44. WEISSIA Plates
59,
60,
61–
62.
Weissia Hedw., Sp. Musc.
168, 1801. Lectotype: Weissia controversa Hedw. fide Mitten, Kew J.
Bot. 8: 258, 1856.
Cavanillea Borkh., Disp.
Pl. 251, 1809, hom. illeg. non Medikus, 1787, non Desf. in Lam.,
1792.
Subg. Weissia
Simophyllum Lindb., Act.
Soc. Sci. Fenn. 10: 74, 1871, nom. illeg. incl. gen. prior.
Rechingerella Fröhl.,
Ann. Naturhist. Mus. Wien 66: 36, 1962 [1963], hom. illeg. non Petrak.
Type: Rechingerella macedonica J. Fröhl.
Weissia subg. Weisia
BSG, Bryol. Eur. 1: 5, 1851 (fasc. 47. Consp. vol. 1: VII), nom. illeg.
Weissia subg. Microweisia
BSG, Bryol. Eur. 1: 5, 1851 (fasc. 46–47 Consp. vol. 1: VII).
Weissia subg. Euweisia
Schimp., Syn. 54, 1860, nom. illeg.
Hymenostomum subg. Weisia
(Hedw.) Andrews, Bryologist 23: 31, 1920, nom. illeg. prior. ut gen.
Tortella subg. Nanotortella
C. Jens., Danm. Moss. 2: 318, 1923, nom. illeg. incl. typ. gen.
prior., p.p.
Bryum sect. Weissia
(Hedw.) Relh., Fl. Cantabr. 425, 1802. Type: Bryum virens Relh.
Weissia sect. Controversae
Nees & Hornsch., Bryol. Germ. 2(2): 25, 31, 1831.
Weissia sect. Viridulae
BSG, Bryol. Eur. 1: 66, 1846 (fasc. 33–36 Mon. 6).
Weissia sect. Euweisia
C. Müll., Syn. 1: 651, 1849, nom. illeg.
Didymodon sect. Pusilli
Kindb., Eur. N. Amer. Bryin. 2: 272, 1897. Type: Didymodon triumphans
(De Not.) Kindb.
Barbula sect. Edentella
C. Müll., Gen. Musc. Fr. 453, 1900. Type: Barbula schweinfurthiana C.
Müll.
Weissia subsect. Sphalerostomae
Nees & Hornsch., Bryol. Germ. 2(2): 24, 25, 1831, nom. illeg.
Subg. Astomum (Hampe) Kindb., Eur. N. Amer. Bryin. 2: 283, 1897.
Astomum Hampe, Flora 20:
285, 1837. Lectotype: Astomum crispum (Hedw.) Hampe fide
Margadant, Acta Bot. Neerl. 8: 274, 1959.
Sphaerangium Schimp.,
Syn. 12, 1860, nom. illeg. incl. gen. prior.
Systegium Schimp., Syn.
30, 1860. Lectotype: Systegium crispum (Hedw.) Schimp.
Astomum subg. Euastomum
(C. Müll.) Broth., Nat. Pfl. 1(3): 384, 1901, nom. illeg.
Hymenostomum subg. Astomum
(Hampe) Andrews, Bryologist 23: 31, 1920.
Tortella subg. Systegium
C. Jens., Danm. Moss. 2: 317, 1923, hom. illeg. Type: Tortella crispa
(Hedw.) C. Jens.
Phascum sect. Astomum
(Hampe) Ångstr. in Fries, Summ. Veg. Scand. 1: 97, 1846.
Weissia sect. Astomum
(C. Müll.) C. Müll., Bot. Zeit. 5: 98, 1847, as autonym.
Weissia sect. Systegium
Lindb., Öfv. K. Vet. Ak. Förh. 21: 230, 1864. Lectotype: Weissia crispa
(Hedw.) Mitt.
Acaulon sect. Macrobryum
C. Müll., Linnaea 43: 353, 1882. Type: Acaulon lorentzii C. Müll.
Phascum sect. Systegium
C. Müll., Hedwigia 37: 76, 1898, nom. illeg. incl. sect. prior.
Subg. Hymenostomum (R. Br.) Limpr., Laubm. Deutschl. 1: 225,
1886, as autonym.
Hymenostomum R. Br., Trans.
Linn. Soc. London 12(2): 572, 1818. Type: Hymenostomum microstomum
(Hedw.) R. Br.
Gymnostomum subg. Hymenostomum
(R. Br.) Schimp., Syn. 33, 1860.
Mollia subg. Hymenostomum
(R. Br.) Lindb., Musci Scand. 21, 1879.
Weissia subg. Hymenostomum
Limpr., Laubm. Deutschl. 1: 225, 1886, nom. illeg.
Hymenostomum subg. Kleioweisia
Limpr., Laubm. Deutschl. 1: 224, 1886. Type: Hymenostomum rostellatum
(Brid.) Schimp.
Weissia subg. Hymenostomum
(R. Br.) Kindb., Eur. N. Amer. Bryin. 2: 283, 1897, nom. illeg.
Hymenostomum subg. Euhymenostomum
Andrews, Bryologist 23: 31, 1920, nom. illeg.
Weissia sect. Hymenostomum
(R. Br.) BSG, 1: 56, 1846 (fasc. 33–36 Mon. 6), as autonym.
Hymenostomum sect. Microstoma
B.&S. in BSG, Bryol. Eur. 1: 56, 1846 (fasc. 33–36 Mon. 6), nom. illeg.
incl. typ. gen. Type: Hymenostomum microstomum (Hedw.) R. Br.
Hymenostomum sect. Tortilia
B.&S. in BSG, Bryol. Eur. 1: 56, 1846 (fasc. 33–36 Mon. 6). Type: Hymenostomum
tortile (Schwaegr.) B.&S.
Weissia sect. Hymenostomum
(R. Br.) C. Müll., Syn. Musc. 1: 660, 1849.
Mollia sect. Hymenostomum
(R. Br.) Braithw., Brit. Moss Fl. 1: 230, 1885.
Subg. Phasconica (C. Müll.) Zand. see below.
Phasconica C. Müll.,
Linnaea 43: 438, 1882. Type: Phasconica lorentzii C. Müll., lectotyp.
nov.
Subg. Pseudopottia Kindb., Eur. N. Amer. Bryin. 2: 283, 284,
1897.
Sect. Gymnostomum Mitt., J. Linn. Soc. Bot. 12: 41, 129, 131,
1869.
Sect. Tortularia Mitt., J. Linn. Soc. Bot. 12: 15, 130, 139,
1869.
Subsect. Hapalostomae Nees & Hornsch., Bryol. Germ. 2(2):
24, 25, 1831.
Plants in low cushions
or turfs or loosely caespitose, green above, brown to tan or yellow below.
Stems branching irregularly, to 1.0 cm in length, transverse section rounded-pentagonal,
central strand present, occasionally hollow or very thick, sclerodermis
weakly differentiated in 1–2 layers, hyalodermis weakly differentiated to
distinct, seldom absent; axillary hairs to 10 cells in length, basal 1–2 cells
thicker walled; sparsely radiculose. Leaves incurved, tubulose, often
contorted or spiralled when dry, spreading and occasionally sharply reflexed
above a sheathing base when moist, long-ligulate, oblong or
triangular to lanceolate, 1.5–2.5(–4.0) mm in length, upper lamina broadly
channeled across leaf, seldom narrowly channeled along costa, margins
sharply incurved (or occasionally tightly involute) above the leaf base,
seldom merely erect-incurved near apex, entire, occasionally fragile and
breaking off in large rectangles; apex sharply acute to subulate, occasionally
broadly acute, obtuse, or weakly cucullate; base scarcely differentiated to
ovate or rectangular, occasionally half-sheathing; costa shortly and sharply
mucronate, seldom subpercurrent, superficial cells quadrate or
occasionally short-rectangular to elongate ventrally, elongate dorsally,
4–8(–10) rows of cells across costa ventrally at midleaf, costal transverse
section ovate, occasionally circular or semicircular, two stereid bands
present, usually of about equal size, differentiated epidermis
present ventrally, absent or occasionally weakly differentiated dorsally, guide
cells 4–6(–8) in 1 layer, hydroid strand absent or occasionally present;
upper laminal cells subquadrate to hexagonal, 7–13 µm in width, 1:1, walls
thin to evenly thickened, superficially strongly bulging on both exposed
surfaces or more strongly protuberant on ventral surface; papillae bifid,
2–6 per lumen, occasionally fused into a capitulate papilla covering the lumen,
occasionally spiculose-branching, seldom absent; basal cells differentiated
across leaf, occasionally rising higher along margins in a vee, rectangular,
occasionally rhomboidal, 2–5:1, walls thin to evenly thickened. Rhizoid-borne
tuber-like propagula reported for W. controversa. Monoicous, occasionally
dioicous. Perichaetia terminal, inner leaves little different from
cauline leaves or somewhat larger, occasionally weakly sheathing the seta,
lower cells long-rhomboidal to midleaf. Perigonia appearing as stalked lateral
buds on perichaetiate plants (but variably present) or terminal on usually
smaller perigoniate plants. Seta 0.1–1.3 cm in length, 1(–2) per perichaetium,
yellowish brown, twisted clockwise; theca 1.0–2.2 mm in length, occasionally
inclined, yellowish to reddish brown, elliptical to ovate or cylindrical,
occasionally nearly spherical or urceolate, exothecial cells rectangular, walls
thin or becoming rather thick, rarely superficially bulging, stomates
phaneropore at base of capsule, annulus of 2–6 rows of persistent, vesiculose
cells, often on a differentiated collar, or annulus not differentiated; cleistocarpous
or stegocarpous, peristome teeth 16, short or rudimentary or
absent, occasionally lifts with operculum, oblong-truncate to
long-triangular, often irregularly cleft into two rami or variously
perforate, papillose to spiculose or spirally ridged, to 125(–250) µm, with up
to 10 articulations, straight or twisted weakly counterclockwise, basal
membrane absent or low, low-spiculose, mouth of capsule occasionally closed
by a hymenium, this internal to the peristome (when peristome present),
seldom macrostomous. Operculum conic to rostrate, 0.3–0.8(–1.5) mm in length,
cells straight or rarely somewhat twisted counterclockwise.
Calyptra cucullate (to short-triangular on cleistocarpous capsules),
smooth, 0.4–2.5 mm in length. Spores 14–28 µm in diameter, brown to yellowish
brown, low-papillose or occasionally highly papillose. Laminal KOH color
reaction yellow, usually pale yellow. Reported chromosome number n =
13, 13+m, 13+0–4m, 14, 26 (subg. Weissia); 13, 13+m, 26 (subg. Astomum);
8, 13, 14, 26 (subg. Hymenostomum).
A large genus found on
all continents except Antarctica, mostly growing on soil.
Weissia is superficially not well distinguished from Trichostomum.
Andrews (1945) felt that the Weissia complex should be “extended to
cover Trichostomum.” In Weissia, the central strand nearly
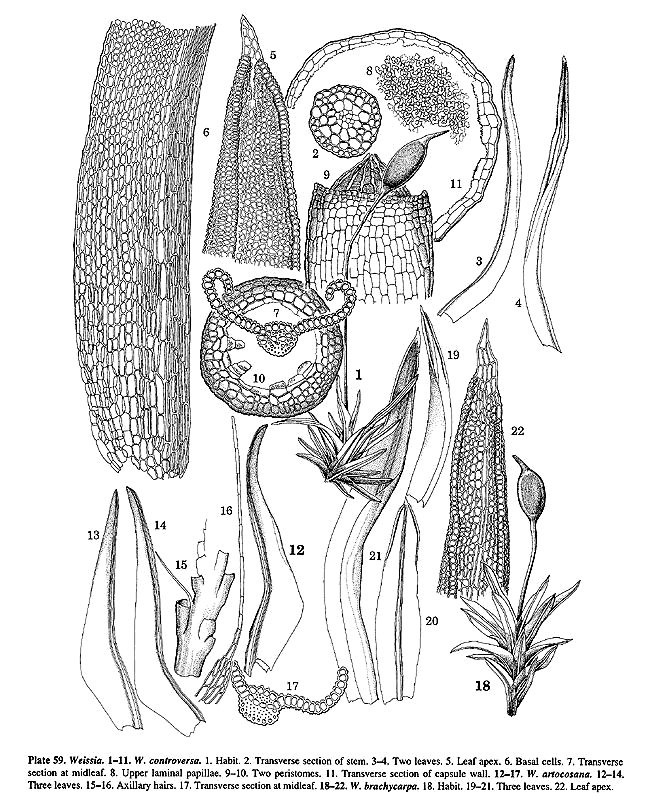
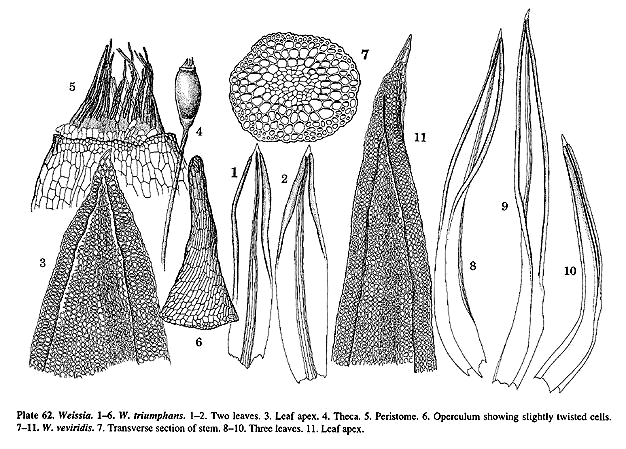
universally present (Pl. 59, f. 2; 62, f. 7) at least in the specimens
examined), upper laminal margins are usually narrowly and sharply incurved (Pl.
59, f. 7; 60, f. 5) as is the case with Chionoloma and Weissiodicranum
(those of Weissia termitidarum and certain others are loosely involute),
the upper laminal cells are often more strongly bulging ventrally than dorsally
(although usually overlain with papillae), the hydroid strand is usually absent
(but present in W. controversa), the sexual condition is commonly
monoicous (position of antheridia often varying between or within collections
of a particular species), perichaetial leaves are often larger than the
cauline, peristomes are generally short with the teeth more or less flattened
(Pl. 59, f. 9–10) or absent, and spores are rather large, 14–28 µm in diameter.
When present, the ventrally colliculate condition, i.e. bulging upper laminal
cells associated with slightly thicker, flattened dorsal superficial walls, has
not been emphasized by previous authors, but is common among the species
examined, being present to various degrees in, e.g. W. artocosana (Pl.
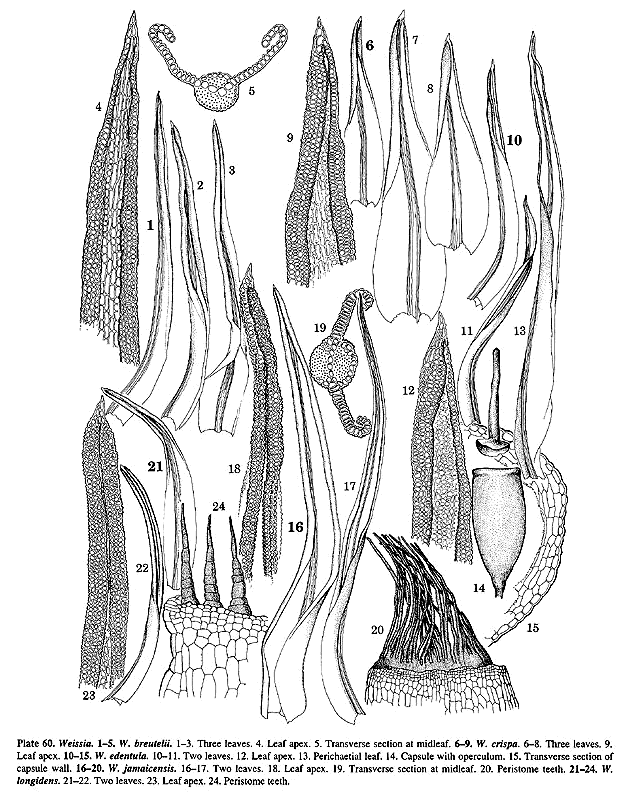
59, f. 17), W. breutelii (Pl. 60, f. 5), W. canaliculata, W.
condensa, W. controversa (Pl. 59, f. 7), W. ghatensis, W.
glaziouii, W. latiuscula, W. ligulaefolia, W. macrospora,
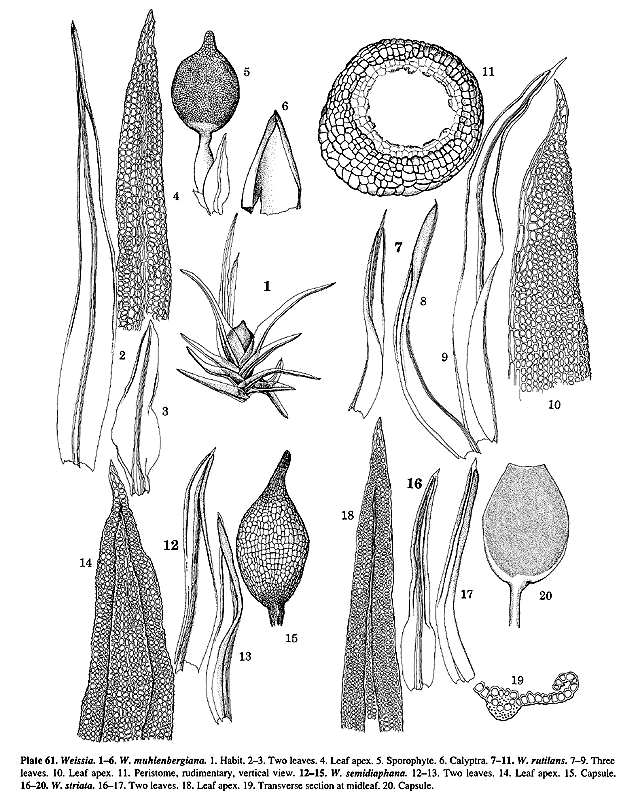
W. occidentalis, W. striata (Pl. 61, f. 19), W. termitidarum,
W. triumphans, W. veviridis and W. wimmeriana.
The best-developed
species of Weissia, W. jamaicensis (Pl. 60, f. 16–20), includes a
potpourri of features characteristic of other genera. It has the basal vee of
hyaline cells of Tortella, the much-enlarged ventral stereid band and
dilated leaf base of Pseudosymblepharis, and a peristome of 16 straight,
linear-lanceolate teeth cleft to a basal membrane into two spiculose rami quite
like that of Trichostomum crispulum (which, itself, occasionally
develops a basal vee and margins incurved near the apex, but lacks the swollen
ventral stereid band). In this case, the species are distinctive, but the
generic definitions break down. Trichostomum crispulum and T. perligulatum
have leaf margins that are often incurved near the apex but generally have
plane or broadly incurved margins throughout most of the leaf. Some species,
such as W. crispa (Pl. 60, f. 6–9) and the doubtfully distinct W.
muhlenbergiana (Pl. 61, f. 1–6, see Crum & Anderson's 1981 discussion),
have perichaetial leaves with plane to erect-incurved margins but cauline
leaves (and those on sterile stems) with weakly but narrowly incurved upper
margins, and are perhaps ultimately better assigned to Trichostomum in
the T. crispulum relationship. Weissia rutilans (Pl. 61, f. 7–11)
has upper laminal margins not easily assigned to either plane or narrowly
incurved status. In any case, to provide an interim classification that better
reflects perceived relationships, all Weissia taxa with clearly plane
upper cauline leaf margins examined in the course of this study are here
transferred to Trichostomum.
The fact that
peristome teeth may occasionally be represented in some species only by very
much reduced, truncate and colorless fragments, these often hidden below the
rim of the capsule mouth, indicates that past generic level distinctions
between peristomate and gymnostomous species of Weissia with similar
leaves may not be worthwhile. There is some correlation between thick capsule
walls and lack of a peristome (reflected in distinctions between Weissia
taxa of my treatments in the Moss Flora of Mexico, in press), but this must be
investigated in greater detail. Stoneburner (1985) demonstrated significant
variation in peristome expression in species of Weissia from the
southwestern United States, and even found rudimentary teeth in a species (W.
ligulaefolia) hitherto considered gymnostomous. She further discussed the
“evanescent membrane” of hymenostomous species as a poor taxonomic character,
explaining it as the upper surface of a somewhat persistent columella in both
eperistomate and peristomate taxa. The present study confirms this in that a
“hymenium” can often be seen below the peristome teeth of recently dehisced
capsules of, for instance, W. controversa. This is an old problem.
Referring to Brown's (1818) description of the then new genus Hymenostomum,
Greville and Arnott (1824b) wrote: “In Dr. Hooker's excellent figure of Gymnostomum
xanthocarpum in the Musci Exotici, the membrane which closes the sporular
bag is admirably delineated.¼ It ought to be observed, that Palisot has described this membrane in
the Mosses without a peristome, but denied it to the others” and (1824a) again
asserted: “Now, every moss possesses this ‘membranula indivisa,' arising
from the lining of the theca; but in all the species of Gymnostomum, it
is peculiarly evident at some particular stage.” However, Greville and Arnott
(1824b) admitted that: “The membrane to which we have already alluded, as
closing the mouth of the sporular bag, there is every reason to suppose, is
stronger and more durable in all such as have a naked peristome.”
A further taxonomic
complication is that the peristome teeth of occasional specimens, e.g. of W.
triumphans, are somewhat twisted counterclockwise, possibly an indication
of a relationship of these species to Tortella.
Regarding the synonymy
of Weissia and Astomum, a review of the problem and new evidence
(existence of sporophytes of intermediate morphology) was offered by
Stoneburner (1985). Many combinations at the genus level in Astomum and Hymenostomum
remain as “correct” names in the list of species and infraspecies given at the
end of this treatment. A mass transfer of these names to Weissia was
considered but rejected because many names, on examination of authentic material,
will prove to be more appropriately placed in Trichostomum (as emended
here). Recognition here of subgenera of Weissia that are based largely
on sporophyte morphology reflects recent studies by other authors (excepting
subg. Phasconica). Quite probably, intraspecific lines of evolution
involving sporophytic features of species with essentially identical
gametophytes will be found that cut across presently recognized subgeneric
limits. Unusual characters of sporophytes, such as the thick thecal walls of
species of subg. Hymenostomum or the strongly bulging exothecial cells
of W. macrospora, however, should also be evaluated and taken into
account.
A monotypic genus now
in the Viridivelleraceae, Viridivellus Stone, may actually represent
reduction of the Weissia gametophyte to an extreme, leaving only the
stem and perichaetial leaves on a persistent protonema; Stone (1980b) indicated
that this genus may be better placed in the Pottiaceae. It should be examined
during future revisions of the genus Weissia.
Brotherus (1924–25) placed Phasconica
between Tetrapterum and Hymenostomum, among genera he felt
intermediate between Astomum and Weissia. Hilpert (1933), who saw
material of the generitype, Phasconia lorentzii, noted the
closely inrolled upper laminal margins and suggested a relationship with Astomum.
Crundwell and Nyholm (1972a, 1974) included Phasconica in Weissia
subg. Astomum. Stone (1980b) carefully described and illustrated what is
surely Phasconica balansae from Australia, showing the strongly incurved
upper laminal margins characteristic of Weissia; however, she indicated
a reluctance to follow Crundwell and Nyholm (1972a, 1974) in assigning the
genus to Weissia subg. Astomum because of its macrostomous
capsule with dehiscent flattened operculum, stout columella, and ventral
surface of laminal cells distinctly mamillose and strongly papillose, among
other characters. Magill (1981) suggested that because of similarities of
gametophyte and dissimilarities of sporophyte (ex descr.) this genus “is
reminiscent of the relationship between Weissia and the segregate genera
Astomum Hampe and Hymenostomum R. Br.” Saito (1975, p. 417)
reduced Phasconica to synonymy with Weissia, with no further
explanation. Material of Phasconica C. Müll. s. str., seen during
this study was found mixed in the type of Trachycarpidium verrucosum (P.
tisserantii, PRE!, is referred to Trichostomum as a synonym of T.
unguiculatum see Crundwell & Nyholm 1974), and the genus Phasconica
is here given a new status as a subgenus of Weissia differing from other
subgenera mainly by the macrostomous, eperistomate capsule with a short seta
and a well differentiated, dehiscent operculum. Because of the unusual
sporophyte morphology, subg. Phasconia may not be part of a reduction
series in Weissia, but may instead be a remnant of a more primitive and
now largely extinct lineage; further analysis should be done at the species
level. Weissia platystegia (illustrated and discussed as Astomum
platystegium by Eddy (1990) and Norris and Koponen (1989) belongs in subg. Phasconica.
The singular Pseudosymblepharis
socotrana, including the synonym Barbula (sect. Edentella) schweinfurthiana
(isotype at BM) is transferred to Weissia (as W. artocosana, a nom.
nov.) on account of the weakly papillose, ventrally bulging upper laminal
cells, ventral stereid band smaller than the dorsal (Pl. 59, f. 17), and the
lack of a peristome, all characters found in Weissia. The strongly
sheathing leaf bases are found in both genera, and are variably demonstrated
among species of Pseudosymblepharis. This species is gametophytically
rather©
similar to Rhamphidium but differs in the entire leaves and lack of a
hydroid strand, as well as absence of a peristome.
Hybrids have not only
been reported between species of different subgenera of Weissia, but
also between Weissia crispa and Tortella flavovirens (Nicholson
1910). The sporophytes in this last case apparently had short setae; the thecae
were ovoid, partially cleistocarpous, rudimentarily peristomate, and covered by
very large calyptrae (characteristic of Tortella flavovirens). The
spores were also reported as remaining adherent in tetrads.
Additional literature:
Anderson and Lemmon (1972, 1973, 1974), Andrews (1920, 1922a, 1924, 1933), Bryan
(1956), Crundwell (1971b), Dietert (1979), Eckel (1986b), Flowers (1973b),
Grout (1900), Khanna (1960, 1964), Lemmon (1968), Lewinski (1983), Mizushima
(1957), Nicholson (1905a, 1906), Reese (1988, 1991), Reese and Lemmon (1965),
Robinson (1966), Shaw (1987a), Stoneburner (1981, 1986), Stoneburner and Wyatt
(1985), Williams (1966b), Zander (1985a), Zuttere et al. (1984).
Number of accepted
species: 97, plus 11 in Astomum and 16 in Hymenostomum.
Species examined: W.
abbreviata (NY), W. andersoniana (BUF, TENN), W.
argentinica (NY), W. artocosana (BM, NY), W. balansae
(PC), W. balansaeana (MO), W. bizotii (PC), W. brachycarpa
(BUF, NY), W. breutelii (BUF, NY), W. canaliculata (NY), W.
condensa (BUF, TENN, NY), W. controversa, W. crispa
(BUF, DUKE), W. diffidentia (US), W. edentula (MICH, NY), W.
fallax (NY), W. felipponei (PC), W. ghatensis (BM),
W. glaziouii (NY), W. jamesonii (TENN), W. jamaicensis
(BUF, NY, TENN), W. latiuscula (BM), W. ligulaefolia
(BUF, FH, NY), W. lineaefolia (NY), W. longidens (DUKE,
NY), W. longifolia (NY), W. ludoviciana (DUKE), W. macrospora
(NY), W. muhlenbergianum (DUKE), W. newcomeri (FH),
W. occidentalis (COLO), W. opaca (NY), W. ovalis
(NY), W. platystegia (BM), W. riograndensis (H), W.
rutilans (BUF, NY), W. semidiaphana (BUF, FH), W. sharpii
(BUF, DUKE), W. subacaulis (NY), W. submicacea (NY), W.
termitidarum (NY), W. triumphans (NY), W. unguiculata
(NY), W. veviridis (H), W. wimmeriana (NY).
New heterotypic
synonymy: Barbula (sect. Edentella) schweinfurthiana C.
Müll. = Weissia artocosana (Mitt.) Zand. Trichostomum bermudanum
(Mitt.) Par. = Weissia jamaicensis (Mitt.) Grout. Trichostomum
ekmanii Thér. = Weissia jamaicensis (Mitt.) Grout. Trichostomum
linealifolium C. Müll. = Weissia jamaicensis (Mitt.) Grout.
New names, statuses and combinations:
Weissia subg. Phasconica
(C. Müll.) Zand., comb. et stat. nov. (Phasconica C. Müll.,
Linnaea 43: 438, 1882).
Weissia abbreviata
(Thwait. & Mitt.) Zand., comb. nov. (Systegium abbreviatum
Thwait. & Mitt., J. Linn. Soc. Bot. 13: 299, 1873; Astomum abbreviatum
(Thwait. & Mitt.) Fleisch.).
Weissia artocosana
(Mitt.) Zand., nom. nov. (Symblepharis socotrana Mitt., Trans. R.
Soc. Edinburgh 31: 331, 1888; Pseudosymblepharis socotrana (Mitt.)
Thér.).
Weissia balansae (C.
Müll.) Zand., comb. nov. (Phasconica balansae C. Müll., Linnaea
43: 438, 1882).
Weissia (subg. Phasconica)
bizotii Zand., nom. nov. (Kleioweisiopsis involuta Biz.,
Rev. Bryol. Lichénol. 40: 119, 1974 invalid, single element not cited, fide
Crosby et al., 1992). Lectotype: Herb. Bizot; paratype: Herb. Pocs.
Weissia diffidentia
Zand., nom. nov. (Phascum recurvirostum C. Müll., Flora
71: 5, 1888; Tetrapterum recurvirostrum (C. Müll.) Broth.).
Weissia glaziouii Zand.,
nom. nov. (Hymenostomum striatum Geh. & Hampe, Vid. Medd. Naturh.
For. Kjoebenh. ser. 4,1: 84, 1879).
Weissia (subg. Phasconica)
lorentzii (C. Müll.) Zand., comb. nov. (Phasconica lorentzii
C. Müll., Linnaea 43: 438, 1882).
Weissia neocaledonica
(Thér.) Zand., comb. nov. (Aschisma neocaledonicum Thér., Diagn.
Esp. Var. Nouv. Mouss. 8: 4, 1910; Astomum neocaledonicum (Thér.)
Andrews).
Weissia riograndensis
(Broth.) Zand., comb. nov. (Hymenostomum riograndense Broth.,
Bih. K. Svensk. Vet. Ak. Handl. 26 Afd. 3(7): 19, 1900).
Weissia triumphans var. monspeliensis
(Schimp.) Zand., comb. nov. (Trichostomum monspeliense Schimp.,
Syn. ed. 2, 175, 1876; Trichostomum triumphans var. monspeliense
(Schimp.) Husn.).
Weissia veviridis Zand.,
nom. nov. (Trichostomum perviride Broth., Dansk. Bot. Ark. 2(9):
3, 1918).
{kind=link}
{kind=link}
{kind=link}
{kind=link}