20. BRYOERYTHROPHYLLUM Plates
24–
25.
Bryoerythrophyllum Chen, Hedwigia 80: 4, 1941. Type: Bryoerythrophyllum
recurvirostrum (Hedw.) Chen.
Globulina C. Müll., Nuov. Giorn. Bot. Ital., N.S. 4: 39, 1897, hom. illeg.
non Link in Nees 1820, nec Turpin 1827, nec Velen. 1834,
nec Spegaz. in Sacc. 1891. Type: Globulina boliviana C. Müll.
Erythrophyllum (Lindb. in Braithw.) Loeske, Hedwigia 47: 175, 1908, hom. illeg. non
J. Ag., 1872.
Erythrobarbula Steere, Bryologist 54: 199, 1951, nom illeg. incl. gen. prior.
Didymodon subg. Erythrophyllum Limpr., Laubm. Deutschl. 1: 405, 1887.
Type: Didymodon rubellus BSG.
Barbula subg. Erythrophyllum (Lindb. in Braithw.) C. Jens., Skand.
Bladmfl. 243, 1939.
Barbula subg. Erythrobarbula Szafr., Fl. Polska Mchy 1: 213, 1957
[1958], nom. inval. descr. polon.
Barbula sect. Erythrophyllum Lindb. in Braithw., Brit. Moss Fl. 1: 260,
1887. Type: Barbula rubella Mitt. in Lindb.
Didymodon sect. Orthocarpae Broth., Nat. Pfl. 1(3): 405, 1902.
Didymodon sect. Amblystegioideae Broth., Nat. Pfl. 1(3): 405, 1902.
Pl
ants turf-forming or loosely caespitose,
usually green above and red-brown below. Stems occasionally branching,
short or to several cm in length, rounded-pentagonal in transverse section, usually
with central strand, sclerodermis usually present, hyalodermis usually
absent; axillary hairs several cells in length, sometimes with 1–3 brownish
basal cells. Leaves appressed when dry, spreading when wet, ovate,
elliptical or lanceolate, 1–5 mm in length, often grooved along the costa,
marins usually recurved below or to near apex, seldom plane, entire or more
often dentate above, often with a border of 3–4 rows of thick-walled
cells; apex rounded-obtuse to acute; base usually ovate to oblongand
sheathing; costa ending a few cells below the apex to
short-excurrent, with quadrate, papillose cells ventrally, cells
elongate dorsally, in transverse section showing a differentiated epidermis
ventrally or on both sides, ca. 2–6 rows of cells across costa ventrally at
midleaf, two stereid bands present, guide cells 2–4 in 1(–2) layers, one or
more hydroid strands occasionally present; upper laminal cells subquadrate to
short-rectangular, ca. 10–15 µm in width, 1(–2):1, walls evenly thickened,
weakly sinuose, superficially nearly flat, occasionally bulging; papillae
solid, or occasionally hollow, bifid, or occasionally
flattened or compound, mostly 4–6 per lumen, obscuring the lumens;
basal cells usually hyaline and rectangular, filling most of the base
medially, little wider than upper cells, occasionally bulging,
2–5:1. Asexual reproduction occasional, by unicellular propagula in
masses in the leaf axils or obovoid brood bodies borne on rhizoids in
the soil. Dioicous or occasionally monoicous. Perichaetia and perigonia
terminal. Perichaetial leaves usually sheathing in the lower half, larger than
the stem leaves, long-oval to long-lanceolate, lower cells long rhomboidal in
lower half. Perigoniate plants occasionally small, bud-like. Seta elongate,
twisted clockwise below, often counterclockwise above, usually 1 per perichaetium;
theca ellipsoidal to cylindric, usually red-brown, 0.7–2.5(–4.0) mm in length,
sometimes curved; stomates phaneropore, at base of theca, annulus of 1–2
rows of vesiculose cells, often revoluble or deciduous in pieces; operculum
short-conic to short-rostrate, cells in straight or oblique rows; peristome
none, rudimentary, or consisting of 16 or 32 linear rami,
erect to twisted to 2–4 times counterclockwise, usually densely spiculose,
basal membrane low or absent. Operculum short-conic to short-rostrate, 0.2–1.2
in length. Calyptra cucullate, smooth, ca. 2 mm in length. Spores ca. 8–15 µm
in diameter, light brown, lightly papillose. Laminal KOH color reaction red
to orange-red. Reported chromosome number n = 13, 13+m, 14, 26+2m.
An
essentially cosmopolitan genus. More extensive treatments of this taxon for the
New World have been done by Zander (1978g, 1980a, 1981a).
Bryoerythrophyllum has much the appearance of Barbula or Didymodon
species, but is easily distinguished by a combination of features, most of
which are usually present: red coloration (at least in older parts of stems—Bryoerythrophyllum
fuscinervium, for instance, generally having immature leaves at the stem
apex with a yellow KOH reaction), the red color usually evident even without
KOH; bifid, crowded laminal papillae obscuring the upper laminal cell lumens;
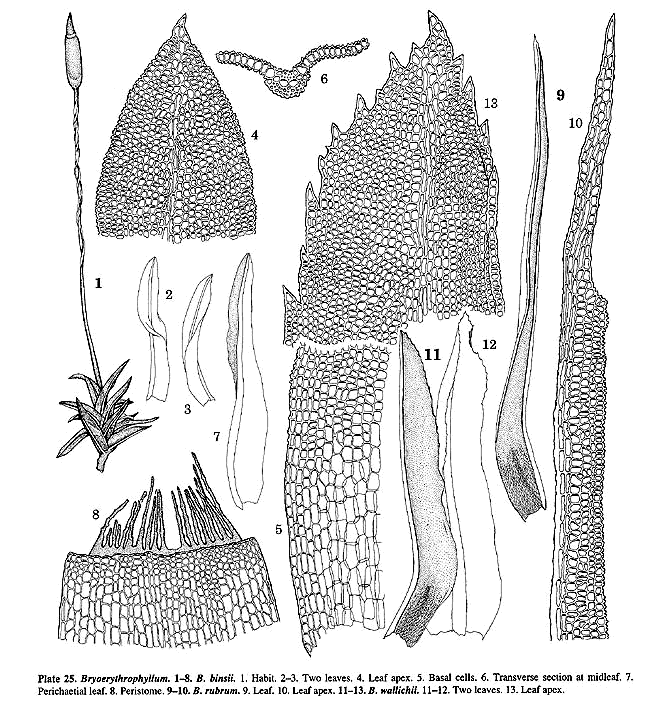
usually flat superficial laminal cell walls (easily seen in section, Pl. 24, f.
7; 25, f. 6); and well-differentiated leaf basal cells (Pl. 24, f. 6; 25, f.
5). Like Barbula and Didymodon, Bryoerythrophyllum
exhibits a full range of peristome development, some species eperistomate (e.g.
B. boliviana, Pl. 24, f. 11), some with rudimentary peristomes
(e.g. B. rotundifolia), others with straight, well-developed teeth (e.g.
B. binsii, Pl. 25, f. 8, and B. recurvirostrum), and finally
those with long, twisted peristomes (e.g. B. inaequalifolium, Pl. 24, f.
10). Only recently has it been recognized (Saito 1975a; Zander 1978g, 1980a)
that this genus is rather large and includes many species previously placed in Barbula
and Didymodon. Bryoerythrophyllum fuscinervium and the similar B.
columbianum have a rather distinctive broad and bulging ventral costal
epidermis of a single layer of cells and may be confused with Pseudocrossidium
species (such as P. chilense) but the red KOH reaction and distinct
ventral stereid band are diagnostic. Bryoerythrophyllum differs from Syntrichia,
which is also KOH red, in having two stereid bands, a usually narrower leaf
shape and dorsal costal epidermis commonly differentiated. The upper areolation
of rather large cells with evenly thickened walls is similar to that of Trichostomum
subg. Oxystegus but the red color in KOH, presence of a stem central
strand and costal hydroid strand(s), and the basal cells extending farthest up
the leaf medially distinguish Bryoerythrophyllum (e.g. cf. B.
chimborazense). The genus Mironia is very similar, but differs in
the keeled leaves with bistratose upper margins (the upper lamina of Bryoerythrophyllum,
in one species, B. sharpii, may be bistratose throughout) and, in two
species of Mironia, the presence of propaguloid leaf apices.
The
genus Globulina was published validly by C. Müller (1897a) in a combined
genus and species description. Although Müller indicated that Seligeria
globifera Hampe (║ Globulinella globifera (Hampe) Steere) was
related to and was probably also a member of the genus (“Globulina
mihi”), he left the genus monotypic with G. boliviana. Later, Müller
(1901) established Seligeria subg. Globulina C. Müll., with S.
globifera the only species, being thus the type of that, different,
taxon. Steere (Steere & Chapman 1946) proposed Globulinella as a new
genus, with Globulina C. Müll. non Link in Nees as a synonym, but
excluded the type of Globulina, G. boliviana, later in the
publication. Steere apparently did not intend Globulinella as a nom.
nov. for Globulina as was suggested by Magill (1977a). This
confusion is reflected in the Index Nominum Genericorum (Farr et al. 1979).
Actually, Globulina C. Müll., hom. illeg., is not a synonym of Globulinella
Steere, but is instead a synonym of Bryoerythrophyllum Chen. Seligeria
subg. Globulina C. Müll. is a synonym of the valid genus Globulinella
Steere, both taxa having the same type species, G. globifera (lectotype fide
van der Wijk et al., 1959–69).
The
type of Husnotiella glossophylla Herz. is not at L, JE or W and may have
been destroyed during the Second World War (pers. comm. F. K. Meyer at JE).
Specimens collected by Hosseus in Argentina (FH, W) identified by Bartram as this
or as H. baueri are eperistomate collections of the rather
variable Didymodon tophaceus. The description and illustrations of H.
glossophylla itself indicate that it is probably the same as Bryoerythrophyllum
calcareum.
Additional
literature: Chen (1941), Crum (1957a), Long (1982a, 1982b), Ratkowsky (1980).
Number
of accepted species: 27.
Species
examined: B. andersonianum (BUF, TENN), B. binnsii
(BUF), B. bolivianum (BUF, JE, TENN), B. byrdii (Bartr.)
Zand. (NY), B. calcareum (BUF, FH, MICH, TENN, US), B. campylocarpum,
B. chimborazense (NY), B. columbianum (UBC, US), B.
ferruginascens, B. fuscinervium (BUF, NY), B. hostile
(JE), B. inaequalifolium (BUF, DUKE, NY, TENN), B. jamesonii,
B. ligulare (NY), B. recurvirostrum, B. rotundatum
(H), B. rubrum (BUF, NY), B. sharpii (BUF), B.
wallichii (NY, US).
New
heterotypic synonymy: Bryoerythrophyllum columbianum var. atacamense
Zand. & Lewis in Lewis = Bryoerythrophyllum fuscinervium (Mitt.)
Zand. Didymodon integrifolius Broth. in Mildbr. var. paucidentatus
Thér. = Bryoerythrophyllum campylocarpum (C. Müll.) Crum. Didymodon
luzonensis Bartr. = Bryoerythrophyllum ferruginascens (Stirt.) Giac.
(the type of the former at FH is of fruiting material and includes the
characteristic propagula). Didymodon merceyoides Broth. in Herz. = Bryoerythrophyllum
campylocarpum (C. Müll.) Crum. Didymodon pelichucensis Williams = Bryoerythrophyllum
campylocarpum (C. Müll.) Crum. Hyophila calymperoides Thér. &
Nav. = Bryoerythrophyllum campylocarpum (C. Müll.) Crum.
New
combinations: Bryoerythrophyllum byrdii (Bartr.) Zand., comb. nov.
(Barbula byrdii Bartr., Ann. Missouri Bot. Gard. 25: 720, 1938). Bryoerythrophyllum
chimborazense (Mitt.) Zand., comb. nov. (Tortula chimborazensis
Mitt., J. Linn. Soc. Bot. 12: 163, 1869; Didymodon chimborazensis
(Mitt.) Broth. in Par.). Bryoerythrophyllum fuscinervium (Mitt.) Zand., comb.
nov. (Tortula fuscinervia Mitt., J. Linn. Soc. Bot. 12: 154, 1869; Barbula
fuscinervia (Mitt.) Jaeg. Bryoerythrophyllum ligulare (Mitt.) Zand.,
comb. nov. (Tortula ligularis Mitt., J. Linn. Soc. Bot. 12: 156,
1869; Barbula ligularis (Mitt.) Jaeg.).
{kind=link}
{kind=link}