-110 Ma (Bissiengou et al. 2015b), (116.9-)110.6, 102.9(-92.3) Ma (Ruhfel et al. 2016: app. S9), ca 90.1 Ma (L. Cai et al. 2025b: Bonnetiaceae not sampled), or as little as 54-45 Ma (Wikström et al. 2001).
Evolution: Divergence & Distribution. Ruhfel et al. (2016) looked at distribution patterns in the whole clade, observing that most of the numerous disjunctions they observed, some forty nine in all (for example, there were no fewer than eleven dispersals to, but none from, Madagascar), were best explained by dispersal rather than Gondwanan-age vicariance-type events.
Estimates of clade ages here depend on where the problematic fossil Paleoclusia (see below) is placed, whether as the most recent common ancestor of Clusiaceae or that of [Bonnetiaceae + Clusiaceae] (Ruhfel et al. 2013; for similarities of its pollen with that of Tovomita and relatives, see Marinho et al. 2015). Estimated ages using the former position were older, indeed sometimes, as with the crown-group age of Clusiaceae, age estimates using different calibrations showed no overlap (Ruhfel et al. 2013, 2016) - and of course Paleoclusia may not be a member of Malpighiales at all.
There are several potential morphological synapomorphies for the clade (see Ruhfel et al. 2013 for some ancestral state reconstructions). Variation in seed/diaspore size is considerable (see also Moles et al. 2005a), as is that of embryo morphology.
Genes & Genomes. A genome duplication involving Garcinia, Hypericum and Mammea, the GRLIα event, has been dated to ca 86.3 Ma (Landis et al. 2018).
Overall there is rather little plastome variation in this clade, although the LSC of Mahurea exstipulata (Calophyllaceae) is about 10,000 bps longer that that of the other taxa (12 total) examined, and the IR some 10,000 bps shorter, while the LSC of the two Podostemaceae included was ca 5,000 bps shorter and the IR ca 3,000 bps shorter.
Chemistry, Morphology, etc.. For a summary of the chemistry of the clusioids, see Crockett and Robson (2011); exactly where on the tree some classes of secondary metabolites are to be placed will depend on more detailed sampling. Xanthones are uncommon elsewhere in seed plants, although they are known from Gentianaceae, some Moraceae, etc.. The xanthones of Podostemaceae are similar to those of both Gentianaceae (in their -6-0-glucosides) and Clusiaceae (in their isoprenyl substitutions); for polyisoprenylated benzophenones, see also Ferraz et al. (2020) and Cabral et al. (2021).
Prado and Demarco (2018) discuss secretory resin ducts lined by epithelium in Calophyllaceae and Clusiaceae; the contents may be white, and so look like latex. See Costa et al. (2021) for the nature of exudates in Kielmeyera (gums, resins) which depends on where in the plant the canals are (secondary phoem, cortex/medulla); this work should be extended throughout the clusioids.
Endress et al. (2013) suggested that that the sepals may be preceded by several bracts; inflorescences in this group seem to be basically cymose. Furness (2012) summarized the palynolgical variation - considerable - in the clusioids; some characters were optimised on an outline tree, but there was not much obvious phylogenetic signal. De Almeida et al. (2023b) discuss the possibility that the connective glands known from Hypericaceae, Clusiaceae and Calophyllaceae and secreting waxes or resins might be a high-level synapomorphy somewhere around here; however, general sampling is unclear, but they are absent in Podostemaceae and apparently unknown for Bonnetiaceae.
Phylogeny. Morphological data in particular (see many of the features immediately above) initially seemed to suggest a grouping of [Elatinaceae + Bonnetiaceae + Clusiaceae/Hypericaceae], seed anatomy and gross morphology of Elatinaceae and some Hypericaceae in particular being similar (e.g. see versions 4 and earlier of this site). This was not a monophyletic group in Savolainen et al. (2000a), indeed, Ploiarium was there even placed in Malvales (but see Wurdack & Davis 2009), although testa anatomy, etc., are strongly against such a position. Analyses in Chase et al. (2002) weakly linked Elatinaceae and Bonnetiaceae with Clusiaceae + Podostemaceae. Evidence now suggests that Elatinaceae are sister to Malpighiaceae (Davis & Chase 2004; Davis et al. 2005a; Tokuoka & Tobe 2006; Wurdack & Davis 2009), and some morphological data support this. Bonnetiaceae link with Clusiaceae s.l., i.e. Clusiaceae s. str., Hypericaceae and Calophyllaceae, in morphological phylogenetic analyses (e.g. Luna & Ochoterena 2004: Hypericaceae not included). Les et al. (1997a) had early linked Podostemaceae with Hydrostachyaceae (see Cornales), although this may in part have been a sampling probem; no Malpighiales, etc., were included; Podostemaceae are so highly modified that prior to the advent of methods for determining relationships using molecular data they might have been put practically anywhere. For more discussion on relationships around here, see above.
Classification. The old Clusiaceae (see versions 8 and before) were strongly paraphyletic. Keeping the genera of the old Clusiaceae in a single family, but making that family monophyletic, would entail the inclusion of Bonnetiaceae, Hypericaceae and Podostemaceae - and the latter has one of the most distinctive morphologies of all flowering plants. For the "price" of recognizing Podostemaceae, we have five coherent, mostly middle-sized and moderately to very easily recognizable clades.
[Bonnetiaceae + Clusiaceae]: root cork superficial; ovule suprachalazal zone long [ca 2/3 length of ovule]; embryo hypocotyl/radicle long [cotyledon:hypocotyl + radicle ratio ca 0.2>].
Age. The age for this node is (89.6-)82.1(-69.6) Ma (Xi et al. 2012b: Table S7), around 93.5 Ma (Tank et al. 2015: table S2) or (114.6-)106.6, 91.5(-89.8) Ma (Ruhfel et al. 2016: app. S9).
BONNETIACEAE Nakai - Back to Malpighiales
Shrubs; anthraquinones +, polyisoprenylated benzophenones 0, biphenyls, biflavones 0; (nodes 3<:3<); schizogenous cavities 0 [plant lacking exudate?]; hypodermal mucilage cells +; plant glabrous; leaves spiral, lamina vernation supervolute, (venation parallel), vascular bundles with endodermis + Casparian strip, with discrete patches of phloem, (abaxial surface glands +), (margins minutely toothed by setae), petiole short/0; (androecium not obviously fasciculate - Bonnetia); tapetal cells binucleate; G [3-5], style long, hollow, or style branches ± separate, stigma surface rounded-papillate; ovule bistomal, outer integument ca 3 cells across, inner integument ca 2 cells across; (cotyledon:hypocotyl + radicle ratio to 0.5); n = 11 [Ploiarium], ca 150 [B. cubensis].
3[list]/35: Bonnetia (30). Cambodia, Malesia (mostly Western), Cuba, South America. [Photo - Flower, another Flower.]
Age. The crown age of this clade is (66.8-)52.6(-35.7) Ma (Xi et al. 2012b: Table S7) or (86-)59.2, 52(-31.7) Ma (Ruhfel et al. 2016: app. S9).
Evolution: Genes & Genomes. For chromosome numbers, see Oginuma and Tobe (2013).
Chemistry, Morphology, etc.. Given the likely phylogenetic relationships above, anatomical studies of Bonnetiaceae are needed to clarify the apparent absence - or near absence - of secretory tissues here. Takhtajan (1993) did describe the pith of Bonnetiaceae as having secretory canals, as in Clusiaceae, but c.f. Baretta-Kuipers (1976); the recently-described Bonnetia ayangannensis has small glands of unknown nature on the abaxial surfaces of its leaves and sepals (Barbosa-Silva et al. 2024).
Bonnettia s.l. has tri- or multilacunar nodes, a mucilaginous epidermis, a foliar endodermis, and foliar sclereids; Archytaea and Ploiarium have unilacunar nodes and lack the mucilaginous epidermis, foliar endodermis and sclereids (Dickison & Weitzman 1996). Keller (1996) described the leaf vernation as being involute.
Bonnetia often has 3-trace petals while the bracetoles sometimes have only a single trace (Dickison & Weitzman 1998). For Archytaea, Wawra von Fernsee (1886) shows a floral diagram in which both the five carpels and the stamen fascicles are drawn opposite the sepals.
For a general account, see Weitzman et al. (2006), for chemistry, see Hegnauer (1969, as Theaceae) and Carvalho et al. (2013: Bonnetia), for anatomy, see Beauvisage (1920), and for some embryology, see Prakash and Lau (1976).
Phylogeny. Relationships are [Bonnetia [Archytaea + Ploiarium]] (e.g. Xi et al. 2012b; Ruhfel et al. 2016).
Previous Relationships. Savolainen et al. (2000b) found that Ploiarium was placed within Thymelaeaceae, but morphologically and anatomically this position would seem rather unlikely - probably the leaf was from Gonostylus (Thymelaeaceae), which grows in the same area.
CLUSIACEAE Lindley, nom. cons. / GUTTIFERAE Jussieu, nom. cons., nom. alt. - Back to Malpighiales
Trees or shrubs; isoflavones, diterpenes; (vessel elements with scalariform perforation plates); exudate usu. in (branched) canals; petiole bundle arcuate to annular; lamina vernation often flat (conduplicate), margins entire, (intrapetiolar hood-shaped structure at base); flowers (3-)4-5(-8)-merous; K and C usu. decussate; A (5-), connate or not, (anthers extrorse), (with small glands), filaments as stout as anthers; G [2-5(-16)], often opposite petals, style short [shorter than ovary], stigmas expanded, wet; seeds few-many, large; embryo chlorophyllous or white, cotyledons minute [cotyledon:hypocotyl + radicle ratio <0.1]; x = 7 (?6).
14/930: [list: tribes] - three tribes below. Throughout the tropics. Map: from Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1. (2003), Gustaffson et al. (2007) and Fang et al. (2011).
Age. The spread of crown ages of this clade is (72.4-)52.9(-30.4) Ma (Xi et al. 2012b: Table S7), around 51-42 Ma (Bissiengou et al. 2015b), or as much as (92.8-)91.1, 63.6(-52.2) Ma (Ruhfel et al. 2016: app. S9).
An interesting and well-preserved late-Cretaceous fossil in Turonian deposits ca 90 Ma, Paleoclusia chevalieri, from New Jersey, U.S.A., is possibly assignable here and so may affect the accepted age of the clade (Crepet & Nixon 1998; see also Friis et al. 2011; Ruhfel et al. 2013). The seeds are described as being arillate, but the position of the aril is unlike that of extant Clusiaceae (it is adjacent to the seed, rather than surrounding it, anf it and the testa s. str. have a similar external morphology), and it may be an aborted seed (Ruhfel et al. 2013). Crepet and Nixon (1998), in an analysis using 61 morphological characters from 35 extant genera, linked it withGarcinia and Clusia, not Vismia and Hypericum, together a member of the poly(11-)tomy they recovered. Ruhfel et al. (2013) also recovered a poly(35-)tomy in their analysis of 69 morphological characters, only 16 also used by Crepet and Nixon (1998); although Paleoclusia was not included in this analysis, Ruhfel et al. also linked it with Clusiaceae, not Hypericaceae. Marinho et al. (2015) noted that its pollen was quite like that of Tovomita and relatives, but if its seeds were not arillate... Schönenberger et al. (2020) found that most maximally parsimonious positions for this fossil were in crown-group Clusiaceae and Hypericaceae, while it ended up in a variety of rosid orders, Sapindales and Malvales being most common, in the analyses by López-Martínez et al. (2023a: Table 3). Its use in calibrating phylogenies around here is discussed briefly above.
1. Clusieae Choisy
Plants (lianes), (epiphytes), (CAM photosynthesis +); (hypodermis +, often multiseriate - Clusia); petioles joined by a line at the node [?level]; plant dioecious; inflorescence terminal (axillary), pedicels articulated (not); (outer K protective in bud), (C 0); androecium not obviously fasciculate, (anthers locellate), (sporangia annular); pollen (in tetrads); ovary with a roof, styluli separate; ovules 1-many/carpel, (micropyle bistomal), outer integument 2-6 cells across, inner integument 3-7 cells across; seeds often ≤3 mm long, arillate, aril vascularized or not; testa and tegmen somewhat multiplicative; n = 30; germination phanero(crypto-)cotylar, epigeal (hypogeal).
5/480: Clusia (300-400), Tovomita (62), Chrysochlamys (55), Arawakia (18). New World tropics. Photo: staminate flower, fruit.
Age. Crown-group Clusieae are (32.8-)21.7, 20.5(-11.4) Ma (Ruhfel et al. 2016: app. S9).
[Garcinieae + Symphonieae]: (paired "stipular glands" on the stem); pollen at least 4-aperturate; style +; fruit indehiscent, baccate; (testa and endocarp ± fused); seed coat complex, outer integument vascularized, multiplicative, exotegmen usu 0; germination crypto(phanero-)cotylar, hypogeal (epigeal), radicle reduced (not).
Age. The crown age for this clade is around (56.6-)49.3, 44.1(-42.3) Ma (Ruhfel et al. 2016: app. S9).
2. Garcinieae Choisy - Garcinia L. —— Synonymy: Cambogiaceae Horaninow, Garciniaceae Bartling
(Buds perulate); plant dioecious; A free/variously cunnate/fasciculate, (filaments thinner than anthers); placentation basal or parietal, style usu. short; ovule 1 (many)/carpel, apotropous, outer integument 4-8 cells across, inner integument 2-3 cells across, (outer integument ca 10 cells across, inner integument ca 22 cells across - sect. Allanblackia), (integument single, 18-20 cells across), (parietal tissue ca 5 cells across), (nucellus protruding up micropyle), suprachalazal zone unremarkable; (fruit septicidal); (exotegmen +); n = 22, 24, 27-29, etc.; an adventitious root developing at the base of the plumule [the main root of the seedling, = Garcinia-type germination].
1/400. Tropical, esp. Old World.
Age. The age of crown-group Garcinieae is some (31.3-)21.1, 9.8(-11.5) Ma (Ruhfel et al. 2016: app. S9).
Garcinioxylon tertiarum, fossil wood from the Deccan Traps, India and 67-65 Ma, has features unique to woods of extant Garcinia (S. Y. Smith et al. 2015), while fossil flowers, Eogarcinia longistaminata (there are also flower buds and fruits) have recently been described from Eocene deposits ca 55-52 Ma in Rajasthan, India (Ali et al. 2026c).
3. Symphonieae Choisy
Buds perulate; (flowers single); C contorted; anthers extrorse, (2-)5-40 mm long, much longer than broad; style (relatively long), branched (not), stigma porose; ovules 4-8/carpel, (micropyle endostomal), outer integument 10-22 cells across, inner integument 10-15 cells across; testa vascularized [Platonia], (exotegmen massively developed); n = 28, ca 29; embryo with plumule, radicle and adventitious root all developing together [Symphonia].
7/48: Symphonia (23). Tropical, few mainland Africa.
Age. Crown-group Symphonieae are around (46.6-)44.3, 44.1(-42.3) Ma (Ruhfel et al. 2016: app. S9).
Pollen about 54 Ma from West Africa has been identified as being that of Symphonia (Dick et al. 2003; Dick & Pennington 2019).
Evolution: Divergence & Distribution. It has been suggested that the around 750 species in Garcinieae and Clusieae, about 93% of the whole family, have diverged within the last 25 Ma or so (Ruhfel et al. 2016: Fig. 1). However, as is evident above, the ages of some fossil pollen and wood are very considerably older, and Ali et al. (2026c) suggest that (the ancestors) of Garcinia were members of the flora that inhabited India as it drifted north following the split of Gondwana...
Indeed. Symphonia, currently known from tropical South America, Africa and Madagascar (most species are from the latter area) has a pollen record dating back to the Eocene ca 54 Ma, and it may have achieved its current range by long distance dispersal (Dick & Pennington 2019).
Ecology & Physiology. In the New World, the speciose Clusia includes epiphytes and stranglers many of which have more or less succulent leaves, and they grow at elevations up to 3500 m (Gustafsson et al. 2007). Furthermore, a number of species in the C. minor and C. flava groups (see Gustafsson & Bittrich 2002) have strong to weak crassulacean acid metabolism (CAM) (Holtum et al. 2004; Lüttge 1996, 2008; Luján et al. 2023 for a summary). CAM has evolved twice or more in species of Clusia from Panama alone (Gehrig et al. 2003), CAM has also been lost several times (Gustafson et al. 2007; Silvera et al. 2010b) - note, however, that Luján et al. (2022) suggested that whether or not CAM was ancestral here depended on the analysis. In species like C. pratensis CAM is facultative (Winter & Holtum 2014; Lüttge 2008), and this is common in the genus. The development of CAM may be promoted by phosphorus deficiency; the epiphytic habitat is not rich in phosphorus, and terrestrial Clusia often grow on poor soils; whether or not the plant is mycorrhizal also affects the plant's phosphorus and carbon metabolism (Maiquetía et al. 2009). CAM here may be an adaptation to increase water use efficiency in the seasonally dry habitats some species favour; it does not occur in species growing in cloud forests (Luján et al. 2022). Barrera Zambrano et al. (2014) discussed CAM and C3 photosynthesis in Clusia in the context of its leaf anatomy which, however, did not seem to be very distinctive. On the other hand, Luján et al. (2022) found that CAM species tended to have thick palisade tissue and large palisade cells, overall, CAM increases C gain buffering against leaf water loss during periods of drought (see also Leverett et al. 2022/2023a). Leverett et al. (2021) found that sustained water limitation favoured obligate CAM, while periods of acute drought favoured species with facultative CAM. CAM requires large palisade chlorenchyma cells, whereas hydraulic capacitance (CFT) is governed by the presence of echlorophyllous hydrenchyma tissue (it is in the hypodermal position), and this latter may be an adaptation to cloud cover at higher altitudes (Leverett et al. 2022/2023a). Constitutive CAM was correlated with veinlet and vein terminus densities when standardized against leaf thickness, moreover, the ratio intervein distance:vein to lower epidermal distance is 1>, probably to provide additional water to the mesophyll and to keep the leaf hydrated when the stomata are open (Leverett et al. 2023b). Perhaps surprisingly, CAM is independent of the turgor loss point (TLP) and thickness of water-storing hydrenchyma tissue, although opening stomata at night reduces water loss (Leverett et al. 2021). Overall, species of Clusia are very flexible both in their habit and in the degree to which CAM photosynthesis is expressed. Clusia, always woody, shrubby to tree-like, and often an inhabitant of lowland tropical rainforests, differs from most other CAM species which are small, either epiphytes (of course, Clusia is often an epiphyte) or plants of dry conditions, and which are quite often annuals. Luján et al. (2023) summarize CAM research in Clusia. For major foci of CAM photosynthesis in angiosperms, see Orchidaceae, centrosperms, etc.; Lüttge 2008).
Clusia hilariana in particular, a plant of Brazilian restingas, is an important nurse tree, a variety of other species germinating and growing in its shade; seedlings of C. hilariana itself are often associated with tank bromeliads (Dias & Scarano 2007). For more on the general ecology of Clusia, see papers in Lüttge (2007).
Garcinia is one of the five most diverse genera in West Malesian l.t.r.f. (Davies et al. 2005); its members are mostly rather small trees.
Pollination Biology. Bittrich et al. (2006) summarize information about the role of oils, resins, etc., in the pollination of the family; see also Amaral et al. (2017). Variation in the androecium in Garcinia and Clusia in particular is extreme. Stamens in Clusia may be fused or free, the filaments are often massive, with canals in which resins are produced and a vascular system that can be quite complex, sometimes forming a ring (Sá-Haiad et al. 2015), the anthers can be locellate and thecate or athecate, or there is an outer annular sporangium surrounding a small central spherical sporangium (C. valerioi: Hochwallner & Weber 2006), and so on (Amaral et al. 2017 and references). The endothecium is locally absent to about three cell layers across, and this affects how the anther dehisces (see also Sá-Haiad et al. 2015; Amaral et al. 2017). In Clusia, resins (almost pure polyisoprenylated benzophenones mixed with fatty acids) are a common floral reward (Porto et al. 2000; Nogueira et al. 2001), and plants may also produce oils so reducing the viscosity of the resins (Porto et al. 2000). Floral resins are involved in pollination in about half the genus, as well as in the Clusia look-alike, Clusiella (Calophyllaceae) (Gustafsson & Bittrich 2002; Bittrich et al. 2006; Gustafsson et al. 2007; Sá-Haiad et al. 2015). Floral resin production has evolved three or four times or so here, but it has also been lost at least twice (Gustafsson et al. 2007). Depending on the species, resin and pollen may be mixed or presented separately (Amaral et al. 2017). Euglossine and especially stingless Trigona (meliponine) bees have been observed at Clusia flowers (Bittrich & Amaral 1986; Porto et al. 2000; Bittrich et al. 2006; R. L. Martins et al. 2007; Amaral et al. 2017), while the cockroach Amazonia platystylata pollinates C. aff. sellowiana (= C. blattophila) (Vlasáková et al. 2008, 2019). Interestingly, species of Clusia growing at higher altitudes, where bees are less common, produce nectar as a floral reward (Armbruster 1984). In general, resins are an uncommon floral reward in angiosperm flowers (e.g. Tölke et al. 2019), but see also Dalechampia (Euphorbiaceae) and Maxillaria (Orchidaceae). For details of the androecial morphology of Garcinia and its immediate relatives, see Sweeney (2008, 2010), Leins and Erbar (1981) and Leal et al. (2012); the nectary is unlikely to be staminodial. Little is known about pollination in that genus.
In the American Symphonia globulifera pollen is suspended in a distinctive oil which is picked up by hummingbirds and then deposited in a droplet that exudes through the pore at the tip of the stylar branches; the droplet and the pollen it contains is then sucked back into the pore (Bittrich & Amaral 1996; Bittrich et al. 2013). All other Symphonieae have similar stigmas, and so the pollination mechanism is likely to be the same throughout the tribe; bird pollination is known in other American Symphonieae like Moronobea and Platonia (Bittrich et al. 2013 for references).
Fragrant oils are produced in the stout filaments of the flowers of Tovomita, and these attract male euglossines. The compositions of the fragrances in three different species growing in the Ducke Nature Reserve were found to be quite different (Noguiera et al. 1998).
Goldberg et al. (2017) discussed the evolution of breeding systems in Garcinia.
Plant/Animal Interactions. Clusia is an important food source for Neotropical birds (Messeder et al. 2020a).
Genes & Genomes. Perhaps two genome duplications can be associated with the ancestor of this clade (Cai et al. 2017/18: no Bonnetiaceae examined).
Chemistry, Morphology, etc.. The distinctive exudates of Garcinia have been called xanthics (Lambert et al. 2013). For resins in Clusiaceae, sometimes called latex, see e.g. Porto et al. (2000), Bittrich et al. (2006) and Alencar et al. (2020); whether the benzophenones are polyisoprenylated or both polyisoprenylated and benzoylated may have taxonomic significance in Clusia. Anholeti et al. (2015) discussed the distribution of polyisoprenylated benzophenones in the context oif the sectional breakdown of the genus.
Roots of Clusia, at least, may have superficial phellogen, as is fairly common in other epiphytic taxa. Genera like Dystovomita and Garcinia have distinctive basal intrapetiolar hood-shaped structures whose development needs study; see Cruz et al. (2015) for the stipular nature of apparently similar structures in Metrodorea (Rutaceae). The resin-containing canals of Clusiaare schizogenous in origin and lined with epithelium, the resin being secreted in various ways - eccrine, granulocrine, or holocrine (K. M. M. Silva et al. 2019; Alencar et al. 2020). For colleters, see Silva et al. (2019).
Hochwallner and Weber (2006) described the androecium of Clusia valerioi as being fasciculate, but this is not obvious from the illustrations. Puri (1939), Leal et al. (2012) and Tobe and Raven (2011) thought that the ovules were bitegmic while Corner (1976) and Asinelli et al. (2011) described them as being unitegmic. Variation in ovule, seed and fruit in Clusiaceae in general is considerable and poorly understood.
In the germination of Garcinia, at least, the radicle may or may not develop, but in the latter case an adventitious root and plumule arise from the same end of the embryo; removal of the radicle from the seedling may have little effect. Indeed, it is common for an adventitious root to develop at the base of the seedling, whether or not a radicle also develops, and in some cases adventitiious root, radicle, and developing seedling all appear at the same end of the embryo - and also in Symphonia globulifera (Jaganathan et al. 2024 and references); it would seem, then, that seeds here can have fundamentally different constructions - largely radicle + hypocotyl, or largely cotyledon. In G. aristata, at least the seedling can start germinating before there is any root growth (Jaganathan et al. 2024). here may be polyembryony, and the parts of a Garcinia seed that has been cut up may develop into separate plants (see also Noor et al. 2016). Altogether a complicated story, and given the very different parts of the seed from which the radicle may appear, a study of embryo development would seem to be in order.
For general information, see Stevens (2006c), for chemistry, see Hegnauer (1966, 1989), Ribeiro et al. (2019) and Ferraz et al. (2020), for floral morphology of the distinctive Clusia gundlachii, see Gustafsson (2000), for that of other Clusiaceae, see Mourão and Beltrati (1995) and Mourão and Marzinek (2009), and for pollen, see Marinho et al. (2015).
Phylogeny. Clusieae are a well-supported clade clearly sister to the rest of the family (e.g. Ruhfel et al. 2016). However, the relationships of Symphonieae and Garcinieae are less clear, although they are provisionally separated here; there is no strong evidence for their reciprocal monophyly. For example, Gustafsson et al. (2002) had found that Garcineae were embedded in Symphonieae, Ruhfel et al. (2011) did recover a monophyletic Symphonieae but there was no support for a monophyletic Garcineae, rather, they were branches of a comb, while Ruhfel et al. (2016) recovered both as monophyletic.
- Within Clusieae, Dystovomita was found to be sister to the rest of the tribe and Tovomita is polyphyletic and the current infrageneric classification is not holding up (e.g. Ruhfel et al. 2016; Marinho et al. 2019, 2021: Dystovomita in basal pentatomy, new genus for some ca 18 species of Tovomita). There is good support for a monophyletic Clusia and many of the classical sections are turning out to be monophyletic, but relationships between these sections are less clear (Gustafsson et al. 2007). Luján (2019) looked at relationships between ca 86 species of Clusia, finding less resolution using four plastid markers than using nrITS; in the latter analysis, he recovered the sectional topology in one half of the tree of [Anandrogyne [Retinostemon + Cochlanthera]], while in the other half of the tree C. clusioides was sister to a clade of eight sections plus unassigned taxa, although deeper relationships here had little support; section Cordylanda - he included ten species of this section - was in this latter group. Interestingly, Luján et al. (2022: 64 species, RADseq) found that C. renggerioides, the only member of section Cordylandra they examined, was in the position of C. clusioides in Luján 2019 (that species was not included), while the bulk of the species examined were in a clade sister to this and again made up of the monophyletic and speciose sections Anandrogyne and Retinostemon (section Cochlanthera was not included); overall, relationships in this latter study had better support.
- Garcinieae. The relationship between Allanblackia, with its numerous ovules per carpel, and Garcinia s.l., with but a single ovule per carpel, have been unclear, and the former may be derived from the latter; an African clade made up of Allanblackia and some species of Garcinia has been recovered (Gustafsson et al. 2002; Sweeney 2008; Ruhfel et al. 2011, 2013, 2016; Gaudel et al. 2024). Gaudel et al. (2024: 111 spp., nuclear ribosomal and 3 chloroplast markers) provided a substantial amount of resolution to relationships within Garcinia s.l., the genus being divided into nine main clades and confirming a broad circumscription of the genus; the old Allanblackia was recognized as a section.
- Within Symphonieae, Symphonia is sister to the other genera (Ruhfel et al. 2016).
Classification. Clusia is to include genera like Renggeria, Decaphalangium, etc., previously segregated from it (Gustafsson et al. 2007). The old Tripetalum, Pentaphalangium and Allanblackia - and several other genera - are to be included in Garcinia (Sweeney 2008; Gaudel et al. 2024). Within Garcinia Gaudel et al. (2024) recognized 11 clades – all with (close to) maximal support - as sections, although a few species remained unplaced including the well-supported New Guinean species pair [G. archboldiana + G. engleriana].
[Calophyllaceae [Hypericaceae + Podostemaceae]]: exudate in mesophyll usu. in pale yellowish glands (unbranched canals).
Age. This node has been dated at (57-)54, 45(-42) Ma (Wikström et al. 2001), (73-)61, 45(-31) Ma (Bell et al. 2011), (88.7-)82.2(-73.6) Ma (Xi et al. 2012b: Table S7), ca 86.4 Ma (L. Cai et al. 2025b), around 91.3 Ma (Tank et al. 2015: table S2), or even older, (113.8-)106, 98.7(-87.4) Ma (Ruhfel et al. 2016: app. S9).
Evolution: Ecology & Physiology. Meseguer et al. (2017/2018) discuss the ecological evolution of the Calophyllaceae-Hypericaceae-Podostemaceae clade, suggesting that in the Late Cretaceous ca 100 Ma the clade grew in the Holarctic where conditions were relatively warm, if maybe somewhat drier than now. Conditions cooled after the Palaeocene-Eocene Thermal Maximum and members of the clade moved south following the warmer environments; Hypericum (q.v.), however, became adapted to cooler conditions.
CALOPHYLLACEAE J. Agardh Back to Malpighiales
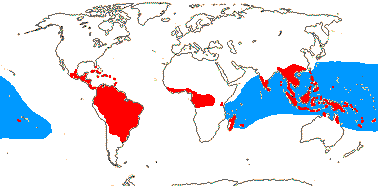
Trees or shrubs; (vessel elements with scalariform perforations); lamina vernation often flat, (paired "stipular" glands at the leaf base), (colleters 0); (plant dioecious); flowers usu. 4-5-merous; (K protective in bud), C (imbricate), (0-)4-5(-8); A not obviously fasciculate, (connate); style usually long, stigma wet; (exotegmen 0); embryo chlorophyllous or white, cotyledons relatively huge [cotyledon: hypocotyl + radicle ratio ≥5]; x = ?8 (?18, ?9).
13 [list: tribes]/460: two groups below. Throughout the tropics. Map: in part see Stevens (1980) and Trop. Afr. Fl. Pl. Ecol. Distr. 1. (2003) - blue is Calophyllum inophyllum.
Age. Crown-group Calophyllaceae are around (72.6-)57.6(-40) Ma (Xi et al. 2012b: Table S7), (93-)61.9, 56.4(-30.9) Ma (Ruhfel et al. 2016: app. S9) or (60.8-)56.9(-53.1) Ma (Cabral et al. 2021).
1. Endodesmieae Engler
Terminal bud 0; leaf with secondary veins closely parallel; G ?1, stigma punctate; ovule 1/carpel, apical; fruit indehiscent, seed single; n = ?; ?germination.
2/2. West Africa. Map: see above, continuous area of red in Central and West Africa, from Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003).
2. Calophylleae Choisy
(Terminal bud 0 - Mesua); leaves (spiral), (two-ranked), lamina vernation often flat (conduplicate; supervolute - Kiel.), secondary veins ± distant (closely parallel - Cal.), tertiary veins reticulate; anthers (locellate - Haploclathra), with complex or simple glands/0; pollen (in tetrads - Kiel.); G [2-5], stigmas much expanded to punctate; ovules (2-few/carpel), (basal), outer integument 20-30+ cells across, inner integument 2-3 cells across [Cal.]/integument single, ca 26 cells across [Mammea]; fruit (indehiscent); seeds 1-many/fruit, (variously winged); testa (multiplicative - Old World genera), vascularized [Mammea], (cotyledons relatively shorter); n = 14-16, 18, nuclear genome [2 C] 1.40±.02 pg; germination phanerocotylar, epigeal, or cryptocotylar, hypogeal.
11/458: Calophyllum (190), Kayea (70), Mammea (70), Kielmeyera (50). Throughout the tropics. Map: see above, blue is Calophyllum inophyllum, in part from Stevens (1980). [Photo - Flower, Flower.]
Age. The crown-group age of Calophylleae is estimated to be (42.9-)31.8, 29.5(-19.4) Ma (Ruhfel et al. 2016: app. S9) or rather younger, (29.4-)21.6(-15.1) Ma (Cabral et al. 2021).
Pollen identified as that of Kielmeyera (= Kielmeyerapollenites) has been reported from various places in India from the Palaeocene onwards (Kumar 1995: c.f. images of fossil and extant taxa). Ashton et al. (2021) suggest that the Kelmeyeropollis (sic) which Prasad et al. (2018a, b) mentioned is in fact Pentachlaena-type pollen (Sarcolaenaceae). Macrofossils, mostly leaf and wood, of Calophylluym are quite widespread and are known from deposits as early as the Palaeocene (Mahato et al. 2024).
Evolution: Divergence & Distribution. Fossils assignable to the Mammea americana group (embedded in an otherwise African clade in the genus - Cabral et al. 2021) are known from Panama in deposits ca 19 Ma (Nelson & Judd 2016). However, as with Clusiaceae, some fossils like Kielmeyerapollenites are both quite old and have very different distributions than those of the extant taxa with which they have been linked; K. eocenicus has, for example, been collected in deposits from Rajasthan in northwest India dated approximately to 50 Ma or a little more (Kumar et al. 2016), but crown-groupKielmeyera, from South America, is dated to ca 7.5 Ma (Cabral et al. 2021)... Other ages suggested by Nelson and Jud (2017) and Ziada (2023) are also quite old, some being Cretaceous.
Although Calophylleae are far more diverse than Endodesmieae, they still have a very long phylogenetic fuse (the age of stem Endodesmieae is not currently known) and it is thought that they have diverged only within the last 35 Ma or so (Ruhfel et al. 2016) or considerably less (Cabral et al. 2021). Calophylleae are probably Old World in origin, and Cabral et al. (2021: see below) recovered an Old World grade in which a New World clade was embedded; the few New World taxa in the former are embedded in the genera in which they occur. Diversification is notably increased in the Calophyllum and [Haploclathra [Kielmeyera + Caraipa]] clades; see Cabral et al. (2021) for more on diversification in the family. Kielmeyera is notably diverse in the Brazilian Cerrado, xylopodia, etc., having evolved, and there seems to have been hybridization there (Caddah et al. 2013).
The recent discovery of Calophyllum africanum, apparently related to the New World C. antillanum (anatomy, etc. of C. africanum are not reported, and its phylogenetic relationships are in fact unknown), in southwest Mali (and also Guinea - van der Burgt et al. 2022 for other examples of similar distributions, also Trichomanes) is biogeographically perplexing (Cheek & Luke 2016); apart from this species and the widespread C. inophyllum, which grows (just) on the east coast of Africa, there are no other species of the genus on the continent.
Pollination Biology. Buzz pollination occurs in Kielmeyera, while the distinctive cup-shaped anther glands more common on the related Caraipa are thought to secrete fragrances (Bittrich et al. 2006).
Vegetative Variation. Calophyllum longifolium (only the one species of the genus examined, but all species are similar) has unusually closely parallel secondary veins and transverse tertiaries (comparison with 484 other species of angiosperms: Sack et al. 2012); some Ochnaceae are similar (J. V. Schneider et al. 2016).
Recent work on Kielmeyera (K. appariciana - see Costa et al. 2021) has shown that there are two kinds of secretory canals in the plant, one in the secondary phloem and the other in the cortex and medulla, the latter being found in both stem and leaf. Although quite similar morphologically - both are schizogenous - resins rich in phenolics and terpenes that probably protect against herbivory are secreted in the cortical/medullary canals, while the canals in the phloem produce gums rich in polysaccharides that are perhaps involved in movement of water (Costa et al. 2021).
Genes & Genomes. There was a genome duplication in the ancestor of the [Calophyllum + Mammea] clade, and one in Mammea (Cai et al. 2017/18).
Chemistry, Morphology, etc.. For xanthones in Mesua, see Chukaew et al. 2019). Although all species of Calophyllum have opposite leaves when adult, a few species have seedlings with alternate leaves (Stevens 1980).
Marila asymmetralis, apparently alone in the clusioids, has obliquely monosymmetric flowers (pers. obs.). The androecial (and gynoecial) morphology of Endodesmia and Lebrunia needs study; is the former fasciculate (Ruhfel et al. 2013)? The glands on anthers of genera like Caraipa are large, paired and crateriform, perhaps because the contents have been removed, while in other genera like Kayea they are small and rounded. As in Clusiaceae, variation in ovule and seed morphology and anatomy is poorly understood; most data on ovule morphology come from Old World taxa.
For general information, see Stevens (2006c, as Clusiaceae), for chemistry, see Hegnauer (1966, 1989, as Guttiferae), for some anatomy, see Beauvisage (1920), for the distinctive foliar fibres of many species of Mammea, see Dunthorn (2009), and for fruits and seeds, see Mourão and Beltrati (2000).
Phylogeny. Endodesmia is sister to the rest of the family (Lebrunia has not been sequenced), which otherwise may separate into largely Old and New World clades, although these are not always well supported (Ruhfel et al. 2011, 2013, 2016), and low support is also true of many branches along the spine of the tree in Cabral et al. (2021). Cabral et al. (2021) found that the Old World taxa formed a grade within which the New World taxa formed a clade - the Old World taxa showed the relationships [Kayea [[Poeciloneuron + Mammea] [[Mesua + Calophyllum]... ]]]. Note that Kayea and Mesua, until quite recently considered to be congeneric, are placed on separate branches in the Old World clade (e.g. Zakaria et al. 2007; Ruhfel et al. 2016). Clusiella is in the New World clade (see also Gustaffson et al. 2002; Ruhfel et al. 2016), sister to Marila (Cabral et al. 2021). Its seeds and vegetative anatomy (including the deep-seated phellogen of the root) are consistent with this position, although the flowers are a little odd, since they do indeed look very like those of Clusia. Also in the New World clade, there is a group of largely alternate-leaved genera that form a clade (Ruhfel et al. 2013, 2016); these genera, [Haploclathra [Kielmeyera + Caraipa]], also have capsular fruits, often with quite large, winged seeds, and their embryos have large cotyledons with cordate bases. Relationships suggested by M. Sun et al. (2016) are somewhat different, while Trad et al. (2021: whole plastome analyses) found that the position of Mammea was problematic - it was either sister to all other Calophylleae examined or sister to a clade [Calophyllum + Mesua]; in different analyses that yielded the latter relationship, very few gene trees support it.
Previous Relationships. Many Theaceae also have spiral leaves, capsular fruits, winged seeds, and flowers with many stamens. Spiral-leaved Calophyllaceae seemed superficially to be similar and so used to be placed in Theaceae, being thought to be be more or less "intermediate" between Theaceae and Guttiferae - but the two families turn out not to be at all closely related (c.f. Baretta-Kuipers 1976).
[Hypericaceae + Podostemaceae]: stigma surface rounded, papillate; outer integument 2 cells across [?sampling]; plastome with ca 50 kb inversion in the large single copy region, accD sequences highly variable.
Age. Divergence between Podostemaceae and Hypericaceae may have occured as early as the Cenomanian, (106.5-)97.6, 90.9(-79.9) Ma (Ruhfel et al. 2016: app. S9), (82-)76, 72(-66) Ma (Davis et al. 2005a), ca 75 Ma (Tank et al. 2015: Table S2), (78.4-)69.7(-59.3) Ma (Xi et al. 2012b: Table S7), ca 76 Ma (L. Cai et al. 2025b) or as recently as (56-)43, 42(-26) Ma (Bell et al. 2010) or still more so, (42-)40, 36(-34) or (28-)26(-24) Ma (Wikström et al. 2001). The stem age of Podostemaceae, as sister to {Malpighiacedae [Cucurbitales + Rosales]], was estimated to be ca 106 Ma (T. Xue et al. 2020).
Evolution: Divergence & Distribution. The morphology of Podostemaceae is so highly derived that finding synapomorphies with Hypericaceae is difficult.
Genes & Genomes. Jin et al. (2020a) looked at the plastomes of Cratoxylum, a couple of podostems, and one species each in the other three families of the clusioid clade; the latter were unremarkable, but there were some similarities between the first two (see above) - something to follow up.
HYPERICACEAE Jussieu, nom. cons. - Back to Malpighiales
Small trees to shrubs; lignans, flavones, flavonols, (ellagic acid) +; stem cork pericyclic; polyderm widespread; petiole bundle arcuate (with wing bundle(s)); lamina vernation?; dark glands also present; (K protective in bud), quincuncial, C (imbricate); A (5-15), fasciculate + (not), development centrifugal, anthers dorsifixed, often with simple glands; styluli + (± connate), (stigma surface not rounded-papillate); ovules usu. many, (micropyle bistomal/zig-zag), inner integument 2-7 cells across; seeds (1-)many; (exotestal cells in vertical lines), (exotegmen 0); embryo chlorophyllous or white, cotyledons moderate in size [to 80% of the length of the embryo]; x = 9 (?10, ?7), nuclear genome [1C] (0.05-)0.398(-3.186) pg.
9 [list: tribes]/477 (590) - three groups below. World-wide (map: from Hultén & Fries 1986; Meusel et al. 1978; Trop. Afr. Fl. Pl. Ecol. Distr. 1. 2003). [Photo - Flower.]
Age. Meseguer et al. (2013: rooting?) suggested an age of (66-)53.8(-43) Ma for crown Hypericaceae, (63-)53.7(-42.5) Ma is the spread in Xi et al. (2012b: Table S7), (62.7-)52.3(-45) Ma in Nürk et al. (2015), or (93.3-)77, 71.5(-56) Ma (Ruhfel et al. 2016: app. S9).
For the fossil record of the family, both pollen and seeds, see Meseguer and Sanmartín (2012). Seeds of the fossil Hypericum antiquum from deposits in W. Siberia 40.4-33.9 Ma (see Budantsev 2005: p. 110, 111) are not particularly uniquely hypericaceous, although the exotestal cells in vertical rows are distinctive. Hypericum virginianum has rather different cell types with the walls of individual cells being largely separate, while the electron micrographs of the latter seem different yet again (Meseguer & Sanmartín 2012); perhaps c.f. also Elatine, some Nymphaea, Passifloraceae s.l., etc., for similar seeds. Paleoclusia chevalieri, based on fossils ca 90 Ma, may end up in crown-group Hypericaceae (Schönenberger et al. 2020; c.f. Crepet & Nixon 1998; Ruhfel et al. 2013), if so, this would lead one to question the dates just mentioned.
1. Vismieae Choisy
Shrubs to trees, (shooting from roots); C adaxially pubescent; A fascicles 5, staminodial fascicles 5; G [5]; fruit fleshy [berry or drupe]; embryo green (yellow); n = 10.
3/100. Vismia (55), Harungana (40). South America and Africa + Madagsacar.
Age. Estimates of the age of crown-group Vismieae are around (32.7-)19.6(-10.2) Ma (Nürk et al. 2015) or (60.9-)44.3, 40.7(-28.3) Ma (Ruhfel et al. 2016: app. S9) - barely overlapping.
Harunganoxylon vismioides was found in deposits of Eocene-Oligocene age on the Plateau de Tademaît, in the Sahara (Fessler-Vrolant & Starostin 1979).
[Cratoxyleae + Hypericeae]: fruit dry.
Age. This node is ca 40 Ma (Nürk et al. 2015).
2. Cratoxyleae Bentham & J. D. Hooker
Shrubs to trees; (C with adaxial basal nectariferous scale); A fascicles 3, staminodial fascicles 3; G [3], (with secondary septae); ovules (2≤/carpel); fruit loculicidal; seeds winged; n = 7, 9, 11.
2/7. Madagascar, tropical Southeast Asia-western Malesia.
Age. Crown-group Cratoxyleae are (41.1-)27.5(-11.2) (Nürk et al. 2015) or (70.4-)46.7, 43,7(-20.8) Ma (Ruhfel et al. 2016: app. S9).
3. Hypericeae Choisy - Hypericum L. —— Synonymy: Ascyraceae Plenck
Often herbs (annuals) to subshrubs (shrubs); vessel elements many [75-ca 500/mm2], short [<225 µm long, not Thornea] and narrow [<81 µm across]; (flowers 4-merous); A fascicles 4-5, variously organised, staminodial fascicles 0, 3 [(2) + (2) + 1], etc.; G [2-5], (placentation parietal); fruit (berry); seeds (winged), (carunculate); endosperm with chalazal cyst [closely packed nuclei], embryo suspensor unicellular; n = (?6, 7-)8-9(-12), etc.; nuclear genome [1C] ca (147-)410(-766) Mb [?level]; plastid transmission biparental.
1/517. Especially northern hemisphere, in the tropics ± montane.
Age. Crown-group Hypericum is estimated to be (37-)34.9(-34) Ma (Meseguer et al. 2013), (33.3-)25.9(-19.6) Ma (Nürk et al. 2015), or (52.2-)39.7, 37.3(-26.1) Ma (Ruhfel et al. 2016: app. S9).
Evolution: Divergence & Distribution. Meseguer et al. (2013) and Nürk et al. (2014, 2015) provide divergence dates within Hypericum.
Meseguer et al. (2013, 2014b, 2017/2018; see also Nürk et al. 2015) discuss diversification of Hypericum, integrating past distributions and past ecological preferences with the present, and they suggest that the ancestor of Hypericaceae may have been African, while the stem lineage of Hypericum itself was in the holarctic region by 50-35 Ma, the particular time depending on the models used. Nürk et al. (2015; see also Meseguer et al. 2017/2018; Tietje et al. 2022) thought that diversification in Hypericum could be explained by a late Eocene niche shift as the stem clade became adapted to cooler conditions, thus only some 15 Ma later did diversification rates in the genus increase in part triggered by cooling temperatures ca 14 Ma and in part by dispersal (twice) into South America, orogenesis in various parts of its range also contributing to the rate shifts. Meseguer et al. (2017/2018) suggested that diversification increased ca 10 Ma, extinction decreasing. There are major Old and New World clades of extant Hypericum, although the African H. lalandii is well embedded in the latter - probably long distance dispersal; Africa is otherwise thought to be the home of ancestors of the genus (see Meseguer et al. 2013 for more details).
Diversification of the ca 70 species of Hypericum in the Andean páramo has been dated to (5.6-)3.8(-2.3) Ma and has been accompanied by extensive variation in flower size, habit, etc., the plants ranging from prostrate shrublets to shrubs up to 6 m tall - there have also been some reversals to the herbaceous habit (Nürk et al. 2014, 2019: (4.3-)3.1(-2.0) Ma are the dates here). Disparification (Simpsonian adaptive radiation) and diversification (species number increase) have both been rapid here, the former perhaps less evident and the latter despite an increase in generation time (Nürk et al. 2019). Hypericum has also diversified in alpine habitats elsewhere (Hughes & Atchison 2015 and references, see also below), interestingly, a North American clade close to the páramo clade shows comparable height variation (Nürk et al. 2019). See also Hawaiian lobelioids and Asteraceae-silverswords, Echium, Lupinus, etc., for similar diversifications on (sky) islands.
Ecology & Physiology. Like several other bat-dispersed plants in the New World, Vismia tends to be a member of early successional communities (Muscarella & Fleming 2008). It dominates succession on old pastures, forming dense and persistent stands partly because of its root suckers (Mesquita et al. 2001).
Meseguer et al. (2017/2018) suggest in their discussion of the ecological evolution of the Calophyllaceae-Hypericaceae-Podostemaceae clade (q.v.) that cooling towards the end of the Eocene/beginning Oligocene may have caused an ecological shift that led to the evolution of crown-group Hypericum. Unlike the rest of the group, Hypericum became adapted to more temperate conditions with lower temperatures and precipitation and moved into various environments in the temperate biome, including newly-uplifted tropical montane habitats (see above: Meseguer et al. 2017/2018).
Pollination Biology & Seed Dispersal. The chirality of the contorted corolla varies within an individual in Hypericum, but with little effect on pollination (Diller & Fenster 2014, 2016).
Fruits of New World Vismia are an important food for phyllostomid bats such as Carollia (Lobova et al. 2009); other genera favoured by phyllostomids include Cecropia, Ficus, Piper and Solanum (Fleming 1986). The bats are fast feeders, ingesting the fruits and voiding the seeds in their faeces. Resin and seeds are collected from Vismia fruits by Melipona bees in the Neotropics and used to construct nest entrances, etc. (Roubik 1988).
Plant-Animal Interactions. Production of hyperforins (acylphloroglucinols, present in pale glands) and hypericins (phototoxic anthraquinones, present in dark glands) increased in Hypericum perforatum when eaten by generalist herbivores, but not by specialists (Sirvent et al. 2003).
Plant-Bacterial/Fungal Associations. The naptha-dianthrone hypericin may be synthesized by an endophytic fungus close to Chaetomium (Kusari et al. 2008).
Genes & Genomes. There was a genome duplication somewhere along the Hypericum clade (Cai et al. 2017/18: no other Hypericaceae examined). For chromosome numbers in Hypericum, etc., see Robson and Adams (1968).
Chemistry, Morphology, etc.. Hypericin, common here, is a red-colored naphthodianthrone derivative of anthraquinone (Zobayed et al. 2006). Details of the system of canals and spherical translucent reddish (schizogenous) or black (solid) glands in the plant, and what these structures secrete, are poorly understood (Nürk et al. 2012 and references). For details of the secretory structures in Hypericum, see in particular Curtis and Lersten (1990), Sirvent et al. (2003), Zobayed et al. (2006) and Lotocka and Osinska (2010) and references. As Ciccarelli et al. (2001) noted, in H. perforatum there were three different kinds of secretory canals, and the their contents varied according to where on the plant they were. Looking at the leaves, there are dark and translucent spots, the latter being similar to the secretory canals, and the contents of these spots also varies (Crockett 2010).
For general information, see Stevens (2006c), for chemistry, see Hegnauer (1966, 1989, as Guttiferae) and Crockett (2012: Hypericum), for the anatomy of Cratoxylum and Eliaea see Baas (1970), for the wood anatomy of Hypericum s.l., see Gibson (1980), for androecial development, etc., see Robson (1974 and references: fascicle/fasciclode number), Leins and Erbar (1991, 2010), Leins (2000), Ronse de Craene and Smets (1991e: Harungana), Prenner et al. (2008a: Hypericum androecium) and Rudall (2010), for ovules, see Guignard (1893) and Nagaraja Rao (1957), for fruits and seeds of Vismia, see Mourão and Beltrati (2001), and for seeds of Hypericum, see Robson (1981) and Meseguer and Sanmartin (2014).
Phylogeny. The basic relationships within Hypericaceae are sometimes recovered as [Cratoxyleae [Vismieae + Hypericeae]] (e.g. Ruhfel et al. 2016), however, Nürk et al. (2015) found the relationships [Vismieae [Cratoxyleae + Hypericeae]].
Within Vismieae, Harungana and the African Vismia rufescens form a clade with the American Vismia examined, and the other African Vismieae studied (= Psorospermum) formed a sister clade (Ruhfel et al. 2011, 2013; Xi et al. 2012b). Hypericeae: Nürk and Blattner (2010) discussed relationships and evolution in Hypericum in an analysis of morphological characters; the groupings they found had little support. Ruhfel et al. (2011) found some support for an expanded Hypericum, including Thornea. In a much more extensive study of Hypericum, Nürk et al. (2012: ITS only; see also Pílepic et al. 2011) found that Thornea was sister to Hypericum, but support for that position was not strong. In the analysis of Meseguer et al. (2013: four genes, inc. ITS), Thornea was a member of a basal polytomy, Triadenum was clearly to be included in Hypericum (see also Ruhfel et al. 2011; Meseguer et al. 2014; Z.-D. Chen et al. 2016), and some of the sections that Nürk et al. (2012) had found to be monophyletic were here paraphyletic; Santomasia was not included in these analyses. More recently, Nürk et al. (2015) has recovered a largely similar set of relationships, but again, support for the basal branchings was not strong. For diversification of South American Hypericum, see Nürk et al. (2014).
Classification. Generic limits need attention, with those of Hypericum and Psorospermum in particular needing to be expanded (Ruhfel et al. 2009, esp. 2011; c.f. in part Stevens 2006c). For a sectional classification of Hypericum, see Robson (2012) and references; Robson (2016) adjusted this somewhat to take on board molecular findings on relationships, unthought of when he began his studies.
PODOSTEMACEAE Kunth, nom. cons. - Back to Malpighiales
Annual herbs (perennials) of fast-flowing water; polyisoprenylated benzophenones 0, quinones 0, flavonoids/anthocyanina in roots; plant ± thalloid, stem, root and leaf often not distinguishable, plant attached to substrate by haptera, basic construction sympodial; roots photosynthetic, flattened, adventitious roots producing shoots; shoots endogenous, [also at least sometimes flowers], branching extra-axillary; vascular cambium 0; cork?; vessels usu 0; secretory cells +; stomata 0, intercellular spaces usu. 0, epidermal cells with SiO2 bodies and chloroplasts; cuticle waxes 0, cuticle + epidermis thin; leaves when present spiral/opposite/2- or 3-ranked, base broad or not; P +, uniseriate; androecial ?arrangement, filaments often basally connate (connective prolonged); pollen microechinate, infratectum granular; when G = P, opposite, stigma linear; ovules (2-few/carpel), integuments develop simultaneously [?level], outer integument (-4) cells across, inner integument ca 2 cells across; embryo sac protruding through the micropyle [?level], monosporic, from the subchalazal spore, tetranucleate [Apinagia type], polar nuclei degenerate; nucellus syncytial [= nucellar plasmodium/pseudo-embryo sac]; capsule ribbed, about the same size as the ovary, pedicels elongating; seeds dust-like [±0.315 mm long; exotesta thick-walled, often mucilaginous, (exo- and) endotegmen ± lignified; no double fertilization [= endosperm 0], terminal cell of suspensor enlarged, coenocytic/with filiform haustoria, cotyledons large; n = 10, x = 10 (?9. ?20); plastome ycf1 and ycf2 genes 0; root developing from hypocotyl, plumule rudimentary.
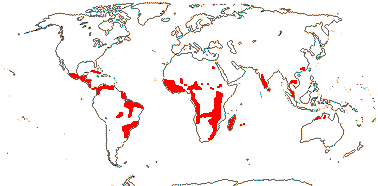
53 [list: to subfamilies]/340 - three subfamilies below. Usually tropical, esp. America.
Age. The age of crown-group Podostemaceae is around (89.4-)80.1, 74.5(-64.3) Ma (Ruhfel et al. 2016: app. S9).
1. Tristichoideae (Willis) Engler —— Synonymy: Philocrenaceae Bongard, Tristichaceae J. C. Willis
Xanthones?; primary roots producing shoots, (root 0), (root cap 0); primary shoot +, (main shoots from cotyledonary axils), shoots terete (dorsiventral), branches determinate, endogenous; stomata?; leaves linear (e.g. 2.67 x ca 0.05 cm) to linear-ovate, univeined; P 3, connate; A (1-)3, anthers sagittate; pollen pantoporate; G [3]; capsule 9-ribbed; hypocotyl 0.
5/28: Terniopsis (14), Dalzellia (9). India and Southeast Asia to Australia, Tristicha trifaria in Africa, Madagascar and the Mascarenes and Central America. Map: from van Royen (1953), Cusset and Cusset (1988a), Kito and Kato (2004) and Kato (2009).
Age. The age of crown-group Tristichoideae is ca (70.1-)56.9, 52.8(-38.6) Ma (Ruhfel et al. 2016: app. S9).

[Weddellinoideae + Podostemoideae]: radicle/primary root 0; G [2], 2-locular; hypocotyl +.
Age. The age of this clade is (76.8-)66.1, 62.0(-49.9) Ma (Ruhfel t eal. 2016: app. S9).
2. Weddellinoideae Engler - Weddellina squamulosa Tulasne
Plant with scales; root-born shoots 0; leaves ± linear, base sheathing; flowers single, terminal; P (4) 5 (6), 1-veined; A 5-25, anthers X-shaped; pollen ?development, lacking spinules, rugulate, ?infratectum?; ovary with apical septum having recurrent dorsal bundle, stigma single, globose; capsule not ribbed; tegmen [?layer] thick walled.
1/1. N. South America. Map: from van Royen (1953).
3. Podostemoideae (Warming) Engler —— Synonymy: Marathraceae Dumortier
Shoots scattered on root, (at root branching point), or lateral, apical meristem 0, (cryptic embryonic meristem), growth determinate; roots (exogenous), "adventitious", apical meristems on underside of the thallus, (crustose), (holdfasts 0), (rootcaps 0); thalli? foliose or ribbon-like; ("laticiferous" tubes +); "epidermal" cells with dimorphic chloroplasts; "leaves" ± endogenous [cell wall breakdown, cell separation], often 2-ranked, in tufts as short shoots/scattered along branches, ensiform, bifacial, (digitate, trilobed), (sHeath +), (dithecous [double-sheathed, one sheath on both sides]), (leaves with  axillary branches, not dithecous - Thelethylax), veins 0, (stipulate); flowers (sessile), monosymmetric (not), (inverted in bud - some African/Madagascan taxa); flower completely enveloped by a tubular non-vascularized spathella [?= connate T], (spathella of non-terminal flower open - Diamantina), (groups of flowers enveloped by spathella - Rhyncholacis clavigera); "P" linear [= staminodes], 2-25, often 2-3 on one side, (minute), A 1-3(-many), (A 2 - basally connate - Paracladopus), anthers often sagittate, (extrorse); (microsporogenesis successive [tetrads tetragonal]), pollen (in dyads, ?tetrads; (a)calymmate), 3(-5)-colpate; androgynophore +; G also [1, 3-7)], (ovary with apical septum having recurrent dorsal bundle), (unilocular), gynophore +/0, style short, branches long, (2 styluli - Diamantina/short, unbranched - Laosia/several branches - Zeylanidium); ovule with outer integument that develops first, (apex of nucellus exposed); (embryo sac bisporic [Polypleurum and Podostemon types]); plumule reduced, (0), (cotyledon 1); (hypocotylar root exogenous); (n = 14). Floral Diagram.
axillary branches, not dithecous - Thelethylax), veins 0, (stipulate); flowers (sessile), monosymmetric (not), (inverted in bud - some African/Madagascan taxa); flower completely enveloped by a tubular non-vascularized spathella [?= connate T], (spathella of non-terminal flower open - Diamantina), (groups of flowers enveloped by spathella - Rhyncholacis clavigera); "P" linear [= staminodes], 2-25, often 2-3 on one side, (minute), A 1-3(-many), (A 2 - basally connate - Paracladopus), anthers often sagittate, (extrorse); (microsporogenesis successive [tetrads tetragonal]), pollen (in dyads, ?tetrads; (a)calymmate), 3(-5)-colpate; androgynophore +; G also [1, 3-7)], (ovary with apical septum having recurrent dorsal bundle), (unilocular), gynophore +/0, style short, branches long, (2 styluli - Diamantina/short, unbranched - Laosia/several branches - Zeylanidium); ovule with outer integument that develops first, (apex of nucellus exposed); (embryo sac bisporic [Polypleurum and Podostemon types]); plumule reduced, (0), (cotyledon 1); (hypocotylar root exogenous); (n = 14). Floral Diagram.
47/294: Apinagia (50: ?paraphyletic, see Philbrick et al. 2001), Hydrobryum (17). Pantropical, perhaps esp. South America-Guianas. Map: from van Royen (1951), van Steenis (1972) and Kato (2009). Photo: Marathrum Flower.
Age. Crown-group Podostemoideae are some (63.9-)53.3, 49.7(-40.3) Ma (Ruhfel et al. 2016: app. S9).
Evolution: Divergence & Distribution. Trying to understand how the remarkable vegetative bodies of Podostemaceae have evolved is very difficult since it is unclear what you are looking at "is" morphologically; the major differences that are being found between taxa do not seem to have anything to do with ecology; and so on. Invoking saltational evolution and fuzzy morphology (e.g. Rutishauser 1983, 1995, 1997; Koi & Kato 2010; Koi et al. 2019; Kato 2025: esp. the latter) seems the best that can be done although from one point of view such concepts explain little; certainly, conventional morphology cannot cope with the family. Since Podostemaceae are sister to Hypericaceae and Calophyllaceae in turn, detailed studies of those families might provide clues about the evolution of the growth of the former, particularly building on the developmental studies of e.g. Katayama et al. (2010, 2013). However, there is little obvious in common between the vegetative construction of podostems and that of the other clusioids - perhaps there are similarities in the genomes?
Katayama et al. (2022) note two increases of molecular evolutionary rates here, one at the family level and the other at that of Podostemoideae, synonymous substitution rates increasing; they also note the accumulation of non-synonymous changes within Podostemoideae, perhaps because of relaxed functional constraints/selection pressures (for high evolutionary rates comapred with allied families, perhaps because of exposure to u.v. light, see Kato 2025). At the family level habitat changes, the adoption of the annual habit, and light-induced mutations, all interrelated, may be implicated in these changes, while in Podostemoideae the absence of a shoot apical meristem and its associated organizing/quiescent centre, i.e. there are no less frequently dividing cells at the apex, may interact with the annual habit and result in a further increase of mutation rates, and selection pressure seems to be reduced there (Katayama et al. 2022: e.g. Fig. 6). Kato et al. (2022b) place the evolution of the family in the context of the variation of 60 morphological characters; their work has been followed above.
Moline et al. (2007) discuss the phylogeny and evolution of African Podostemoideae, Koi and Kato (2010) that of Asian Podostemoideae, and Khanduri et al. (2015: also a morphological matrix) that of Indian members of the family.
However, it may be noted that "the most basic of taxonomic questions, ‘What species does this specimen represent?’ often remains difficult to answer, especially for members of Podostemoideae" (Ruhfel et al. 2024: p. 2).
Ecology & Physiology. Podostemaceae typically grow in fast-flowing rivers and in rapids, several species often being found together (they may be quite differently constructed), and they may be the only angiosperms in these places. Such plants are called rheophytes (van Steenis 1981, 1987; see also Kato et al. 2022b), and Podostemaceae are the major rheophytic clade. They are unusual among aquatic angiosperms in that they do not reproduce asexually and even the perennial species flower and fruit profusely (Philbrick & Novelo R. 1997).
For germination and establishment of the minute seeds, see e.g. Grubert (1970, 1976), Mohan Ram and Sehgal (1997), Leleeka Devi et al. (2016), and others. The testa swells and becomes mucilaginous, firmly attaching the seed to a rock, and although the radicle may not develop there are often dense "rhizoids" (= root hairs: Rutishauser 1997) at the base of any hypocotyl or radicle that is present, allowing the seedling to attach firmly. Haptera ("hapters") or holdfasts also develop, apparently from the stem, and attach the plant to the rock. Furthermore, although there have been suggestions (e.g. Rutishauser 1997) that Podostemaceae are attached to rocks by means of a special glue that they produce, it is more likely that it is materials in a biofilm produced by associated cyanobacteria that attach the plant to the substrate, hooked hairs on the root or stem of the plant sticking to the cyanobacterial filaments and associated biofilm (Jäger-Zürn & Grubert 2000). Indeed, these cyanobacteria may even produce nitrate that is used by the plant; Podostemaceae usually grow in oligotrophic rivers flowing over gneiss or granite, being absent in rivers over limestone (Jäger-Zürn & Grubert 2000). A common habitat for Podostemaceae is torrents in monsoon-savanna/rainforest areas, although rainforests are less favoured (Cook & Rutishauser 2006; Kato et al. 2022b).
Pollination Biology & Seed Dispersal. Wind pollination may be common in Podostemaceae, but the sometimes very large numbers of seeds per fruit produced would be distinctly unusual if that is the case (Philbrick & Novelo R. 1997). Self pollination seems to be pervasive in the family (Krishnan et al. 2019), the pollen tubes even growing through the tissue of the flower to the ovules (Sehgal et al. 2009).
Development of the fruit can be very rapid, taking a mere five days after fertilization (Krishnan et al. 2019). The seeds are minute, little larger than the ovules and often less than 320 µm long, although they can be around twice that size (Philbrick & Novelo R. 1997; Kato et al. 2022b). The seed coat is mucilaginous, aiding in the establishment of the seedling (Kato et al. 2022b).
Plant-Animal Interactions. Podostemaceae are major items in the diets of some South American serrasalmid myleine fish like Tometes that graze the plants on the rocks (Kolmann et al. 2020).
Plant-Bacterial/Fungal Associations. For the biofilm produced by a variety of cyanobacteria that attaches the plant to the rocks on which it grows, see above.
Vegetative Variation. In the following discussion, P = Podostemoideae, T = Tristichoideae and W = Weddellinoideae.
Interpretations of the construction of the plant body of Podostemaceae, sometimes called a thallus, vary greatly. The plant body can be thought of as a very highly modified but ultimately fairly conventional plant (Jäger-Zürn 1997b: p. 93, importance of "principles of variable proportions", 2005a), to a somewhat less enthusiatic embrace of classical morphological categories (Rutishauser 1997, 2016a), to thinking about the plant body as being sui generis (Koi & Kato 2010). See also Koi and Kato (2007) for hypotheses on the nature of shoots and leaves, Sehgal et al. (2007) for organ identity, and Rutishauser and Moline (2005: emphasis on "homology") and Kirchoff et al. (2008) for thinking about morphology in general - several examples in the latter are taken from podostems.
Seedlings of Dalzellia (T) lack a radicle, shoots develop from meristems in the cotyledonary axes, leaves of different sizes are born on the backs and sides of the shoots, and the meristematic region(s) at the apex show a complex developmental sequence of movements. The and reult is a plant body that is and apparently dichotomously branched thallus, the coenosome, made up of congenitally fused shoot axes (Imaichi et al. 2004).
The evolution of the remarkable flattened roots of some Podostemoideae and Tristichoideae, roots that lack root caps but have meristematic regions on both sides, from the more ordinary-looking roots found in Weddellinoideae and some other Tristichoideae has been carefully documented by Koi et al. (2006; see also Imaichi et al.2004). In Hydrobryum (P) foliose roots may revert to being ribbon-like (Koi & Kato 2020). The exogenous or superficial origin of secondary roots of some Podostemoideae is unusual, roots normally being endogenous and initiated inside the pericycle; Cladopus has both exogenous and endogenous lateral roots (Rutishauser & Pfeifer 2002), while in Zeylanidium roots may change into shoots. Some roots do have root caps.
In Tristichoideae the apex of the stem has a meristem with a tunica-corpus construction, but that of Podostemoideae nearly always lacks such a meristem (Koi et al. 2005). There is some controversy over whether normal axillary branching occurs or not (e.g. Rutishauser et al. 2005; Jäger-Zürn 2009a). Branch (= ramulus) growth in at least some Tristichoideae is determinate, and the ramulus complex is a sympodially-branched shoot system (Fujinami & Imaichi 2009). In Podostemoideae shoots may arise endogenously in the cortex (e.g. Moline et al. 2007). There is no conventional shoot apical meristem with its associated quiescent centre in Podostemoideae. There the leaf/bract is initiated as a shoot apical meristem and differentiates into a single apical leaf/bract, the result being the evolution of a shoot-leaf mixed organ (Katayama et al. 2010) - and with implications for the evolution of Podostemoideae (see above). In the dithecous leaves of Podostemoideae the leaf bases have two concave sheaths facing in opposite directions and with a flower or branch bud in the axils of each; these leaves usually terminate growth of the axis that bears them (Rutishauser et al. 2003); for the optimisation of these dithecous leaves on a phylogeny of Podostemoideae, see Moline et al. (2007: note the adaxial position of the prophyll of the axillary dithecal vegetative shoot illustrated). Jäger-Zürn (2009b) also depicts the dithecous leaf as being adaxial on the axillary shoot that bears it. Interestingly, the determinate ramulus system of Tristichoideae also involves the initial development of one or two cataphylls, the first (or only) cataphyll may be adaxial in position (Fujinami et al. 2013). There a distinction is made between cataphylls and scale leaves, although Terniopsis, sister to the rest of the subfamily, which is supposed to form a pair of leaves (?cataphylls) on all branches before the "ordinary scaly leaves" is not shown in their Fig. 6 as having such leaves. Fujinami and Imaichi (2015) put the evolution of flattened shoots in Tristichoideae in a phylogenetic context.
As mentioned, Katayama et al. (2010, see also 2013) found from gene expression patterns in two Podostemoideae that it was almost as if a determinate "leaf" capped the indeterminate stem, new leaves/branches developing endogenously from the base of the "leaf"; a local shoot apical meristem developed, and this was a mixed shoot/leaf structure (see also Xue et al. 2020). Since separation of the young "leaves" from the rest of the plant is by cell death, they will lack an epidermis, hence, perhaps, the absence of stomata in the subfamily - although they might be found on the flowers, in which there is in general more normal development in the family (e.g. Katayama et al. 2008). Katayama et al. (2019) followed the development of the zig-zag two-ranked sympodial shoot in the seedling of Zeylanidium tailichenoides, and they thought it represented a composite of two shoots that started as buds in the axils of the cotyledons; there was no terminal bud/plumule.
To summarize: The large and complex literature on podostem morphology needs a comprehensive and critical examination. Major reinterpretations continue, for instance, Jäger-Zürn et al. (2016) suggested that what had been called pinnate leaves in Castelnavia noveloi were in fact foliaceous roots.
Genes & Genomes. T. Xue et al. (2020) looked at the pattern of gene enrichment, etc., in the genome of Cladopus chinensis. The rate of change in the nuclear genome is highest in Podostemoideae and that of the plastome is highest in Tristichoideae (Katayama et al. 2022 and references).
Bedoya et al. (2019) discuss evolution in the plastomes of Podostemaceae; the plastomes are somewhat smaller than those of the few other members of the order with which they were compared, and Trad et al. (2021) found them to be somewhat smaller than other clusioids they examined, although not by that much (see also above). There have been a number of gene losses and pseudogenizations in the podostems. Thus the loss of the ycf1 and ycf2 genes there is uncommon in other angiosperms (Bedoya et al. 2019); they were also found to be absent in the podostems examined by Jin et al. (2020a), M. Zhang et al. (2022), M. Wu et al. (2022), etc.. The divergence of the accD sequences here seems to be associated with the loss of the ycf1 and -2 genes (Jin et al. 2020a). There is also an inversion of over 50 kb in the trnK-UUU-rbcL area, and this was not found in Cratoxylum (Hypericaceae: Trad et al. 2021, c.f. earlier reports).
Chemistry, Morphology, etc.. The "epidermal" cells of Podostemoideae have dimorphic chloroplasts; smaller chloroplasts are found against the outer periclinal walls and much larger normal-looking chloroplasts against the inner periclinal walls (Fujinami et al. 2011, 2015). Grubert (1976) noted distinctive contents in cells of young plants of several Podostemaceae, earlier Went (1926) had described "latex" (a white exudate) or "resin" that he found in sometimes elongated cells in the family. For SiO2 bodies, see Machado da Costa (2011) and in particular Machado da Costa et al. (2018); they are not found in all Podostemaceae.
The nature of the spathella has been unclear. Unlike bracts in Podostemoideae examined, its development is that of a "typical organ of leaf homology" (Katayama et al. 2010: p., see also Katayama et al. 2008), there is sometimes an apical meristem in the vegetative body of Podostemoideae and the spathella may be produced by the connation of two foliar structures (Jäger-Zürn 2005b). Eckardt and Baum (2010) suggest more specifically that it is calycine, however, there can be more than one flower per spathella so one would not have thought that it could be be simply calycine. Kato et al. (2022a: fig. 4 for floral diagrams) opt for an interpretation of the spathella as being perianthial. The "pedicel" is thus an androgynophore, while the linear "tepals" may be in the same whorl as the stamens, sometimes being lateral to/alternating with them, and they are interpreted as being staminodes. This interpretation is followed above, although there are still one or two loose ends. Some species of Dalzellia (Tristichoideae) have a cupule at the base of the pedicel that is formed by leafy shoot axes (Mathew et al. 2001).
As Koi and Kato (2012), Koi et al. (2019), and others describe the tepals, they seem more associated with the stamen(s) than the ovary, but they seem to be borne between the stamen and ovary in Paracladopus chiangmaiensis (M. Wu et al. 2022: Fig. 4). Tristicha - A 1, adaxial, median carpel abaxial? (Schnell 1998). The positions of the stamens may suggest that the flower has an oblique plane of symmetry (Cusset & Cusset 1988b), while Razi (1955; see also Endress & Matthews 2006a) described the flowers of Zeylanidium olivaceum (Podostemoideae), which have a spathella, two stamens and two carpels, as being monosymmetric.
The outer integument develops early and the embryo sac plus nucellar epidermis comes to protrude beyond it, even if the ovule is described as having an exotegmic micropyle (e.g. Arekal & Nagendran 1977). There is considerable variation in the development of the embryo sac in particular (and a corresponding amount of controversy), perhaps especially in Weddellina (see e.g. Mukkada 1962; Battaglia 1971; Arekal & Nagendran 1975, 1977a, b; Nagendran et al. 1976; Murguía-Sánchez et al. 2002; Sehgal et al. 2011a, b). Is it mono- or bisporic?; can the synergids be chalazal?; does it have 3, 4 or 5 cells at maturity? (note: never eight, there are no antipodals). However, there is a general consensus that there is no double fertilization, and the central cell degenerates, or never really differentiates in the first place (Krishnan et al. 2019). During development of the embryo, the nucellus immediately beneath it becomes plasmodial/syncytial, the walls breaking down (different pathways are involved), the so-called pseudo-embryo sac or nucellar plasmodium, and this helps in the nutrition of the embryo (Jäger-Zürn 1997b; see also Plachno & Swiatek 2010b). Kato et al. (2022b: Fig. 1) suggest that this nucellar plasmodium develops before (Tristichoideae), just before (Weddellinoideae), or after (Podostemoideae) fertilization.
Tobe and Raven (2011) described the tegmen as being unspecialised. Although the embryo usually lacks a plumule and radicle, these have been reported in Malaccotristicha sp. (Kita & Kato 2005). Seedlings of Podostemoideae have a hypocotyl, if short, and short hypocotyls have also been reported in Tristichoideae (e.g. Mohan Ram & Sehgal 1997). Koi et al. (2012b) discuss seedling evolution in the family, and see also Suzuki et al. (2002) for seedlings. More information is needed on embryo morphology (but see Koi & Kato 2010).
For general information, see classics such as Willis (1902a, b), also Graham and Wood (1975), Grubert (1974b), Aquatic Bot. vol. 57 (1997), Ameka et al. (2002) and Kato (2004, 2006, 2008, 2013: ?coffee table book on podostems), while much information is taken from Rutishauser (1997), also Cook and Rutishauser (2006); see also Hegnauer (1969, 1990), Contreras et al. (1993) and Kato et al. (2005), all chemistry, Barlow (1986), Hiyama et al. (2002) and Koi and Kato (2003), all roots, Jäger-Zürn et al. (2006: microsporogenesis), O'Neill et al. (1997), Lobreau-Callen et al. (1998) and Passarelli (2002), pollen, Jäger-Zürn (2003: apical septum, 2007: shoot apex; 2011: possible new characters), Sehgal et al. (2002: seeds, etc.), Cusset and Cusset (1988a, b) and Rutishauser and Huber (1991), Tristichoideae, Rutishauser et al. (2004: Diamantina), Rutishauser and Grubert (1993: Mourera, 2000: Apinagia), Grob et al. (2007b) and Jäger-Zürn (2008), both Thelethylax, Thiv et al. (2009: African Podostemoideae), Ghogue et al. (2009: Djinga, morphology), Koi and Kato (2010: vegetative body, Hydrodiscus et al.) and de Sá-Haiad et al. (2010: Podostemon: floral morphology).
Phylogeny. The basic phylogenetic structure of the family is [Tristichoideae [Weddellinoideae + Podostemoideae], all clades having very strong support (Kita & Kato 2001; see also Kita 2002 and notably: phylogeny and morphology; esp. Koi et al. 2012a; Ruhfel et al. 2016; etc.).
In New World Podostemoideae morphological analyses allowed the recovery of a few small generic clades, but molecular data resolved quite a number of nodes (Tippery et al. 2011). Diamantina appeared to be sister to all other Podostemoideae (Ruhfel et al. 2009), although that genus was not studied by Tippery et al. (2011), who found Podostemon and a paraphyletic Mourera as successively sister to the rest of the subfamily, while Diamantina was sister to the other members of one of the two major clades into which Podostemoideae was resolved in Ruhfel et al. (2016) - Mourera belonged to that clade, Podostemon to the other. Koi et al. (2012a: very good sampling) confirmed the position of Diamantina, although support was weak; further large scale structure in Podostemoideae mostly had very little support, and generic monophyly seems to be almost a foreign concept there. See also M. Sun et al. (2016) for a phylogeny, Werukamkul et al. (2018) for relationships among Thai taxa in particular, and Koi et al. (2019) for a matK analysis of Asian Podostemoideae (Zeylanidium poly/paraphyletic). Ruhfel et al. (2024: 68 spp., ca 1/4 the family, 73 protein-coding plastome genes, focus on New World Podostemoideae) found the relationships [Devillea [[Clade A - African-Madagascar and S.E. Asia-Australia taxa derived] [Clade B, all New World]]], Apinagia, Marathrum, Oserya and Podostemum were all polyphyletic.
Koi et al. (2009) discussed the phylogeny of Tristichoideae (and described a distinctive new genus), while Koi et al. (2012a) found groupings in Tristichoideae to be well supported and the genera to be monophyletic (see also Ruhfel et al. 2016: Terniopsis sister to the other genera). Koi et al. (2022) found some differences between the ITS and matK trees in the topologies they recovered. Thus in the former Cussetia was embedded in Terniopsis, while in the latter the two were sister taxa; generic limits followed the latter topology, but this is another case where analysis of nuclear genomes may be interesting.
Classification. Many genera (22/53, 41% - Katayama et al. 2022) are monotypic, the morphology of the thallus being so bizarre. For a synopsis of Podostemoideae, see Koi et al. (2022), and for generic limits in some African Podostemoideae, see Thiv et al. (2009).
Previous Relationships. Prior to molecular work, systematists were largely at a loss as to where the relationships of Podostemaceae were to be found. The micropylar suspensor haustorium seemed like that of Crassulaceae, perhaps suggesting a link between the two families (e.g. Les & Philbrick 1996; Ueda et al. 1997a), or isolated in its own order in Rosidae (Cronquist 1981), etc.. Indeed, Podostemaceae have sometimes been placed apart from all other angiosperms (e.g. Cusset & Cusset 1988b), while as mentioned elsewhere nuclear genome data may rather change things (see Baker et al. 2021: esp Seed Plant Tree of Life).
Possible clade.
[[[Lophopyxidaceae + Putranjivaceae], Caryocaraceae, [Centroplacaceae [Elatinaceae + Malpighiaceae]], [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]]] [[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]]] [ Euphorbiaceae [[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]]]: ovules 2/carpel, apical, pendulous, epitropous.
Evolution: Divergence & Distribution. Although ovule number can be optimised to this node, both the position and orientation of the two ovules varies...
[[Lophopyxidaceae + Putranjivaceae], Caryocaraceae, [Centroplacaceae [Elatinaceae + Malpighiaceae]], [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]]] / Clade 3 of Xi et al. (2012b): G tends to have longitudinal bulges above the placentae; outer integument 3-7 cells across, inner integument 5-10 cells across [inner integument often thicker than outer].
Age. The spread of ages for this clade was estimated to be (111.4-)107, 102.3(-95.8) Ma (Xi et al. 2012b: Table S7).
Evolution: Divergence & Distribution. There are three cases of the evolution of monosymmetric flowers in which the odd=banner petal is adaxial, i.e. in Malpighiaceae, Chrysobalanaceae and Trigoniaceae (Bukhari et al. 2017).
Phylogeny. For discussion on relationships here, see below.
[Lophopyxidaceae + Putranjivaceae] / Putranjivoids: mucilage cells 0; hairs unicellular; pedicels at most barely articulated; flowers imperfect; K not protective in bud; staminate flowers: A in one whorl, anthers ± basifixed, pistillode +; carpellate flowers: staminodes 0; style ± 0; ovules with inner integument much thicker than the outer [5≤ cells across], endothelium +, parietal tissue +, but only weakly crassinucellate, funicular-placental obturator +; fruit indehiscent, 1-seeded; plastome inverted repeat [IRb] 0.
Age. Lophopyxidaceae and Putranjivaceae may have diverged in the Cretaceous at end-Coniacian or thereabouts ca 85 Ma (Davis et al. 2005a) or (35.8-)34.2(-33.1) Ma (Xi et al. 2012b: Table S7).
Evolution: Divergence & Distribution. Jin et al. (2020b: Fig. 2) noted that gene and intron losses both characterized the group as a whole and also individual species/species groups/?families within it; these have not been added to the hierarchy above.
Genes & Genomes. Jin et al. (2020b) sequenced some plastomes from this family pair, and found that the chloroplast inverted repeat (IRb) was missing. Associated with this loss, there were gene and intron losses, large inversions, and translocations and duplications within the plastome (Jin et al. 2020b). The plastome was some 40,000 bp shorter that that of Balanops, which had a normal plastome.
Chemistry, Morphology, etc.. For detailed studies of the floral morphology of these two families, see Matthews and Endress (2013); much information is taken from this account.
Phylogeny. Davis et al. (2005a) found a strong association between this family pair, and they may in turn be associated with the group of families with parietal placentation. However, the position above has strong support in Xi et al. (2012b).
LOPHOPYXIDACEAE H. Pfeiffer - Lophopyxis maingayi J. D. Hooker - Back to Malpighiales
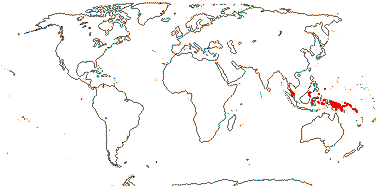
Lianas, climbing by leaf tendrils; chemistry?; secondary thickening anomalous, with included phloem; vessel elements with simple perforation plates; phloem stratified; pith pentagonal; nodes ?; stomata paracytic;; petiole bundles arcuate and with wing bundles; branches with prophyllar bud at base; leaves spiral; plant monoecious; inflorescence branched, ultimate units clusters; pedicels slender; flowers small, [<4 mm across]; K connate basally, valvate, C shorter than K; staminate flowers: nectary glands cordate or reniform, adnate to C; A = and opposite K; carpelate flowers: nectary glands basally connate; G [(4) 5], opposite C, ovary ridged, stigmas spreading, subulate, adaxially channelled; ovules with outer integument 3-5 cells across, inner integument 5-10 cells across, suprachalazal zone long; fruit a 5-winged samara, K persistent; seed single, coat?; endosperm ?development, +, cotyledons long; n = ?
1/1 [list]. Malesia to the Solomon and Caroline Islands. Map: from Sleumer (1971b).
Evolution: Divergence & Distribution. The diversification rate in Lophopyxidaceae seems to have slowed down (Xi et al. 2012b).
Chemistry, Morphology, etc.. Sleumer (1971b) described the tendrils both as leaves and bud-bearing branches; the ultimate spirally-recurved portion does seem to be foliar.
Utteridge (2019) described the fruit as being a capsule.
For general information, see Kubitzki (2013b), for ovule morphology, see Mauritzon in Sleumer (1942).
Lophopyxis is poorly known.
Previous Relationships. Lophopyxidaceae were included in Celastraceae by Cronquist (1981) and Hutchinson (1973), and in Celastrales by Takhtajan (1997).
PUTRANJIVACEAE Meisner - Back to Malpighiales
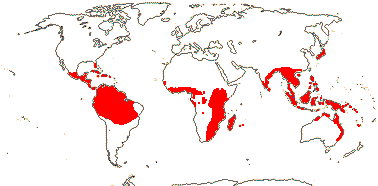
Trees; cucurbitacins [triterpenes], glucosinolates, biflavonoyls +; cork?; vessel elements with scalariform perforation plates; stomata various; petiole bundles elliptic; leaves two-ranked, (lamina with veins running into opaque deciduous teeth, or spines); plant dioecious; inflorescence fasciculate; (K with single trace), C 0, 4-5(-7); nectary + or 0; staminate flowers: A (2-)3-20(-many), (extrorse), (with pseudopit); carpellode?; carpellate flowers: (staminodes 0); G [(1-)2-4(-9)], (style very short, branched), stigmas large, often flap-like, bifid, ?type; ovules (micropyle exostomal), outer integument 3-9 cells across, inner integument 6-14 cells across, (integument single, 6-9 cells across - Drypetes macrostigma), parietal tissue 1-2 cells across, disintegrating early, suprachalazal zone long to short, hypostase massive; megaspore mother cells 2-3; fruit a drupe; testa vascularized, exomesotesta sclereidal, tegmen ± multiplicative, 6-24 or more cells across, exotegmic cells sclereidal, flat-lying; endosperm copious; n = (19) 20 (21).
3 [list]/210: Drypetes (200). Tropical, esp. Africa and Malesia (map: from FloraBase 2005; Trop. Afr. Fl. Pl. Ecol. Distr. 2. 2006; Andrew Ford, pers. comm.). [Photo - Flower, Fruit]
Evolution: Divergence & Distribution. Some analyses suggest that the diversification rate in Putranjivaceae increased (Xi et al. 2012b).
Pollination Biology. Glucosinolates make the flowers of Drypetes smell very strongly; bees, wasps and beetles have been recorded visiting D. natalensis where constitutively-released isothiocyanates are part of the floral odour, although what exactly they do is unclear (S. D. Johnson et al. 2009a, c).
Plant-Animal Interactions. Perhaps not surprisingly, caterpillars of pierid butterflies have quite often (23/2690 records) been recorded eating Putranjivaceae (see also Brassicales and Fabaceae). Species of the Indo-Malesian Appias subgenus Catophaga (albatrosses) feed more or less indiscriminately on Drypetes (Putranjivaceae) and Capparaceae (Yata et al. 2010).
Genes & Genomes. Drypetes (Euphorbiaceae sister) has a genome duplication (Cai et al. 2017/18).
Chemistry, Morphology, etc.. Older literature is under Euphorbiaceae. For general information, see Levin (2013), for chemistry, see Hegnauer (1966, 1989), for glucosinolates, see Montaut et al. (2016), for wood anatomy, Hayden and Brandt (1984: like that of Aporosa, etc. [= Phyllanthaceae]), for epidermal features, see Kadiri and Muellner-Riehl (2021), for pollen, Köhler (1965), and for embryology and seed anatomy, see Singh (1970) and Stuppy (1996).
Phylogeny. Tokuoka and Tobe (1999, 2001) included Lingelsheimia here, but it has a tegmen only 3-4 cells thick and a vascularized testa and it is to be placed in Phyllanthaceae (see Kathriarachchi et al. 2005).
Classification. For a checklist and bibliography, see Govaerts et al. (2000).
Previous Relationships. Putranjivaceae were usually included in Euphorbiaceae (as by Webster 1994b, in Phyllanthoideae), but can be distinguished i.a. by their chemistry, embryology and fruit. They are certainly not to be placed with the rest of the glucosinolate families in Brassicales (e.g. Rodman et al. 1997, 1998).
CARYOCARACEAE Voigt, nom. cons. - Back to Malpighiales
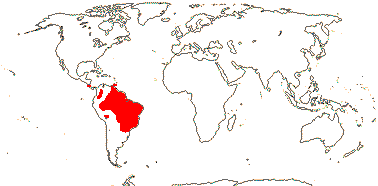
Trees to shrubs; ellagic acid; vessel elements with simple (scalariform) perforations; nodes 5 or more:5 or more; petiole bundles incurved arcuate, irregularly annular, etc.; pericyclic sheath little lignified; branched sclereids +; cuticle waxes smooth to irregular platelets; colleters +; leaves trifoliolate, leaflets ± articulated, (margins entire), stipellate or not; inflorescences terminal, racemose-corymbose; pedicels articulated, bracts 0; flowers large [>5 cm across], (6-merous); K imbricate, C protective in bud, with 3 traces, connate below, or forming a deciduous calyptra; A many, connate at base and adnate to C, (in 5 bundles), filaments long, tuberculate/vesiculate esp. towards apex [non-volatile oils secreted], anthers basifixed, inner stamens staminodial; placentation basal, styluli +, impressed, arising together, with 2 vascular bundles, stigmas punctate-impressed; ovule 1/carpel, basal, erect, campylotropous to anatropous, sessile [attachment broad], micropyle endostomal, outer integument 2-3 cells across, inner integument 3-4 cells across, parietal tissue?, epidermal cells of nucellar apex radially elongated, nucellus below embryo sac massive, obturator 0; fruit a drupe, stone separating into 1-seeded units; seeds reniform, hilum large; coat undistinguished, testa vascularized, aerenchymatous or not, exotegmen?; endosperm ?type, at most thin; n = 23.
2 [list]/27. Tropical America, esp. Amazonia. Map: from Prance and Freitas da Silva (1973). [Photo - Flower, Flower, Fruit.]
Age. Crown-group Caryocaraceae are (57.2-)55.8(-54.8) Ma (Xi et al. 2012b: Table S7).
Triterpenoid saponins +; leaves opposite, stipules inter-intrapetiolar; nectary at base of staminodia; G [4-6], pericarp with radiating fibres.
1/18. Tropical America.
Stipules ± intrapetiolar; K small, ± connate, lobed; nectary at base of staminodia 0; G [8-20]; ovules unitegmic, integument 4-5 cells across; hypocotyl very large, oily, spirally-twisted.
1/8. Tropical America.
Evolution: Pollination. Both genera are pollinated by bats (Fleming et al. 2009).
Plant-Animal Interactions. The diversity of ant species visiting the extrafloral nectaries of Caryocar, especially of C. brasiliense, is particularly noteworthy (Novais et al. 2025).
Genes & Genomes. The plastome of Caryocar brasiliense was quite like that of most other Malpighiales, differences like having a somewhat larger inverted repaet, its boundary with the SSC region being flanked by ndhH and a ndhH pseudogene, not ycf1, being relatively minor (Nunes et al. 2020b).
Chemistry, Morphology, etc.. Prance and Freitas da Silva (1973) described Anthodiscus as lacking stipules.
Dickison (1990c) described details of the complex floral vasculature and other floral features; in Anthodiscus each stylulus receives a vascular bundle from adjacent carpels and so is presumably commissural, while the styluli of Caryocar are vascularized from single carpels. The androecium may form a ring primordium (Ronse de Craene & Smets 1992b).
See Hegnauer (1964, 1989, the latter also under Lecythidaceae) and Chisté and Mercadante (2012) for chemistry; the fruits of both genera are used as fish poisons.
For general information, see Prance (2013) and Nunes et al. (2020a), for vegetative anatomy, see Beauvisage (1920) and for leaf teeth and colleters, see Rios et al. (2020).
All in all, the family is rather poorly known, especially embryologically.
Previous Relationships. Both Cronquist (1981) and Takhtajan (1997) included Caryocaraceae in Theales.
[Centroplacaceae [Elatinaceae + Malpighiaceae]] / Malpighioids: pedicels articulated; K persistent in fruit.Age. The age of this clade is (109.4-)102.1(-93.3) Ma (Xi et al. 2012b: Table S7) or ca 90.3 Ma (Tank et al. 2015: table S2, younger than next node up); Cai et al. (2016) suggested that it may be ca 105 or (110.1-)109(-106.1) Ma.
Phylogeny. For relationships in this area, see W. Zhang et al. (2009a, 2009b, 2010) and Wurdack and Davis (2009). Xi et al. (2012b) found only weak support for the inclusion of Centroplacaceae in this clade (63% ML bootstrap; 0.51 PP).
CENTROPLACACEAE Doweld & Reveal - Back to Malpighiales
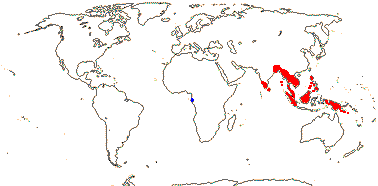
Trees; inflorescence branched; A 5, opposite K; style branches widely diverging, stigmas little expanded; ovules collateral; fruit a loculicidal capsule, seed one/loculus, with [?]exostomal aril; exotegmic cells laterally compressed, thick-walled; embryo short.
2 [list]/6. West Africa, Indo-Malesia.
Age. The crown age of this clade is around (94.3-)63.7(-35.1) Ma (Xi et al. 2012b: Table S7) or (87.7-)69(-78.5) Ma (Cai et al. 2016).
1. Centroplacus glaucinus Pierre
Chemistry?; cork?; vessel member perforations?; sclereids +; stomata anisocytic; leaves two-ranked, stipules cauline; plant dioecious; flowers small; nectary outside A, lobed, lobes alternating with K; staminate flowers: anther dehiscence oblique-apical, connective well developed; pollen psilate, perforate; pistillode +; carpelate flowers: C 0; staminodes +, minute; G [3], placentae swollen; ovules subapical, ?morphology, obturator 0; fruit also septicidal, opening from the base; exotestal cells rather tall, outer wall thickened, mesotegmic cells flattened, at right angles to exotegmen, endotegmen ± thick-walled; n = ?
1/1. W. Africa. Blue on map above: from Trop. Afr. Fl. Pl. Ecol. Distr. vol. 2 (2006).
2. Bhesa Arnott
Chemistry?; (cork mid-cortical); vessel elements with scalariform perforation plates; paratracheal parenchyma +; nodes 5:5; calcium oxalate as crystals [?always]; petiole with bundles forming a U or flattened-annular, 2-3 medullary bundles, (also wing bundles); stomata laterocylic; leaves spiral, lamina vernation conduplicate, margins entire, tertiary venation closely scalariform, petiole ± pulvinate apically, stipules almost encircling the stem, colleters +; inflorescence racemose; C contorted; A extrorse to introrse; pollen finely striate; nectary lobed or not; G [2]; ovules basal, apotropous, micropyle exostomal, outer integument 6-8 cells across, inner integument 4-5 cells across, hypostase +; fruit loculicidal; aril sheet-like; exotegmic cells massive, exotegmic cells laterally compressed tracheidal; n = ?; germination epigeal.
1/5. Indo-Malesia. Red on map above: from Ding Hou (1962).
Evolution: Divergence & Distribution. An odd couple. Xi et al. (2012b: some analyses) found that diversification rate in this clade slowed down...
Plant-Bacterial/Fungal Associations. Bhesa is reported to be ectomycorrhizal (Smits 1994).
Genes & Genomes. Bhesa may have a genome duplication (Cai et al. 2017/18).
Chemistry, Morphology, etc.. The ribbon-shaped exotegmic cells of Bhesa are longer than those of Centroplacus, and its integument is much thicker. The endostome of Centroplacus is lignified and more or less protruding.
For general information, see Kubitzki (2013b): Bhesa has often been included in Celastraceae, see Pierre (1894), Ding Hou (1962) and Wurdack and Davis (2009), general information, for seed and vegetative anatomy of B. ceylanica, see Jayasuriya & Balasubramaniam 3107, for seeds of B. robusta, see Corner (1976). Centroplacus has often been included in Euphorbiaceae: see Forman (1966) and Radcliffe-Smith (2001), both general, Stuppy (1996: seed anatomy and good discussion), and Tokuoka and Tobe (2001: seed anatomy).
Studies of the embryology, etc., of both genera are much needed.
Phylogeny. Cai et al. (2016) show Centroplacus rather surprisingly embedded in a paraphyletic Bhesa; no comment was made, but this might perhaps be a rooting artefact.
Classification. Recognising the two genera as a single family seems most reasonable, although they are very different in their gross morphology.
Previous Relationships. Centroplacus glaucinus has been placed in Pandaceae (Takhtajan 1997; Mabberley 1997), but Webster (1994b) and Radcliffe-Smith (2001) included it in Euphorbiaceae, but only with hesitation and with little certainty as to where it should be placed within the family.
Bhesa was distinctive in morphological analyses of Celastraceae, in which it used to be included (Simmons & Hedin 1999; Matthews & Endress 2005b), if sometimes with some doubt (e.g. Pierre 1894 [he thought it might be in a separate family]; Metcalfe & Chalk 1950; Ding Hou 1962). Its huge stipules, distinct styles, vessels with scalariform perforation plates, etc., were somewhat out of place there, although Celastraceae were so heterogeneous that a strong case could not be made for its removal. The seed coat, with its massive exotegmic cells, is also very different, as are its pentalacunar nodes.
[Elatinaceae + Malpighiaceae]: vessel elements with simple perforation plates; sieve tube plastids lacking starch and protein inclusions; leaves opposite, with glands[?], (lamina margins entire); inflorescence cymose; flowers with inverted orientation [odd petal adaxial]; connective glandular; nectary 0; when G 3 median member adaxial; hypostase +; fruit septifragal; endosperm slight; x = 6.
Age. Malpighiaceae and Elatinaceae may have separated some (113-)98, 89(-85) Ma (Davis et al. 2005a), (110-)107, 98(-94.5) Ma (Cai et al. 2016), ca 93.7 Ma (Tank et al. 2015: table S2), (99.9-)86.1(-72.9) Ma (Xi et al. 2012b: Table S7) or ca 97.2 Ma (L. Cai et al. 2025b).
Evolution: Divergence & Distribution. Cai et al. (2016) suggested that the American/African disjunction between stem-group Malpighiaceae and Elatinaceae reflected Gondwanan vicariance.
Depending on the interpretation of variation, both the evolution of monosymmetric flowers and flowers with an inverted orientation can be pegged to this node (W. Zhang et al. 2009a, 2010: see also the discussion after Malpighiaceae).
Chemistry, Morphology, etc.. For the foliar glands and resin and latex production in Elatinaceae and Malpighiaceae, neither well understood, see Vega et al. (2002) and Davis and Chase (2004). Vega et al. (2002) suggested that the laticifers of Galphimieae might be a symplesiomorphy with those of Euphorbiaceae, however, the phylogenetic status of laticifers in Euphorbiaceae is unclear, and that family is not immediately related to Elatinaceae-Malpighiaceae; I know nothing of the composition of the latex of Malpighiaceae. Bonifácio et al. (2023) discuss glands on the sepals and connective in both Malpighiaceae and Elatinaceae; what, if anything, is secreted in the latter family is unclear
For variation in seed size, see Moles et al. (2005a).
Phylogeny. Support is strong for the sister-group relationship between Malpighiaceae and Elatinaceae (e.g. Davis & Chase 2004; Tokuoka & Tobe 2006; Korotkova et al. 2009; Wurdack & Davis 2009; Wang et al. 2009; Xi et al. 2012b, etc.).
ELATINACEAE Dumortier, nom. cons. - Back to Malpighiales —— Synonymy: Cryptaceae Rafinesque
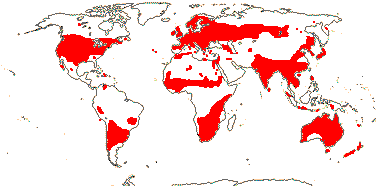
Herbs to subshrubs (annuals) of moist/wet habitats; flavonols, ellagic acid +; plant resinous; (cork from inner cortex); stem with endodermis [Elatine]; nodes 1:1; mucilage cells +; (plant glabrous), leaves with colleters; lamina with marginal hydathodes/glandular hairs, stipules scarious; ?pedicel articulation; flowers (single), 2-5(-6)-merous, K connate (free), (with an apical "gland"), vascular trace 1, C contorted or imbricate; A (1), = and opposite K, or 2x K, (bisporangiate), endothecial thickening 0; tapetal cells binucleate; (pollen grains tricellular - E.); G [2-5], opposite K, stigma papillate; ovules many/carpel, (not vascularized), micropyle bistomal, zigzag, outer integument 2-3(-5) cells across, inner integument 2-3 cells across, parietal tissue 1-4 cells across, suprachalazal zone at least initially long [Bergia]; megaspore mother cells several; fruit a capsule; seeds ± curved; exotestal cells in longitudinal series, exotegmen with low lignified sinuous anticlinal cell walls; embryo ± fusiform; n = (9, 18, 20), x = 10 (?14, ?5); duplication of CYC genes.
2 [list]/35: Bergia (25), Elatine (10). Worldwide, but more tropical, not Arctic. Map: from Meusel et al. (1978), Frankenberg and Klaus (1980), FloraBase (consulted 2006) and Popiela et al. (2012).
Age. The age of crown-group Elatinaceae is thought to be (76.3-)48(-25.1) Ma (Xi et al. 2012b: Table S7), (96.8-)85, 82(-73.6) Ma (Cai et al. 2016) or ca 59.6 Ma (L. Cai et al. 2025b).
Seeds (they are straight) identified as cf. Elatine have been found in deposits 36.0-32.5 Ma from Peruvian Amazonia (Antoine et al. 2021).
Evolution: Divergence & Distribution. See Cai et al. (2016) for a discussion on the evolution and biogeography of the group - i.a. they suggest that Bergia originated in Africa, moved to North America, and thence to Australia. The stem groups of Bergia and Elatine are estimated to be ca 35 and 40 Ma respectively, and much of the diversification in the genera has a strong geographic component, rather unexpected for aquatic taxa (Cai et al. 2016; see also Table S3 for rather different ages).
The flowers are small, probably associated with their autogamy, and Bonif´cio et al. (2023: esp. Table 1) discuss their morphology and anatomy in some detail. They described the endothecium as being thin-walled, although they show that of both genera as having transverse thickenings (ibid. Fig. 3H, 7G).
Pollination & Seed Dispersal. Autogamy is common here (Bonif´cio et al. 2023).
Chemistry, Morphology, etc.. Eichler (1878) draws three-merous flowers of this family with the odd sepal abaxial, i.e., in the monocot position, and W. Zhang et al. (2010: Supplement 1) consider that this orientation also occurs in flowers of the 5-merous Bergia texana. Kubitzki (2013b) recorded the ovules as being epitropous; they are shown as being apotropous in the literature that I have seen. Friesendahl (1927) suggested that the mesotegmen was lignified in Elatine while Dathan and Singh (1971 and references) thought that the seed coat of Bergia was exotegmic.
For general information, see Kubitzki (2013b); for embryology, see also Kajale (1940a), Raghavan and Srinivasan (1940a) and Tobe and Raven (1983b: summary).
Phylogeny. The African Bergia capensis and the European Elatine alsinastrum are sister to the rest of their respective genera, but classical infrageneric groupings do not correlate that well with phylogeny (Cai et al. 2016). For relationships within Elatine, see Razifard et al. (2017).
MALPIGHIACEAE Jussieu, nom. cons. - Back to Malpighiales
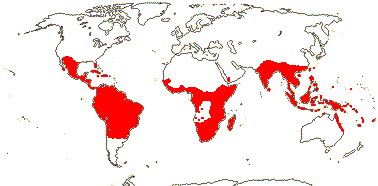
Shrubs or trees; (inulin +), ellagic acid 0; sclerenchyma as sclereids; cork subepidermal; pits vestured; (nodes 1:1); petiole bundle arcuate, wing bundles +; cuticle waxes as rosettes; stomata usu. paracytic; branching from current flush; hairs unicellular, ± T- or Y-shaped, surface rough (smooth); leaf venation brochidodromous, leaf glands common, abaxial and petiolar, stipules cauline, intrapetiolar and hooded or petiolar; inflorescence various; flowers monosymmetric (polysymmetric), banner petal adaxial [plane of symmetry oblique]; oil flowers, K with pairs of large abaxial oil glands (0, esp. Old World taxa), C clawed, often crumpled in bud, often fringed, one adaxial-lateral often different to the others; A obdiplostemonous, often basally connate, anthers basifixed, connective elaiophores + [of globose cells, oils non-volatile]; tapetal cells multinucleate; G [3(-5)], styles +, (connate); ovule 1/carpel, apical, pendulous, epitropous, micropyle exo/endo/bistomal, (apex of nucellus exposed), outer integument 2-4 cells across, inner integument 3-6 cells across, parietal tissue 10< cells across, nucellar beak +, (epistase +), hypostase +, (suprachalazal area massive), (pachychalazal); megaspore mother cells 1-several, embryo sac tetrasporic, 16-nucleate [Penaea type], (bisporic, 8-nucleate); K and A often persistent in fruit; seed smooth, exotegmen 0; endotegmic cells (elongated), lignified; (endosperm pentaploid), chalazal endosperm haustoria +, (embryo spirally coiled), cotyledons incumbent; x = 6 (?5), nuclear genome [1 C] (0.069-)0.918(-12.261) pg; duplication of CYC2-like gene.
72/1,499: [list, also see website - two subfamilies, 12 tribes below. Tropical and subtropical, especially American. Map: C. C. Davis, from Arènes (1957), Anderson (2011) and Australia's Virtual Herbarium (consulted xii.2013). [Photo - Flower, Flower, Fruit.]
Age. The age of crown-group Malpighiaceae has been estimated at (39-)36, 32(-29) Ma (Wikström et al. 2001), 75-64 Ma (Renner & Schaefer 2010), or ca 68 Ma (Davis & Anderson 2010); ages of (69-)59.8(-52.5) Ma were suggested by Xi et al. (2012b: Table S7), (93.2-)88, 80(-70.6) Ma by L. Cai et al. (2016) and ca 62.4 Ma by Cai et al. (2025b).
The earliest fossils attributable to Malpighiaceae are from the Northern Hemisphere in the later Eocene Claiborne Formation in Tennessee, U.S.A.; the deposits are ca 43 Ma old (Taylor & Crepet 1987: Eoglandulosa; see also Friis et al. 2011), Early Eocene fruits ca 52 Ma and placed in Tetrapterys have recently been described from Argentina (Siegert et al. 2025).
1. Byrsonimoideae W. R. Anderson
Vessels radially arranged, foraminate perforation plates 0, septate fibres +, rays 1-3 cells wide, with procumbent body cells, erect/square marginal cells/cells mixed, axial parenchyma marginal, prismatic crystals + (0); bracteoles glandular; adaxial C eglandular; A homomorphic, connective elaiophores prominent; style subulate, stigma punctate; fruit a drupe or schizocarp, smooth, not winged.
Age. An approximate age for crown-group Byrsonimoideae is 78-77 Ma (L. Cai et al. 2016).
1A. Galphimieae Niedenzu / the Galphimioid clade
Naphthopyrans, oxazinanes +; woof fibres thin- to thick-walled, articulated laticifers +; C keeled in bud; anther projections lateral.
5/40: Galphimia (26). Tropical America.
Age. Thryallis eocenicus (T. = Galphimia) was recently described from Early Eocene lignite deposits in Rajasthan Western India that are 54-52 Ma (Ali et al. 2025a).
[Acmanthereae + Byrsonimeae]]: ?
Age. The age of this clade is around 47 Ma (de Almeida et al. 2024b).
1B. Acmanthereae W. R. Anderson
Diazanapaphthalenes, propargyl-type 1,3-dipolar compounds +, benzopyrans, lactams, lignam glycosides, pyrimidine nucleosides, pyrimidine nucleotides, saccharolipids, sulfenyls 0; stipules 0; leaf venation camptodromous; K enclosing C in bud, (glands 0); G free, styles not apical.
3/23: Pterandra (15). South America, 1 sp. Panama.
1C. Byrsonimeae W. R. Anderson
Hydroxy acids and derivatives, imidolactams, keto acids, organic phosphoric acids, organofluorides, galloyl esters +, oxanes 0; (vessels not radially arranged), (foraminate perforation plates +), (septate fibres 0), rays often 4-5 cells wide, (axial parenchyma 0); phloem (stratified), sclerenchymatous cells varied; (sclerified phelloderm +, cells with U-shaped thickenings; leaves eglandular, stipules petiolar; C cucullate; fruits dry or fleshy.
3/181: Byrsonima (135-164). Tropical America, esp. Brazil.
Age. Crown-group Byrsonimeae are ca 40 Ma (de Almeida et al. 2024b).
2. Malpighioideae Burnett
Cincinni 1-flowered; adaxial C with nectar gland(s); A heteromorphic; style capitate, uncinate, truncate or expand-rarely subulate, stigmas capitate, crateriform (punctate); fruits nuts or schizocarps, winged, setose (smooth).
2A. Acridocarpeae R. F. de Almeida
Shrubs, trees, lianes; diazanaphthalenes, isoflavonoids, oxacyclic compounds +, tetrahydrofurans 0; leaves spiral to subopposite, stipules 0; main axis of inflorescence deflexed; bracts with 1-2 glands (usu.); adaxial C 2, (glands 0); anthers poricidal, pollen 3-syncolporate; G [2], (inferior - Acridocarpus), styles 2, deflexed in flower, reflexed in fruit; n = 9.
2/36: Acridocarpus (34). Africa to Madagascar and Iran, Thailand, West Malesia, also New Caledonia.
Age. Crown-group Acridocarpeae are ca 42 Ma (de Almeida et al. 2025).
2B. Mcvaughieae R. F. de Almeida
Linear 1,3-diarylpropanoids +, dithiols, indoles 0; vessels in radial multiples, perforation plates simple; stipules completely connate; cincinni 2-7-flowered; bracteoles with glands; C cucullate; pollen 4-zonocolporate (3-zonocolporate - Glandonia); stigma lateral; ovule 1; fruit a drupe, epicarp twisted; adaxial chamber with seed, abaxial chamber with oily substance; n = ?
3/12: Burdachia (6). Tropical South America, not Andean.
Age. The crown-group age of Mcvaughieae is around 25-15 Ma (C. C. Davis et al. 2014).(with sister - 38-33.9 Ma).
[Barnebyeae [Ptilochaeteae [Bunchosieae [Hiraeeae [Hiptageae [Malpighieae + Gaudichaudieae]]]]]]: ?
2C. Barnebyeae R. F. de Almeida - Barnebya W. R. Anderson & B. Gates
Diarylheptanoids, keto acids, oxazinanes only; cincinni 2-3-flowered; bracteole glands 0; adaxial C glands 0; pollen 4-zonoporate; fruit a schizocarp, samaras.
1/2. Brazil.
[Ptilochaeteae [Bunchosieae [Hiraeeae [Hiptageae [Malpighieae + Gaudichaudieae]]]]]: ?
2D. Ptilochaeteae R. F. de Almeida
2-aryl-benzofuran flavonoids, dibenzyl-butane lignans, isoflavonoids, oxacyclic compounds, oxanes, pyrrolidines, thiocarbonyls +; lamina margin revolute when young; pollen 8-zonocolporate.
3/10: Lasiocarpus (4). Dry forests, Mexico and pretty much throughout South America.
2E. Bunchosieae R. F. de Almeida
Habit various; azolidines, benzodioxoles, organochlorides, quinolizines +, organic carbonic and phosphoric acids 0; leaves glandular near base (not glandular, glandular along margin), stipules petiolar; abaxial C limb/claw ± hairy; pollen 4–12-pantoporate (colporate - Echinopterys, Heladena); mericarps unwinged.
5/122: Bunchosia (93), Tristellateia (21). Neotropical, Tristellateia Palaeotropical.
2F. Hiraeeae A. de Jussieu
Lianes; piperidines +, benzofurans, benzopyrans, dithiols, furanoid lignans, hydroxy acids etc., naphthopyrans, pteridines etc., pyrimidine nucleosides 0; lamina apex glandular, tertiary veins scalariform, stipules petiolar; C margins fimbriate [?all]; (pollen porate - Psychopterys); styles divergent, apex uncinate, stigma lateral.
5/105: Hiraea (81). Mexico to South America, not the Antilles.
[Hiptageae [Malpighieae + Gaudichaudieae]]: ?
2G. Hiptageae de Candolle / the Tetrapteroid clade
Habit various; organic phosphonic acids 0; (articulated laticifers + - Tetrapterys); lamina adaxial basal glands + (0), stipules inconspicuous (0); (bracteoles or K enclosing flower); K valvate/imbricate, 2-10 glands, oil secretion - New World, 0-10 glands, nectar secretion - Old World, C pubescent; A 9 long, 1 short/vice versa/subequal; (pollen porate - Hiptage, some Heteropterys); (styles 1, 2), stigmas lateral; fruit with 4 free lateral wings [?apo.].
17/377: Heteropterys (166), Hiptage (47), Tetrapterys (46), Glycophyllum (28), Niedenzuella (18). Pantropical.
Age. The Tetrapteroid clade is (35-)34(-33) Ma (de Almeida & van den Berg 2022).
Fruits of Tetrapterys dolgopolae, early Eocene in age (ca 52 Ma) and clearly likely to have New World affinities, have recently been described from Laguna del Hunco, Argentina (Siegert et al. 2025), the authors noting that this was a well-known site in which the fossils had hitherto overwhelmingly shown Indo-Pacific relationships.
[Malpighieae + Gaudichaudieae]: stipules interpetiolar.
2H. Malpighieae de Candolle
(3'–>5')-dinucleotides, piperidines, monofluoroacetates +, benzofurans, furanoid lignans, imidolactams, lignan glycosides 0; (phloem stratified [± sclereidal]); (plants dioecious); (4-12 pantoporate - Palaeotropical spp.); stigmas lateral; fruit winged [dorsal, lateral encircling fruit, or 4 lateral], (bristly), (drupe, pyrenes separate); n = (9) 10.
13/253: Malpighia (110), Mascagnia (48), Microsteira (27), Aspidopterys (24). Pantropical, esp. the Americas, subtropical.
Age. Crown-group Malpighieae may be around 74 Ma (Cai et al. 2016).
2I. Gaudichaudieae Horaninow / stigmaphylloid clade
Macrolactams +, biotin and derivatives, sulfenyls 0; (articulated laticifers + - Stigmaphyllon); druses + (prismatic crystals - Banisteriopsis); (petiole with 4 wing bundles); (leaf blades palmately lobed/-veined, margins crenate/serrate - Stig.); inflorescence umbellate (not); A (homomorphic - Bronwenia), (2 + 3 staminodes [Cottsia]-)5; connective glands prominent [with overlapping globose cells/secretory papillae]; (pollen porate - Stig. subg. Ryssopterys, Philgamia, Sphedamnocarpus); (integument 1, 3-5 cells across - Janusia); mericarps with dorsal (2 lateral) wing.
14/336: Stigmaphyllon (119), Banisteriopsis (65), Diplopterys (31), Peixotoa (29), Aspicarpa (27). Pantropical.
Evolution: Divergence & Distribution. An origin of the family in South America during the Late Cretaceous has been suggested, with several - it now appears to be nine - subsequent migration/dispersal events to the Old World often followed by the loss of oil secretion by the sepals in the taxa involved (Davis et al. 2002a, b, 2004; esp. Davis & Anderson 2010; see also below). Bonifácio et al. (2021) suggest oil glands have been lost at least 14 times in Malpighiaceae, including seven times in Neotropical taxa. To summarize diversity/diversification in the family: Most species are Neotropical (about 90% of the total), where overall floral diversity is relatively less and fruit diversity is greater, and while there are far fewer Old World taxa, overall floral diversity is greater there (Davis & Anderson 2010: Fig. 3; Davis et al. 2014b). Davis et al. (2014b) attribute this relative invariance in the New World malpigs to stabilizing selection rather than some kind of inherent developmental stasis, furthermore, the rate of diversification may have increased in Malpighiaceae (Xi et al. 2012b). This latter is explained in part by the small geographic ranges of most of its solitary oil bee pollinators, the male bees being territorial, nevertheless, Malpighiaceae clades tend to have wide geographic ranges, hence allopatric speciation (Davis 2014b). De Almeida et al. (2024b) examined diversification in Byrsonima; sampling was a bit exiguous - ca 1/5 the species - but there seems to have been a fairly long phylogenetic fuse of ca 29 My, and diversification of the Cerrado species (the genus is diverse there) began ca 7.7 or 6.0 Ma, in line with that in other Cerrado genera.
Malpighiaceae in Mexico are very diverse in seasonally-dry tropical forest, and this is because of the numerous (over 30) dispersal events from the south as much as any subsequent diversification of these clades. Initially animal-dispersed clades (more l.d.d.) were involved, but there was an increase in wind-mediated dispersal events in the mid-Miocene ca 24 Ma when connections with South America may have been established (Anderson 2013; Willis et al. 2014a [see also B. Bremer & Eriksson 1992]; Bacon et al. 2015a; Montes et al. 2015: geology re-evaluated, but c.f. Proc. National Acad. Sci. 112(43): E5765-E5678 and especially O'Dea et al. 2016: comprehensive refutation).
Although Malpighiaceae are primarily New World, recent fossil discoveries in India make one think. Fossil fruits of Thryallis (= Galphimia) have been reported from 54-52 Ma Eocene deposits in western India (Ali et al. 2025a), and although the climate in that part of the world has changed considerably since then, no extant Byrsonimoideae, to which Thryallis belongs, are known from the Old World. Hiptage is the largest and most diverse Palaeotropical member of Malpighiaceae; with a centre of diversification in Vietnam/IndoChina, the genus may have arrived there ca 23 Ma, perhaps via long-distance dispersal and by way of North America and Europe (de Almeida & van den Berg 2022). Dicella (Hiptageae) suggests complications: It is a New World genus, but recently Dicella indica was described from Pliocene fossil fruits in eastern India (Hazra et al. 2024). Acridocarpeae are another Old World clade, and de Almeida et al. (2025, q.v. for optimization of 20 morphological characters on the tree) suggest that they arrived there via a Gondwanan dispersal route, not Boreotropical. Acridocarpus itself is made up of two main clades, one from the African mainland, the other from Madagascar (crown-group age ca 32.2 Ma), but A. austrocaledonica (it diverged ca 10.4 Ma) is from Mew Caledonia (de Almeida et al. 2025).
Anastomosing laticifers, not articulated in the Galphimia clade but articulated in the other cases and varying in distribution within the plant (remember, the articulated/non-articulated distinction is increasingly appearing to be illusory), have evolved three times in Malphigiaceae. They contain alkaloids, tannins, terpenoids, etc. (Pace et al. 2019), although isoprenes, a component of latex s. str., are not mentioned. Sanchez et al. (2025) looked at a number of characters of wood anatomy in the Galphimieae; they suggested that their characters were discrete, although it is unclear if the states of those characters are, thus Galphimieae were characterized by having thin- to thick-walled versus very thick-walled fibres. Perhaps unsuprisingly, Galphimieae were divided into two groups characterized by plant habit.
R. F. Almeida and van den Berg (2021) placed a number of characters on the phylogeny they obtained for the Tetrapterys group (Malpighioideae). De Almeida et al. (2024a) optimized variation in 31 characters of external morphology across the tree of the family; the results they found at the lower nodes are incorporated into the characterizations above, but potential apomorphies for a number of genera/generic groups were also suggested. Bueno et al. (2024) looked at the variation of wood and cork anatomy of Byrsonimeae in particular, but putting it in the context of that of Byrsonimoideae as a whole; variation here was conservative, and that that did occur showed much homoplasy. Chemical variation within Byrsonima was also rather limited (Santos-Zanuncio et al. 2021), and neither group of characters seemed to have much to do with infrageneric classification here..
Recent work using three molecular markers, nrITS, rbcL and matK, suggests that the type specimen of Keraunea brasilensis, a genus quite recently described in Convolvulaceae, albeit it had some rather odd characters (Cheek & Simão-Bianchini 2013), is to be placed in Malpighiaceae, where it associates with Mascagnia, and with good support (Muñoz-Rodríguez et al. 2022). In the context of Malpighiaceae, it has very odd characters indeed - note its leaves, corolla, anthers and gynoecium. However, this is probably because an errant leaf, probably of M. cordata, inadvertently associated with the type specimen was sampled (de Almeida et al. 2023a). A paratype of K. brasilensis was placed in Ehretiaceae, and recent work suggests that Keraunea is indeed to be placed in Ehretiaceae (Moonlight & Cardoso 2023; Cheek et al. 2023b).
Ecology & Physiology. Malpighiaceae are one of the three ecologically most important groups of lianas in the New World tropics, around 400 species being lianescent (see also Bignoniaceae-Bignonieae and Sapindaceae-Sapindoideae: Gentry 1991; additional information in Angyalossy et al. 2015). The main origin of the liane habit is at the Bunchosieae et al. node, and a number of Malpighiaceae in the clade involved have anomalous secondary thickening, phloem wedges evolving ten times or so, and the cambium beuing either continuous or discontinuous (Quintanar-Castillo & Pace 2022: esp. Fig. 2).
Pollination Biology. New World Malpighiaceae are noted for having oil flowers, a trait that is probably plesiomorphic in the family. Oil is secreted by epithelial elaiophores, often paired, that are found on the backs of usually four of the sepals - not the abaxial sepal (Possobom & Machado 2017a, b and references; Tölke et al. 2019; Aliscioni et al. 2022), although genera like Galphimia lack such glands (Bonifácio et al. 2021). (Note that flowers of New World Malpighiaceae have a "monocot" orientation, the flower being rotated 36o, and so the odd petal, the banner petal, is adaxial - W. Zhang et al. 2010.) Tapinotaspidini, Tetrapediini and solitary Centridini (Epicharis, Centris: paraphyletic, see below), all restricted to the New World, are the pollinators there. Centridini bees have tufts of hairs on four legs that the insects use to get the oil from the glands (A. C. Martins et al. 2014), at the same time holding on to the flower by grasping the narrow base of the adaxial banner petal with their mandibles; this banner petal is often distinctively coloured, and may change colour as the flower ages (Renner & Schaefer 2010 and references). At least some species with flowers that are apparently yellow - yellow flowers are common in the family - are in fact bee UV-green (Papadopulos et al. 2013). Bezerra et al. (2009) discussed the pollination networks formed by oil bees and plants, and found them to be very resilient to the loss of species of either group (see also Mello et al. 2012: 75 bee and 64 malpig spp.), although activities of the bees on flowers other than those of malpigs were not discussed. Note that these centrid bees also collect resins. Trigonid bees visit the flowers of New World malpigs for pollen (Anderson 1979), and some taxa are buzz pollinated (Sigrist & Sazima 2004).
The flowers of some Orchidaceae-Oncidiinae mimic (both Batesian and Müllerian mimicry may be involved) those of Malpighiaceae, having similarly-coloured flowers - bee UV-green (see esp. Papadopulos et al. 2013; Pansarin et al. 2021). The orchids may even provide distinctive fatty acids as a reward that are similar to those of Malpighiaceae (Reis et al. 2007 and references). The mimicry unit of the orchid is formed largely by the labellum, with the column being visually similar to the malpighiaceous banner petal. Other Oncidiinae have a distinctive shiny green nectary-look alike on the labellum, which may otherwise be white. Castro et al. (2021) tested this mimicry hypothesis comparing Gomesa flexuosa (Epidendroideae-Cymbidieae-Oncidiinae) and Janusia guarantica (Malpighiaceae) and suggested that such orchid flowers, which also occur in other oncidiine orchids that do not grow with Malpighiaceae, were rather exploiting the sensory biases (e.g. flower colour, rewards) of their pollinating bees (see also Ruxton & Schaefer 2011).
Cardinal and Danforth (2013) suggested that Apinae-Centradini (Centris 230 spp.; Epicharis 35 spp.) and Xylocopinae-Tetrapediini-Tetrapedia stem lineage, bees which take oil from Malpighiaceae, evolved in the Late Cretaceous, 87-52 and 92-66 Ma respectively, and these ages and the malpig family ages above are broadly consistent (see also Bossert et al. 2018: monophyletic; Cardinal 2018: some ages). The first two of these genera probably originated in humid forests (A. C. Martins & Melo 2016). However, Martins et al. (2014a; see also Hedtke et al. 2013) found Centradini to be paraphyletic, Epicharis diverging (102-)91(-79) Ma and Centris (95-)84(-72) Ma, about contemporaneous with Xi et al.'s (2012b) estimate of the stem-group age of Malpighiaceae of (100-)86(-73) Ma (75-60(-32) Ma above), and broadly consistent with some kind of co-evolutionary story. There are other suggestions, e.g. that the crown age of Epicharis is (40-)28(-19) Ma and that of Centris is (59-)46(-36) Ma (Martins et al. 2014a) and 35-30 and 40-35 Ma respectively (Ramirez et al. 2011), rather younger than most malpig family ages. Fossils of Eoglandulosa warmanensis from the Eocene Claiborne Formation in Tennessee, U.S.A., from ca 34 Ma show the distinctive paired glands on the sepals (Taylor & Crepet 1987; Friis et al. 2011). Aguiar et al. (2019) discussed the evolution of Tapinotaspidini (Apidae, Apinae) oil bees, some 180 species of solitary bees that originated ca 57 Ma. Malpighiaceae appear to have been their ancestral hosts, Palaeocene cerrado-like savannas in South America their ancestral habitat, the bees soon moving on to different hosts and, much more recently, into forested habitats. However, details of the evolution of the association between bees and malpigs are somewhat unclear.
In addition to the calyx glands, Malpighiaceae have small osmophores on the fimbriate margins of the petals, as well as glands on the anther connectives, but these have a less clear role in pollination (Possobom et al. 2015). The morphology of the floral elaiophores and osmophores is variable, and they are to be found on the anther and/or connective (Possobom et al. 2015; Possobom & Machado 2017b; Arévalo-Rodrigues et al. 2020). However, a recent survey by de Almeida et al. (2023b: 46 genera included) suggests that connective glands are in fact widespread in the malpigs, perhaps an apomorphy for them, and they usually (apart from some species in two clades) consist of globose epidermal cells. Little is known of the composition and function of their products, but these appear to be non-volatile oils - and/or perhaps mucilage (Avalos et al. 2020: esp. the anthers of the small stamens - division of labour?), or a mixture of lipids, polysaccharides and phenolics - that help attach the pollen grains to the bees, or they may indeed be osmophores, attracting the bees (de Almeida et al. 2023b).
There are 150-250 species of Old World Malpighiaceae, relatively few compared to those in the New World. However, these Old World taxa show much more variation in basic floral morphology than their New World relatives, and this is associated with the adoption of different pollinators and pollination mechanisms. Old World taxa do not have oil flowers, and Renner and Schaefer (2010) suggested that there had been at least six losses of oil glands in African clades (all told calyx glands have been lost at least 14 times in the family, including in a few New World taxa - Bonifácio et al. 2021). Their flowers may be radially symmetrical and/or have a "normal" floral orientation for a core eudicot with the odd petal abaxial (W. Zhang et al. 2010), and this has happened independently in separate clades (Davis et al. 2014b). Polysymmetry is associated with changes in the expression of the CYC2 gene, and these differed in each case of the evolution of polysymmetry studied (W. Zhang et al. 2013). Hiptage, especially H. benghalensis, shows very pronounced monosymmetry with unequal stamen lengths and anther sizes (heteranthy), an adaxial banner petal, and a single nectar-secreting gland between the two adaxial sepals; this is associated with the presence of four CYC2-like genes (W. Zhang et al. 2016: see cover photo of Internat. J. Plant Sci. 177(7). 2016). The calyx glands may secrete nectar (Vogel 1974, 1990; M.-X. Ren et al. 2013; Zhang et al. 2016: the median gland in Hiptage secretes oils in some species), although in most pollen is the only obvious reward (Davis & Anderson 2010). The anthers may be porose or have slits, and the flowers are on occasion mirror images and have a single stamen much larger than the rest (Ren et al. 2013). The flowers tend to be more or less polysymmetric, the petals are less strongly clawed, and the style branches are longer and more widely spreading (Davis & Anderson 2010: Fig. 3 for illustrations).
Self-fertilization is common in species of Gaudichaudia, Janusia and relatives. Here pollen tubes grow from the indehiscent anther through the tissues of the flower to the embryo sac (Anderson 1980; X.-F. Wang et al. 2011). Apomixis - nucellar polyembryony - is common, and embryo sac development and reproduction in general is very variable; as Johri et al. (1992: p. 450) noted "failure of fertilization is a common feature of Malpighiaceae".
Plant-Animal Interactions. Byrsonima is considered to be a keystone species, along with Cecropia and in particular Miconia, in the context of the maintainance of general plant—frugivore (bird) interactions throughout the Neotropics (Messeder et al. 2020a).
Genes & Genomes. There have been duplication(s) of CYC genes, and CYC2 genes are expressed only in the adaxial part of the flower in monosymmetric Malpighiaceae. However, details of exactly when gene duplication occured and what changes in pattern of gene expression there have been are unclear (W. Zhang et al. 2010), although Cai et al. (2017/18) found that Galphimia (Tristellateia sister) may have two duplications, one quite recent, and Landis et al. (2018) dated a duplication that involved the whole family to 69.8 Ma - this was the GAGRα event.
Chemistry, Morphology, etc.. Acridocarpus has spiral, exstipulate leaves. Some species of Stigmaphyllon have leaves with palmate venation and toothed margins; some taxa, especially when young, have almost fimbriate lamina margins, albeit distantly so (the fimbriae are ca 4 mm long). Stipules are very diverse, being petiolar in Hiraea and cauline in many vines and also in Malpighia; in the latter genus they may be lobed or toothed. Glands on the leaves are common, and in Galphimia brasiliensis the subbasal glands on the lamina margins have the same anatomy as small glands on the sides of the sepals towards the base, both secreting oil (Castro et al. 2001). However, Lobreau-Callen (1989) recorded the leaf/bracteole glands of G. bracteata as producing sugars.
The flowers of Acridocarpus have an inferior ovary with only two fertile carpels. Although Lorenzo (1981) suggested that the nucellar beak in Janusia was produced by periclinal divisions of the epidermal cells, i.e., it would technically be a nucellar cap, the cell wall patterns do not suggest this.
Souto and Oliveira (2020) summarize what is known about fruit anatomy in the family; lignification of the inner part of the mesocarp and the endocarp, which can be multiplicative, is quite variable. Given that the main protective part of the propagule is not the seed coat, testa and tegmen are likely to be more or less reduced, however, seed coat anatomy may repay investigation. In Mascagnia macrodisca there is a layer of thin-walled, slightly elongated cells with brown contents over a layer of more or less isodiametric, lignified cells with somewhat more thickened and straight anticlinal walls, the latter layer having a "frosted" appearance (pers. obs.); I do not know the origin of these layers. Banisteriopsis has vascular tissue in the testa and the seed is more or less exotestal (Souto & Oliveira 2008). Endotegmic fibres may be quite conspicuous (Souto & Oliveira 2006). However, since fibres in other Malpighiales are exotegmic and Elatinaceae also have exotegmic seeds (but see above), what is going on in Malpighiaceae is unclear.
C. Anderson et al. (2006 onwards) provided vast amounts of general information, especially on phylogeny and nomenclature, and links to papers for the whole family; for Mcvaughia, see de Almeida et al. (2019). Some information on chemistry is taken from Hegnauer (1969, 1989: iridoids have been reported from Stigmaphyllon), Lee et al. (2012: monofluoroacetates) and Mannochio-Russo et al. (2022); for interxylary cambia, distinctive stratified phloem, etc., in climbers, see Cabanillas et al. (2017) and Pace et al. (2018a), for some leaf anatomy, see Mello et al. (2019) and Vilarinho et al (2023), for embryology, etc., see also Stenar (1937), A. M. S. Rao (1941 and references), Subba Rao (1980) and Souto and Oliveira (2005, 2008), for fruit and seed, see Takhtajan (2000), and for seedlings, see Barbosa et al. (2014).
Phylogeny. There is information on relationships within the family in Davis et al. (2001), Cameron et al. (2001) and Davis and Anderson (2010: all genera, 3 plastid, 1 nuclear genes). Within Malpighioideae, for the most part well-supported branches including Acridocarpus, McVaughia, Barnebya, Ptilochaeta, [Bunchosia, Tristellateia] and Hiraea (these are the main genera in the clades) are successively sister to the remaining taxa. A study by Davis and Anderson (2010: four genes, all genera sampled) returned the same set of relationships, and with good support; there are many other well-supported clades. For some details of relationships, see also M. Sun et al. (2016) and Cai et al. (2016). De Almeida et al. (2024a: 3 plastome and 1 nuclear genes, all genera, to 3 species/genus) looked at relationships throughout the family and retrieved the for the most part quite well supported relationships that are the basis of the classification above; of course generic limits could not be examined there.
De Almeida and van den Berg (2021: 4 chloroplast and 1 nuclear markers) looked at relationships in the Tetrapterys clade.
Acridocarpeae. De Almeida et al. (2025: 21/34 spp., ITS and 5 plastid genes) looked at relationships here, recovering two main clades; previous relationships did not hold up.
Within Byrsonimeae, relationships are [Galphimia group [Acmanthera group + Byrsonima group]] (Davis & Anderson 2010). De Almeida et al. (2024b: 33 species, 1 plastid + 2 nuclear markers) obtained substantial resolution of relationships within Byrsonima; all the currently-recognized infrageneric groupings were polyphyletic.
Hiptageae. De Almeida and van den Berg (2022: ITS, 16/44 spp. examined) found that the subgenera of Hiptage were all polyphyletic.
Classification. Recently there have been substantial modifications to generic limits (e.g. de Almeida & van den Berg 2021 and references). Van Do et al. (2024: ITS data) found that a new species was sister to Hiptage, and they placed it in a new genus, Chlorohiptage, but not only was the sampling only moderate (12 spp. Hiptage) but support along the backbone of the tree there was rather poor. De Almeida et al. (2024a) provide a formal classification with full synonymy for the whole family that is followed above, also a species-level check list, and so on. A new infrageneric classification for Acridocarpua has been proposed - 3 subgenera (de Almeida et al. 2025).
Previous Relationships. The relationships of Malpighiaceae have been difficult to work out; they were included in Vochysiales by Takhtajan (1997) and in Polygalales by Cronquist (1981).
[Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]] / Chrysobalanoids: hairs simple [unicellular always?]; ovules collateral, micropyle bistomal, outer integument ≥5 cells across, inner integument ≥5 cells across, nucellus evanescent by maturity, endothelium +; endosperm at most slight, embryo chlorophyllous.
Age. This node can be dated to (106)100(-95.5)/(95-)90(-88.5) Ma (Davis et al. 2005a), (94.9-)83.5(-74.8) Ma (Xi et al. 2012b: Table S7), ca 76 Ma (Tank et al. 2015: table S2), (83-)71-70(-58) Ma (Bell et al. 2010), or (62-)59, 57(-54) Ma (Wikström et al. 2001).
Evolution: Divergence & Distribution. There are polarization problems again: Is the fruit plesiomorphically a drupe, with transitions to septicidal capsules, or are fruit types apomorphies of individual families...? For a comparison of general morphology within the clade, see Litt and Chase (1999), for floral morphology in particular, see Matthews and Endress (2006a); for ovule position, see Merino Sutter and Endress (2003).
Chemistry, Morphology, etc.. Tobe and Raven (2011) suggested that there is a multiplicative inner integument; however, although it is thick, it does not usually become thicker after fertilization. There is insufficent information to know if a vascularized testa might be an apomorphy for the clade.
For pollen, see Furness (2013b).
Phylogeny. For relationships, see especially Litt and Chase (1999). This group (= chrysobalanoids: Xi et al. 2012b) has held together strongly in subsequent studies.
BALANOPACEAE Bentham & J. D. Hooker, nom. cons. - Balanops Bentham & J. D. Hooker - Back to Malpighiales
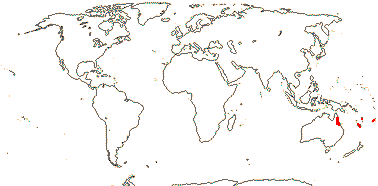
Trees; ellagic acid?, prob. tanniniferous; vessel elements long [usu. 1,000 µm<], with scalariform perforation plates; parenchyma diffuse; sclereid nests with rhomboidal crystals in bark; rhomboid crystals +; petiole bundle?; (cristarque cells in leaf); buds perulate; cuticle waxes 0 (platelets); stomata usu. laterocytic; leaves spiral, lamina tooth ?type, stipules minute; plant dioecious; staminate plants: inflorescence catkinate; P +, uniseriate, of small teeth; A (1-)3-6(-14), anthers much longer than filaments; pollen 3-5-colpate, microechinate, exine columellate-granulate; pistillode common; carpelate plants: inflorescence fasciculate; flowers with cupule made up of spirally arranged bracts; P 0; staminodes 0; G [(2)], styles long, once or twice bifid, stigma adaxial on styles, ?type; ovules subbasal, apotropous, facing laterally, outer integument 5-7 cells across, inner integument 5-9 cells across, parietal tissue ca 2 cells across; fruit a drupe, with 2-3 stones; testa vascularized, persistent, cell walls not much thickened; endosperm ?type, slight, embryo large, cotyledons cordate; n = 20 (21), x = ?7 (?6, ?11); germination epigeal, phanerocotylar, cotyledons with single adaxial papilla.
1 [list]/9. S.W. Pacific, especially New Caledonia. Map: from van Steenis and van Balgooy (1966). [Photo - Fruits © Andrew Ford, CSIRO.]
Evolution: Divergence & Distribution. Some analyses suggest that the diversification rate in Balanopaceae decreased (Xi et al. 2012b) - ummm.
Chemistry, Morphology, etc.. The leaves are often described as being dimorphic (Carlquist 1980; Cronquist 1983), but they are no more so than in many plants that have perulate buds.
Pollen descriptions in Feuer (1991) and Herendeen et al. (1995) differ somewhat.
For general information, see Carlquist (1980) and Kubitzki (2013b); Batygina et al. (1991) provide details of testa anatomy, Merino Sutter and Endress (2003) of the floral morphology of carpelate flowers.
Previous Relationships. Relationships of Balanopaceae were for long problematic. Cronquist (1983) compared their wood anatomy with that of Hamamelidaceae, Balanopales were included in Daphniphyllanae by Takhtajan (1997), while Merino Sutter and Endress (2003) found that many features of the female flowers were consistent with a position in Malpighiales, Balanopaceae perhaps being somewhat similar to Euphorbiaceae s.l..
[[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]: vessel elements with simple perforation plates; vestured pits +; mucilage cells +; stomata paracytic; lamina margins entire, (flat surface glands or glandular hairs +); pedicels articulated; flowers obliquely monosymmetric; K basally connate, with epidermal mucilage cells, quincuncial, 2 outer members shorter, C unequal in size; fertile stamens abaxial, ± connate, anthers with a little pit where the filament joins, connective well developed abaxially with endothecium continuous there, staminodes adaxial; G with longitudinal furrows, unicellular unlignified hairs +, style +, stigmas commissural; ovules with zig-zag micropyle, outer integument 2-5 cells across, inner integument 3-8 cells across, parietal tissue?, obturator +.
Age. This node can be dated to (53-)50, 41(-38) Ma (Wikström et al. 2001) and (72-)60, 59(-46) Ma (Bell et al. 2010).
Evolution: Divergence & Distribution. Matthews and Endress (2008) elaborate the floral morphology of this clade and suggest synapomorphies for its members.
Previous Relationships. Including these four families in Chrysobalanaceae s.l. was optional in A.P.G. II (2003), and specimens of Chrysobalanaceae and Dichapetalaceae are quite often misidentified as the other family (G. T. Prance, pers. comm.), however, the families are kept separate by Prance and Sothers (2003a), A.P.G. III (2009) and A.P.G. IV (2016).
[Trigoniaceae + Dichapetalaceae]: (vessel elements with scalariform perforation plates); petiole bundle arcuate; lamina with secondary veins strongly arching towards the margin; inflorescences cymose; K with mesophyllar mucilage cells; nectary with lobes or scales, semi-annular, apparently outside A [staminodial?]; ovary and lower style completely synascidiate; testa multiplicative.
Age. The age of this node may be (43-)40(-37), (32-)29(-26) Ma (Wikström et al. 2001), (66-)50, 48(-34) Ma (Bell et al. 2010), ca 57 Ma (Tank et al. 2015: table S2), or (71.4-)59.7(-47.4) Ma (Xi et al. 2012b: Table S7).
TRIGONIACEAE A. Jussieu, nom. cons. - Back to Malpighiales
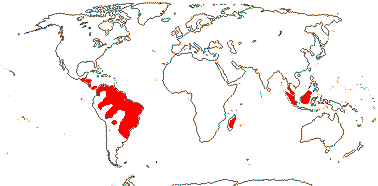
Tall trees or shrubs, lianas; helical thickening in ray and axial parenchyma; wood fluorescing [1 sp]; parenchyma in apotracheal bands; branched sclereids +; (nodes with split laterals); hairs unicellular, T-shaped or not; leaves opposite, spiral or two-ranked, lamina with dense whitish hairs below (not), stipules interpetiolar [when leaves opposite]; (inflorescence racemose - Isidodendron); C contorted, adaxial-lateral petal basally spurred or saccate [the standard], plicae in abaxial + abaxial-lateral petals form the keel, or these petals saccate; A 5-13, filaments ± connate, fertile stamens 4-9, adaxial, endothecial cells extending over the back of the connective, staminodes 0-6; pollen 3-5-porate; nectary of 1 or 2 [and then each to 3-lobed] glands at base of standard (glands on base of staminodes - Isododendron); G [(4)], median member adaxial, placentation also parietal, stigma capitate to slightly trilobed, papillate; ovules 1-10/carpel, (apotropous), micropyle (endostomal); outer integument 2-3 cells across, inner integument 4-6 cells across, inner integument endothelial; fruit a septicidal capsule, valves opening internally, central fibrous strands persisting, (hairs from endocarp - Trigoniastrum), or samara; seeds (winged), exotesta with thickened outer walls, tanniniferous, with short to long lignified hairs (spines), (nothing - Humbertidendron), tegmen multiplicative, endotegmic cells tanniniferous, walls slightly thickened; endosperm +/0, development?; cotyledons large; n = ca 10; germination epigeal, phanerocotylar.
5 [list]/28: Trigonia (24). Central and South America, Madagascar (Humbertiodendron), W. Malesia (Trigoniastrum) (map: from van Steenis 1949c; Lleras 1978). [Photo - Flower.]
Age. Crown-group Trigoniaceae can be dated to (49.6-)31.6(-12.6) Ma (Xi et al. 2012b: Table S7 - sampling).
Chemistry, Morphology, etc.. Trigonia has opposite leaves, interpetiolar stipules, and split lateral vascular bundles; lamina glands are not obvious. The bracts of Trigoniastrum have large glands on the abaxial surfaces, Trigonia has stalked glands variously on pedicels or petioles and margins of bracts and leaves, while Humbertiodendron has concave marginal glands towards the base of the leaf blades.
For floral morphology, I follow the interpretations of Warming (1875) and Eichler (1878) rather than that of Cronquist (1981). Schnizlein (1843-1870: fam. 233) draws the flowers as being more or less vertically monosymmetric, while the orientation shown by Schatz (2001) is difficult to work out; in the latter it appears that the corolla may be quincuncial. Warming (1875) showed the nectary glands as being part of the androecial whorl; Lleras (1978) describes them as being "disc glands". In any event, the androecium at least sometimes seems to have more than 10 stamens. Kopka and Weberling (1984) also discuss floral morphology.
Some general information is taken from Lleras (1978), Takhtajan (2000), Fernández-Alonso et al. (2000) and Bittrich (2013); see Hegnauer (1973) for chemistry, Carlquist (2012c) for some wood anatomy, and Mauritzon (1936) and Boesewinkel (1987) for embryology, seeds.
Previous Relationships. Trigoniaceae were included in Vochysiales by Takhtajan (1997), while Cronquist (1981) placed them in Polygalales.
DICHAPETALACEAE Baillon, nom. cons. - Back to Malpighiales —— Synonymy: Chailletiaceae R. Brown
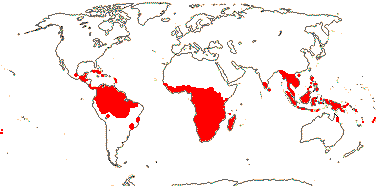
Trees or lianas; (monofluoroacetates, pyridine alkaloids +); xylem parenchyma ± paratracheal; sieve tubes with non-dispersive protein bodies; pericyclic sheath interrupted; fibres common; branching from current flush; hairs warty[?]; leaves spiral, (lamina with flat abaxial glands), stipules fimbriate or not; inflorescence epiphyllous, from petiole, (not); flowers small [≤6 mm across], (polysymmetric), (4-merous); C (connate), bifid (unlobed), (3 small entire petals forming a "lip" - Tapura), drying black; nectary a ring, or bilobed lobes opposite C; stamens 5, opposite K, (3 + 2 staminodes), (connate), (adnate to C), (anthers without pits); pollen (≤30µm), aperture fastigiate; G [(2-4)], (inferior), (styluli +), stigmas ± punctate, wet, papillate; ovules pendulous, outer integument 3-5 cells across, inner integument 6-8 cells across, hypostase 0, funicular obturator +; fruit a flattened drupe, 1(-3, then often lobed) locular, (loculicidal capsule); 1 seed/loculus, (arillate); testa vascularized, only enlarged tanniniferous (divided) exotestal cells and remains of vascular bundles persist, exotegmen 0; embryo (orange), oily; n = (?10) 12, chromosomes 1-2 µm long.
3 [list]/165: Dichapetalum (130). Pantropical, few in Malesia. Map: see Prance (1972b), 2022), Leenhouts (1957a), van Steenis (1963), Heywood (1978) and Trop. Afr. Fl. Pl. Ecol. Distr. vol. 2 (2006). Photo - Flower, Photo - Fruit.
Age. This node can be dated to (36-)20.7(-6.8) Ma (Bell et al. 2010 - inc. Tap).
Evolution: Divergence & Distribution. Some analyses suggest that the diversification rate in Dichapetalaceae may have increased (Xi et al. 2012b).
Chemistry, Morphology, etc.. Dichapetalum at least has fluoracetic and related acids in its seed oils and is often very poisonous as a result (e.g. Badami & Patil 1981; Lee et al. 2012); it also contains the cytotoxic dichapetalins, tetracyclic triterpenes (Long et al. 2013: also in Phyllanthus!). The petiole bundles are arcuate above the insertion of the inflorescence.
The flowers of Tapura in particular are quite complex (Prance 1972b). Some species of Dichapetalum are reported to have arils (Prance 2013).
Some information is taken from Prance (1972b - see 2022 for an update, 2013); see also Hegnauer (1966, 1989) for chemistry, Barth (1896) for petiole anatomy, Punt (1975) for pollen morphology, and Boesewinkel and Bouman (1980) for ovule and seed.
Previous Relationships. Dichapetalaceae were placed in Celastrales by Cronquist (1981) and in Euphorbiales by Takhtajan (1997).
[Euphroniaceae + Chrysobalanaceae]: flowers with widespread calcium oxalate crystals; hypanthium +, spurred, nectary on inside, unlobed; C clawed, with lignified hairs; style furrowed; embryo with chlorophyll.
Age. The age of this node is around (74.9-)66.2(-60.3) Ma (Xi et al. 2012b: Table S7), ca 86 Ma (Bardon et al. 2012), ca 53.5 Ma (Tank et al. 2015: table S2), or (56.8-)49.2(-42.3) Ma (Bardon et al. 2016).
Chemistry, Morphology, etc.. Whether or not this clade has a spur in the floral cup is a matter of perspective; I prefer to interpret the flower in many Chrysobalanaceae as having the gynoecium adnate to one side of the hypanthium rather than being spurred as may appear to be the case in a l.s. of the flower. Hairs have been found in the ovary loculi in flower, but they are not obvious in the capsule, even if it has not yet opened (pers. obs.).
EUPHRONIACEAE Marcano-Berti - Euphronia Martius & Zuccarini - Back to Malpighiales
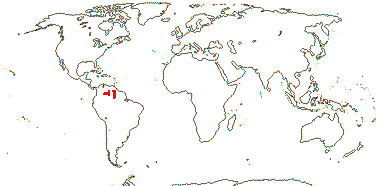
Tree; parenchyma ± aliform-confluent; petiole bundles annular or arcuate, (not joining immediately with stele); cortical and foliar sclereids +; hypodermis mucilaginous; leaves spiral, lamina with revolute vernation, white tomentose below; inflorescence terminal; bracteoles 0; C 3 [abaxial-lateral and abaxial C absent], contorted; disc 0; stamens 4-7, in two groups, adnate to C, filaments basally connate, staminodes one long, abaxial-lateral, retrorsely pilose, between the stamen groups, and 4-5 small, appearing dentate; G with median carpel adaxial, stigma clavate; ovule apotropous; fruit a septicidal capsule, columella persisting; seeds winged, 1/carpel, coat?; endosperm development?; n = ?
1 [list]/3. The Guyana Shield, South America, Map: from Steyermark (1987).
Evolution: Divergence & Distribution. Some analyses suggest that the diversification rate in Euphroniaceae decreased (Xi et al. 2012b).
Chemistry, Morphology, etc.. Lleras (1976) suggested that the (long) staminode of Euphronia was in the position of the fertile stamen of Vochysiaceae (now Myrtales, but that was at a time when the two were thought to be related) and Marcano-Berti (1995) that there was a staminal tube and four stamens of two different lengths. Prance and Sothers (2003: table comparing the four families) note that Euphronia lacks any disc.
Some information is also taken from Warming (1875) and Kubitzki (2013b), both general, Barth (1896: general anatomy), and de Pernia and ter Welle (1995: wood anatomy), and some more data come from Euphronia guianensis: Colonnello-Medina 712, vegetative anatomy.
Ovule morphology, etc., of Euphronia are still very poorly known.
Previous Relationships. Euphronia has been included in Trigoniaceae (Airy Shaw 1966; Hutchinson 1973; Takhtajan 1997) or Vochysiaceae (Cronquist 1981; Lleras 1978, Mabberley 1997). However, Euphronia and Trigoniaceae differ in a number of features, including those of wood anatomy (see Lleras 1976 for a table) and other aspects of vegetative anatomy (e.g. Trigoniaceae lack the mucilaginous hypodermis of Euphronia: Sajo & Rudall 2002) and are not sister taxa.
CHRYSOBALANACEAE R. Brown, nom. cons. —— Synonymy: Hirtellaceae Horaninow, Licaniaceae Martynov
- Back to Malpighiales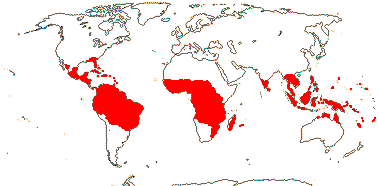
Trees or shrubs; trihydroxyflavonoids, distinctive unsaturated fatty acids in the seeds +, ellagic acid 0; trunk often with red exudate; (cork ± deep-seated); true tracheids +; wood siliceous [with SiO2 grains], parenchyma in apotracheal bands; nodes 5:5; petiole vasculature arcuate/annular (with medullary plates, etc.), wing bundles +; branching from previous flush; (foliar sclereids +); leaves often two-ranked, lamina vernation (flat-)conduplicate, abaxial surface often with flat glands, esp. near base, (margins toothed), (stipules petiolar or intrapetiolar); inflorescence various; (flowers almost polysymmetric); (C 0); A (2-)5-many, usually long-exserted, abaxial members only/best developed, filaments (connate), (coiled in bud), (staminodes adaxial); tapetal cells 2-nucleate; pollen (4-colporate), angled in polar view; usu. only abaxial carpel developed, (loculus divided), (all three carpels fertile), often borne on side or top of tube, style ± gynobasic, stigma punctate to 3-lobed; ovules (initiated when carpels are open), ± basal, erect, outer integument 5-12 cells across, inner integument 5-12 cells across, (micropyle not zig-zag); megaspore mother cells several, embryo sac lacking antipodals; fruit a 1-seeded drupe, medium-sized to large, (germination plug or lines), endocarp densely hairy (not); (seed ruminate), testa (multiplicative), vascularized, undistinguished or mesotestal, exotesta collapsed-fibrous, (tanniniferous), tegmen multiplicative; n = 10, 11, x = 11 (?10); germination cryptocotylar, hypogeal.
18 [list]/530: Licania (220), Hirtella (107), Couepia (70), Parinari (39). Pantropical, especially American. Map: from van Balgooy (1993), Prance and Sothers (2003a, b) and Trop. Afr. Fl. Pl. Ecol. Distr. vol. 2 (2006). [Photo - Flower.]
Age. The age of crown-group Chrysobalanaceae may be Palaeocene, around 59-58 Ma (Bardon et al. 2012), rather older, some (74.9-)66.2(-60.3) Ma (Xi et al. 2012b: Table S7, Atuna and the rest), or as young as (37.3-)33.4(-30.2) Ma (Bardon et al. 2016) or (43.9-)38.9(-34.2) Ma (Chave et al. 2020).
Evolution: Divergence & Distribution. For the fossil record of Chrysobalanaceae, see Jud et al. (2016).
Chrysobalanaceae are possibly Old World in origin, probably moving from the paleotropics to the Neotropics either by long distance dispersal or by the North Atlantic land bridge. Most of the diversity in the family is now in a Neotropical clade whose arrival there is dated to ca 29.5 Ma, mid Oligocene, although most diversification within this clade is Miocene and younger (Jud et al. 2016; Bardon et al. 2016; Dick & Pennington 2019; Chave et al. 2020; Hoorn et al. 2023); note, however, that the crown-group age of the Neotropical clade in Bardon et al. (2012) is (57-)47(-40) Ma. The net rate of diversification was overall higher in the Neotropics than the palaeotropics, despite the extinction rate also being higher in the former. The driver of all this is unclear, particularly since the family is found in lowland habitats and Africa has often been thought of as an area that is likely to have a higher extinction rate (Bardon et al. 2012). Chave et al. (2020, see also Hoorn et al. 2023) linked the Andean orogeny to diversification within the family, and again diversification was Miocene in age, although that of the very speciose Hirtella began only in the very latest Miocene.
Ecology & Physiology. Chrysobalanaceae are common in terms of both numbers of species and individuals with stems at least 10 cm d.b.h. in the Amazonian tree flora, but they do not have a disproportionally high number of common species (ter Steege et al. 2013).
Seed Dispersal. For seed dispersal, generally by animals, see Prance and Mori (1983).
Plant-Animal Interactions. Amazonian Hirtella species of section Myrmecophila form obligate associations with ants, Allomerus spp. (Rico-Gray & Oliveira 2007; Ruiz-González et al. 2011; Orivel et al. 2017: good summary of costs and benefits for the various partners). The ant cultivates an ascomycete fungus Trimmatostroma (Chaetothyriales - see Vasse et al. 2017), whose hyphae strengthen the carton that makes the walls of the galleries within which the ants lurk, waiting to ambush their prey (Dejean et al. 2005). However, the ant domatia drop from older leaves of H. myrmecophila, otherwise A. octoarticulata might well eat the inflorescences, effectively sterilizing the plant (Izzo & Vasconcelos 2002). Nitrogen moves from the fungus - ultimately the ant? - to the plant (Leroy et al. 2017).
Chemistry, Morphology, etc.. Chrysobalanaceae, rich in silica, contribute notably to the pool of phytoliths (Piperno 2006). Syllepsis is uncommon both here (Keller 1994), and probably more generally in the chrysobalanoids.
Cronquist (1981) suggested that the pollen grains might also be colpate.
For more information, see Prance (2013) and Prance and White (1988: nicely illustrated), both general, Hegnauer (1973, 1990: chemistry), Badami and Patil (1981: seed fatty acids), Morvillez (1918: petiole vasculature), Corrêa et al. (2018: leaf anatomy), Tobe and Raven (1984: embryology), Hill (1937: germination) and LaFrankie (2011: field characters).
Phylogeny. Prance (2020) summarizes much recent work on the family. A molecular study (Yakandawala et al. 2010) yielded support for the monophyly of the genera but poor resolution of deeper relationships, and suggested that groupings apparent in earlier morphological "taximetric" studies (Prance et al. 1969; Prance & White 1988) should be reexamined; the situation remained largely unchanged in the study by Bardon et al. (2012), with practically no support for relationships along the backbone of the tree. Licania may be wildly para/polyphyletic, and Hirtella and Coupeia, especially the latter, are also not monophyletic (Sothers et al. 2014; for problems with the first two genera, see also Bardon et al. 2016). There is some support for Atuna being sister to the rest of the family, perhaps in a clade with one or two other taxa including Kostermanthus (Bardon et al. 2012; see also Wurdack & Davis 2009), although basal relationships were unclear in Sothers et al. (2014: Atuna not included), Maranthes and Parinari perhaps being near basal. For relationships around Licania, see Sothers et al. (2016). Bardon et al. (2016) found the poorly-known Kostermanthus alone to be sister to the rest of the family, while the clade [Bafodeya + Kostermanthus] occupied this position in Chave et al. (2020).
Classification. Compare Prance (1989: New World Taxa), Prance and Sothers (2003a, b: world monograph) and Prance (2020) - from 12 to 24 genera in 60 years. Generic limits are beuing reworked as relationships become better understood (Sothers et al. 2014: Coupeia, 2016: Licania).
Previous Relationships. The flowers of some Chrysobalanaceae look rather like those of Prunus, and Chrysobalanaceae and Rosaceae were often considered to be close (e.g. Cronquist 1981; Takhtajan 1997), either recognized as separate families but placed more or less adjacent in the sequence, or Chrysobalanaceae might even be included as a subfamily of Rosaceae. However, there are numerous differences between them (see table in Prance 1972a).
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04985_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04125_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04685_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02421_001.jpg',600,500)){kind=link}
){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04731_001.jpg',600,500)){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02994_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02362_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04996_001.jpg',600,500)){kind=link}
![[Photo - Fruit]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-03966_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04833_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-03677_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02103_001.jpg',600,500)){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04480_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
![Fruit]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02387_001.jpg',600,500)){kind=link}
){kind=link}
![Flower, Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02012_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-03095_001.jpg',600,500)){kind=link}
){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04120_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02056_001.jpg',600,500)){kind=link}