EMBRYOPSIDA Pirani & Prado
Gametophyte dominant, independent, multicellular, initially ±globular, not motile, branched; showing gravitropism; glycolate oxidase +, glycolate metabolism in leaf peroxisomes [glyoxysomes], acquisition of phenylalanine lysase* [PAL], flavonoid synthesis*, microbial terpene synthase-like genes +, triterpenoids produced by CYP716 enzymes, CYP73 and phenylpropanoid metabolism [development of phenolic network], xyloglucans in primary cell wall, side chains charged; plant poikilohydrous [protoplasm dessication tolerant], ectohydrous [free water outside plant physiologically important]; thalloid, leafy, with single-celled apical meristem, tissues little differentiated, rhizoids +, unicellular; chloroplasts several per cell, pyrenoids 0; centrioles/centrosomes in vegetative cells 0, microtubules with γ-tubulin along their lengths [?here], interphase microtubules form hoop-like system; metaphase spindle anastral, predictive preprophase band + [with microtubules and F-actin; where new cell wall will form], phragmoplast + [cell wall deposition centrifugal, from around the anaphase spindle], plasmodesmata +; antheridia and archegonia +, jacketed*, surficial; blepharoplast +, centrioles develop de novo, bicentriole pair coaxial, separate at midpoint, centrioles rotate, associated with basal bodies of cilia, multilayered structure + [4 layers: L1, L4, tubules; L2, L3, short vertical lamellae] (0), spline + [tubules from L1 encircling spermatid], basal body 200-250 nm long, associated with amorphous electron-dense material, microtubules in basal end lacking symmetry, stellate array of filaments in transition zone extended, axonemal cap 0 [microtubules disorganized at apex of cilium]; male gametes [spermatozoids] with a left-handed coil, cilia 2, lateral, asymmetrical; oogamy; sporophyte +*, multicellular, growth 3-dimensional*, cuticle +*, plane of first cell division transverse [with respect to long axis of archegonium/embryo sac], sporangium and upper part of seta developing from epibasal cell [towards the archegonial neck, exoscopic], with at least transient apical cell [?level], initially surrounded by and dependent on gametophyte, placental transfer cells +, in both sporophyte and gametophyte, wall ingrowths develop early; suspensor/foot +, cells at foot tip somewhat haustorial; sporangium +, single, terminal, dehiscence longitudinal; meiosis sporic, monoplastidic, MTOC [= MicroTubule Organizing Centre] associated with plastid, sporocytes 4-lobed, cytokinesis simultaneous, preceding nuclear division, quadripolar microtubule system +; wall development both centripetal and centrifugal, 1000 spores/sporangium, sporopollenin in the spore wall* laid down in association with trilamellar layers [white-line centred lamellae; tripartite lamellae]; plastid transmission maternal; nuclear genome [1C] <1.4 pg, main telomere sequence motif TTTAGGG, KNOX1 and KNOX2 [duplication] and LEAFY genes present, ethylene involved in cell elongation; chloroplast genome with close association between trnLUAA and trnFGAA genes [precursors for starch synthesis], tufA, minD, minE genes moved to nucleus; mitochondrial trnS(gcu) and trnN(guu) genes +.
Many of the bolded characters in the characterization above are apomorphies of more or less inclusive clades of streptophytes along the lineage leading to the embryophytes, not apomorphies of crown-group embryophytes per se.
All groups below are crown groups, nearly all are extant. Characters mentioned are those of the immediate common ancestor of the group, [] contains explanatory material, () features common in clade, exact status unclear.
STOMATOPHYTES
Abscisic acid, L- and D-methionine distinguished metabolically; pro- and metaphase spindles acentric; class 1 KNOX genes expressed in sporangium alone; sporangium wall 4≤ cells across [≡ eusporangium], tapetum +, secreting sporopollenin, which obscures outer white-line centred lamellae, columella +, developing from endothecial cells; stomata +, on sporangium, anomocytic, cell lineage that produces them with symmetric divisions [perigenous]; underlying similarities in the development of conducting tissue and of rhizoids/root hairs; spores trilete; shoot meristem patterning gene families expressed; MIKC, MI*K*C* genes, post-transcriptional editing of chloroplast genes; gain of three group II mitochondrial introns, mitochondrial trnS(gcu) and trnN(guu) genes 0.
[Anthocerophyta + Polysporangiophyta]: gametophyte leafless; archegonia embedded/sunken [only neck protruding]; sporophyte long-lived, chlorophyllous; cell walls with xylans.
POLYSPORANGIOPHYTA†
Sporophyte well developed, branched, branching dichotomous, potentially indeterminate; hydroids +; stomata on stem; sporangia several, terminal; spore walls not multilamellate [?here].
TRACHEOPHYTA†
Vascular tissue + [tracheids, walls with bars of secondary thickening]; stomata numerous, involved in gas exchange.
II. TRACHEOPHYTA / VASCULAR PLANTS
Sporophyte long lived, cells polyplastidic, photosynthetic red light response, stomata open in response to blue light; plant homoiohydrous [water content of protoplasm relatively stable]; control of leaf hydration passive; plant endohydrous [physiologically important free water inside plant]; PIN[auxin efflux facilitators]-mediated polar auxin transport; (condensed or nonhydrolyzable tannins/proanthocyanidins +); borate cross-linked rhamnogalactan II, xyloglucans with side chains uncharged [?level], in secondary walls of vascular and mechanical tissue; lignins +; roots +, often ≤1 mm across, root hairs and root cap +; stem apex multicellular [several apical initials, no tunica], with cytohistochemical zonation, plasmodesmata formation based on cell lineage; vascular development acropetal, tracheids +, in both protoxylem and metaxylem, G- and S-types; sieve cells + [nucleus degenerating]; endodermis +; stomata numerous, involved in gas exchange; leaves +, vascularized, spirally arranged, blades with mean venation density ca 1.8 mm/mm2 [to 5 mm/mm2], all epidermal cells with chloroplasts; sporangia in strobili, sporangia adaxial, columella 0; tapetum glandular; sporophyte-gametophyte junction lacking dead gametophytic cells, mucilage, ?position of transfer cells; MTOCs not associated with plastids, basal body 350-550 nm long, stellate array in transition region initially joining microtubule triplets; archegonia embedded/sunken [only neck protruding]; suspensor +, shoot apex developing away from micropyle/archegonial neck [from hypobasal cell, endoscopic], root lateral with respect to the longitudinal axis of the embryo [plant homorhizic].
[MONILOPHYTA + LIGNOPHYTA]Sporophyte growth ± monopodial, branching spiral; roots endomycorrhizal [with Glomeromycota], lateral roots +, endogenous; G-type tracheids +, with scalariform-bordered pits; leaves with apical/marginal growth, venation development basipetal, growth determinate; sporangium dehiscence by a single longitudinal slit; cells polyplastidic, MTOCs diffuse, perinuclear, migratory; blepharoplasts +, paired, with electron-dense material, centrioles on periphery, male gametes multiciliate; nuclear genome [1C] 7.6-10 pg [mode]; chloroplast long single copy ca 30kb inversion [from psbM to ycf2]; mitochondrion with loss of 4 genes, absence of numerous group II introns; LITTLE ZIPPER proteins.
LIGNOPHYTA†
Sporophyte woody; stem branching axillary, buds exogenous; lateral root origin from the pericycle; cork cambium + [producing cork abaxially], vascular cambium bifacial [producing phloem abaxially and xylem adaxially].
SEED PLANTS† / SPERMATOPHYTA†
Growth of plant bipolar [plumule/stem and radicle/root independent, roots positively geotropic]; plants heterosporous; megasporangium surrounded by cupule [i.e. = unitegmic ovule, cupule = integument]; pollen lands on ovule; megaspore germination endosporic, female gametophyte initially retained on the plant, free-nuclear/syncytial to start with, walls then coming to surround the individual nuclei, process proceeding centripetally.
EXTANT SEED PLANTS
Plant evergreen; nicotinic acid metabolised to trigonelline, (cyanogenesis via tyrosine pathway); microbial terpene synthase-like genes 0; primary cell walls rich in xyloglucans and/or glucomannans, 25-30% pectin [Type I walls]; lignin chains started by monolignol dimerization [resinols common], particularly with guaiacyl and p-hydroxyphenyl [G + H] units [sinapyl units uncommon, no Maüle reaction]; roots often ≥1 mm across, stele diarch to pentarch, xylem and phloem originating on alternating radii, cork cambium deep seated, gravitropism response fast; stem apical meristem complex [with quiescent centre, etc.], plasmodesma density in SAM 1.6-6.2[mean]/μm2 [interface-specific plasmodesmatal network]; eustele +, protoxylem endarch, endodermis 0; wood homoxylous, tracheids and rays alone, tracheid/tracheid pits circular, bordered; mature sieve tube/cell lacking functioning nucleus, sieve tube plastids with starch grains; phloem fibres +; cork cambium superficial; leaf nodes 1:1, a single trace leaving the vascular sympodium; leaf vascular bundles amphicribral; guard cells the only epidermal cells with chloroplasts, stomatal pore with active opening in response to leaf hydration, control by abscisic acid, metabolic regulation of water use efficiency, etc.; branching by axillary buds, exogenous; prophylls two, lateral; leaves with petiole and lamina, development basipetal, lamina simple; sporangia borne on sporophylls; spores not dormant; microsporophylls aggregated in indeterminate cones/strobili; grains monosulcate, aperture in ana- position [distal], primexine + [involved in exine pattern formation with deposition of sporopollenin from tapetum there], exine and intine homogeneous, exine alveolar/honeycomb; ovules with parietal tissue [= crassinucellate], megaspore tetrad linear, functional megaspore single, chalazal, sporopollenin 0; gametophyte ± wholly dependent on sporophyte, development initially endosporic [apical cell 0, rhizoids 0, etc.]; male gametophyte with tube developing from distal end of grain, male gametes two, developing after pollination, with cell walls; embryo cellular ab initio, suspensor short-minute, embryonic axis straight [shoot and root at opposite ends], primary root/radicle produces taproot [= allorhizic], cotyledons 2; embryo ± dormant; chloroplast ycf2 gene in inverted repeat, trans splicing of five mitochondrial group II introns, rpl6 gene absent; ??whole nuclear genome duplication [ζ/zeta duplication event], 2C genome size (0.71-)1.99(-5.49) pg, two copies of LEAFY gene, PHY gene duplications [three - [BP [A/N + C/O]] - copies], 5.8S and 5S rDNA in separate clusters.
MAGNOLIOPHYTA
Lignans, O-methyl flavonols, dihydroflavonols, triterpenoid oleanane, apigenin and/or luteolin scattered, [cyanogenesis in ANA grade?], lignin also with syringyl units common [G + S lignin, positive Maüle reaction - syringyl:guaiacyl ratio more than 2-2.5:1], hemicelluloses as xyloglucans; root cap meristem closed (open); pith relatively inconspicuous, lateral roots initiated immediately to the side of [when diarch] or opposite xylem poles; epidermis probably originating from inner layer of root cap, trichoblasts [differentiated root hair-forming cells] 0, hypodermis suberised and with Casparian strip [= exodermis]; shoot apex with tunica-corpus construction, tunica 2-layered; starch grains simple; primary cell wall mostly with pectic polysaccharides, poor in mannans; tracheid:tracheid [end wall] plates with scalariform pitting, multiseriate rays +, wood parenchyma +; sieve tubes enucleate, sieve plates with pores (0.1-)0.5-≥10 µm across, cytoplasm with P-proteins, not occluding pores of plate, companion cell and sieve tube from same mother cell; ?phloem loading/sugar transport; nodes 1:?; dark reversal Pfr → Pr; protoplasm dessication tolerant [plant poikilohydric]; stomata randomly oriented, brachyparacytic [ends of subsidiary cells ± level with ends of guard cells], outer stomatal ledges producing vestibule, reduction in stomatal conductance with increasing CO2 concentration; lamina formed from the primordial leaf apex, margins toothed, development of venation acropetal, overall growth ± diffuse, secondary veins pinnate, fine venation hierarchical-reticulate, (1.7-)4.1(-5.7) mm/mm2, vein endings free; flowers perfect, pedicellate, ± haplomorphic, protogynous; parts free, numbers variable, development centripetal; P = T, C-like, each with a single trace, outer members not sharply differentiated from the others, not enclosing the floral bud; A many, filament not sharply distinguished from anther, stout, broad, with a single trace, anther introrse, tetrasporangiate, sporangia in two groups of two [dithecal], each theca dehiscing longitudinally by a common slit, ± embedded in the filament, walls with at least outer secondary parietal cells dividing, endothecium +, cells elongated at right angles to long axis of anther; tapetal cells binucleate; microspore mother cells in a block, microsporogenesis successive, walls developing by centripetal furrowing; pollen subspherical, tectum continuous or microperforate, ektexine columellate, endexine restricted to the apertural regions, thin, compact, intine in apertural areas thick, orbicules +, pollenkitt +; nectary 0; carpels present, superior, free, several, spiral, ascidiate [postgenital occlusion by secretion], stylulus at most short [shorter than ovary], hollow, cavity not lined by distinct epidermal layer, stigma ± decurrent, carinal, dry; suprastylar extragynoecial compitum +; ovules few [?1]/carpel, marginal, anatropous, bitegmic, micropyle endostomal, outer integument 2-3 cells across, often largely subdermal in origin, inner integument 2-3 cells across, often dermal in origin, parietal tissue 1-3 cells across, nucellar cap?; megasporocyte single, hypodermal, functional megaspore lacking cuticle; female gametophyte lacking chlorophyll, four-celled [one module, egg and polar nuclei sisters]; ovule not increasing in size between pollination and fertilization; pollen grains bicellular at dispersal, germinating in less than 3 hours, siphonogamy, pollen tube unbranched, growing towards the ovule, between cells, growth rate (ca 10-)80-20,000 µm h-1, tube apex of pectins, wall with callose, lumen with callose plugs, penetration of ovules via micropyle [porogamous], whole process takes ca 18 hours, distance to first ovule 1.1-2.1 mm; male gametophytes tricellular, gametes 2, lacking cell walls, ciliae 0, double fertilization +, ovules aborting unless fertilized; fruit indehiscent, P deciduous; mature seed much larger than fertilized ovule, small [<5 mm long], dry [no sarcotesta], exotestal; endosperm +, ?diploid [one polar nucleus + male gamete], cellular, development heteropolar [first division oblique, micropylar end initially with a single large cell, divisions uniseriate, chalazal cell smaller, divisions in several planes], copious, oily and/or proteinaceous, embryo short [<¼ length of seed]; plastid and mitochondrial transmission maternal; Arabidopsis-type telomeres [(TTTAGGG)n]; nuclear genome [2C] (0.57-)1.45(-3.71) [1 pg = 109 base pairs], ??whole nuclear genome duplication [ε/epsilon event]; ndhB gene 21 codons enlarged at the 5' end, single copy of LEAFY and RPB2 gene, knox genes extensively duplicated [A1-A4], AP1/FUL gene, palaeo AP3 and PI genes [paralogous B-class genes] +, with "DEAER" motif, SEP3/LOFSEP and three copies of the PHY gene, [PHYB [PHYA + PHYC]]; chloroplast IR expansions, chlB, -L, -N, trnP-GGG genes 0.
[NYMPHAEALES [AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]]: wood fibres +; axial parenchyma diffuse or diffuse-in-aggregates; pollen monosulcate [anasulcate], tectum reticulate-perforate [here?]; ?genome duplication; "DEAER" motif in AP3 and PI genes lost, gaps in these genes.
[AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]: phloem loading passive, via symplast, plasmodesmata numerous; vessel elements with scalariform perforation plates in primary xylem; essential oils in specialized cells [lamina and P ± pellucid-punctate]; tension wood + [reaction wood: with gelatinous fibres, G-fibres, on adaxial side of branch/stem junction]; anther wall with outer secondary parietal cell layer dividing; tectum reticulate; nucellar cap + [character lost where in eudicots?]; 12BP [4 amino acids] deletion in P1 gene.
[MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]] / MESANGIOSPERMAE: benzylisoquinoline alkaloids +; sesquiterpene synthase subfamily a [TPS-a] [?level], polyacetate derived anthraquinones + [?level]; outer epidermal walls of root elongation zone with cellulose fibrils oriented transverse to root axis; P more or less whorled, 3-merous [?here]; pollen tube growth intra-gynoecial; extragynoecial compitum 0; carpels plicate [?here]; embryo sac monosporic [spore chalazal], 8-celled, bipolar [Polygonum type], antipodal cells persisting; endosperm triploid.
[CERATOPHYLLALES + EUDICOTS]: ethereal oils 0 [or next node up]; fruit dry [very labile].
EUDICOTS: (Myricetin +), asarone 0 [unknown in some groups, + in some asterids]; root epidermis derived from root cap [?Buxaceae, etc.]; (vessel elements with simple perforation plates in primary xylem); nodes 3:3; stomata anomocytic; flowers (dimerous), cyclic; protandry common; K/outer P members with three traces, ("C" +, with a single trace); A ?, filaments fairly slender, anthers basifixed; microsporogenesis simultaneous, pollen tricolpate, apertures in pairs at six points of the young tetrad [Fischer's rule], cleavage centripetal, wall with endexine; G with complete postgenital fusion, stylulus/style solid [?here], short [<2 x length of ovary]; seed coat?; palaeotetraploidy event.
[PROTEALES [TROCHODENDRALES [BUXALES + CORE EUDICOTS]]]: (axial/receptacular nectary +).
[TROCHODENDRALES [BUXALES + CORE EUDICOTS]]: benzylisoquinoline alkaloids 0; euAP3 + TM6 genes [duplication of paleoAP3 gene: B class], mitochondrial rps2 gene lost.
[BUXALES + CORE EUDICOTS]: mitochondrial rps11 gene lost.
CORE EUDICOTS / GUNNERIDAE: (ellagic and gallic acids +); leaf margins serrate; compitum + [one position]; micropyle?; γ genome duplication [allopolyploidy, 4x x 2x], x = 3 x 7 = 21, 2C genome size (0.79-)1.05(-1.41) pg, PI-dB motif +; small deletion in the 18S ribosomal DNA common.
[ROSIDS ET AL. + ASTERIDS ET AL.] / PENTAPETALAE / [SANTALALES, CARYOPHYLLALES, SAXIFRAGALES, DILLENIALES, VITALES, ROSIDAE, [BERBERIDOPSIDALES + ASTERIDAE]: root apical meristem closed; (cyanogenesis also via [iso]leucine, valine and phenylalanine pathways); flowers rather stereotyped: 5-merous, parts whorled; P = K + C, K enclosing the flower in bud, with three or more traces, odd K adaxial, C with single trace; A = 2x K/C, in two whorls, alternating, (many, but then usually fasciculate and/or centrifugal); pollen tricolporate; G [(3, 4) 5], when 5 opposite K, whorled, placentation axile, style +, stigma not decurrent, compitum + [one position]; endosperm nuclear/coenocytic; fruit dry, dehiscent, loculicidal [when a capsule]; floral nectaries with CRABSCLAW expression, RNase-based gametophytic incompatibility system present.
Phylogeny. Prior to the seventh version of this site asterids were part of a major polytomy that included rosids, Berberidopsidales, Santalales, and Caryophyllales, but then the order of branching below the asterids seemed to be stabilizing, perhaps with a clade [Berberidopsidales [Santalales [Caryophyllales + Asterids]]] while rosid relationships seemed to be [Saxifragales [Vitales + Rosids]]]. However, recent work suggests a polytomy is indeed probably the best way to visualize relationships around here at present. So for further discussion of relationships at the base of asterids and rosids, see the Pentapetalae node.
[BERBERIDOPSIDALES + ASTERIDAE]: ?
ASTERIDAE / ASTERANAE Takhtajan: nicotinic acid metabolised to its arabinosides; (iridoids +); tension wood decidedly uncommon; C enclosing A and G in bud, (connate [sometimes evident only early in development, C then appearing to be free]); anthers dorsifixed?; if nectary +, gynoecial; G [2], style single, long; ovules unitegmic, integument thick [>5 cells across], endothelium +, nucellar epidermis does not persist; exotestal [!: even when a single integument] cells lignified, esp. on anticlinal and/or inner periclinal walls; endosperm cellular.
[CORNALES + ×ERICALES] / Ericornids: ?
×ERICALES Dumortier [= Cornales × euasterids/Gentianidae] - Main Tree.
Woody; nonhydrolysable tannins, triterpenoids incl. saponins +; vessel elements with simple perforation plates; nodes 1:1; leaves spiral, teeth with single vein and opaque deciduous cap; fruit capsular, K persistent; duplication of the PI gene. - 22 families, 346 genera, 12,005 species.
Includes Actinidiaceae, Balsaminaceae, Cyrillaceae, Clethraceae, Diapensiaceae, Ebenaceae, Ericaceae, Fouquieriaceae, Lecythidaceae, Marcgraviaceae, Mitrastemonaceae, Pentaphylacaceae, Polemoniaceae, Primulaceae, Roridulaceae, Sapotaceae, Sarraceniaceae, Sladeniaceae, Styracaceae, Symplocaceae, Tetrameristaceae, Theaceae.
Note: In all node characterizations, boldface denotes a possible apomorphy, (....) denotes a feature the exact status of which in the clade is uncertain, [....] includes explanatory material; other text lists features found pretty much throughout the clade. However, the precise node to which many characters, particularly the more cryptic ones, should be assigned is unclear. This is partly because homoplasy is very common, in addition, basic information for all too many characters is very incomplete, frequently coming from taxa well embedded in the clade of interest and so making the position of any putative apomorphy uncertain. Then there are the not-so-trivial issues of how character states are delimited and ancestral states are reconstructed (see above).
Age. Sytsma et al. (2006) proposed that diversification of Ericales began 109-103 Ma, Rose et al. (2018) suggest ca 110 Ma, Wikström et al. (2001: note topology) offered an age of (97-)92, 85(-80) Ma, Anderson et al. (2005) an age of 103-99 Ma, while Janssens et al. (2009) dated the crown group to 117±9.2 Ma and K. Bremer et al. (2004a) to 114 Ma (see also C. Zhang et al. 2020). D. Soltis et al. (2008: a variety of estimates) suggested an age of (126-)113(-85) Ma, Magallón and Castillo (2009) one of ca 98.85 Ma, Bell et al. (2010) an age of (102-)92 Ma, Shenk and Hufford (2010) an age of 100.7-87 Ma, Tank and Olmstead (pers. comm.) an age of (101.4-)96.2(-92.4) Ma, and Lemaire et al. (2011b) an age of (125-)118(-110) My; around 103.6 Ma is the age in Magallón et al. (2015), (117-)109(-100) Ma in Wikström et al. (2015), ca 114.1 Ma in C. Zhang et al. (2020) and (117.1-)113.7(-110.0) Ma (Bansal et al. 2021).
The fossil Archaeamphora was assigned to Sarraceniaceae and was described from rocks about the same age as those in which Archaefructus was found, i.e. ca 124 Ma (Li 2005), although it is probably a conifer gall (Herendeen et al. 2017 for literature). Otherwise the oldest fossils assignable to Ericales are in rocks ca 90 Ma old (Crepet et al. 2004; see also Martínez-Millán 2010). Indeed, in the late Cretaceous of E. North America there is a great diversity of fossil flowers that may belong to Ericales (Crepet et al. 2001, 2004; see also Herendeen et al. 1999; Friis et al. 2011). Some of these fossil flowers are quite unlike extant members of the clade, e.g. some have sepals with numerous huge abaxial and/or marginal glands (Crepet 2008), or are covered with scales on the outside, scales which also cover the gynoecium, the tiny flowers having the rather odd floral formula ↑ K [5] valvate; C 5; A 8 + 8 nectariferous staminodes; G [3] (Crepet et al. 2018: Teuschestanthes). This plant, from rocks some 90 Ma, comes out around Cyrilla and Clethra in morphological analyses, although with little support; note that its flowers are drawn inverted (Crepet et al. 2018). Schönenberger and Friis (2001) described Paradinandra suecica from the Late Cretaceous of Sweden, and this has a number of Ericalean features, some suggesting relationships with Pentaphylacaceae in particular (perhaps the relationships could more accurately be described as being with Ericales minus the Balsaminaceae and Polemoniaceae clades). Its placentation was intrusive parietal, the pollen was tricolpate, and there was a nectary disc around the base of the ovary; there were paired stamens opposite the petals and single stamens opposite the petals, as in some Sapotaceae, Ebenaceae, Styracaceae, Pentaphylacaceae and perhaps even Actinidiaceae (see also Friis 1985: ?Diapensiaceae; Keller et al. 1996: ?Actinidiaceae; Martínez-Millán et al. 2009: ?Pentaphylacaceae). However, Schönenberger et al. (2020) found over 100 places (maximum parsimony or MP + 1) in their angiosperm-wide analysis where Paradinandra might go, and although many were in crown-group Ericales, others were scattered throughout Pentapetalae, but not in gentianids... (see also López-Martínez et al. 2023a Sapindales or Malvales perhaps most likely). The tricolpate pollen of the fossil is uncommon in extant Ericales, being known only from Lecythidaceae and Balsaminaceae.
Evolution: Divergence & Distribution. For a good summary of the diverse Late Cretaceous fossils assigned to Ericales, see Friis et al. (2011).
Magallón et al. (2018) suggested that there was an increase in diversification at this node and dated it to (112.3-)107.8(-103.6) Ma. Diversification rate increases in the order are little nested - only 2/8 such events (Zuntini et al. 2024).
Sytsma et al. 2006 (see also Magallón et al. 2015; Wikström et al. 2015) thought that almost all families in Ericales had diverged by the early Eocene, some 50 Ma. However, Rose et al. (2018) found that all families had separated earlier, by ca 79 Ma in the Late Cretaceous, most major lineages separating 104-102 Ma, the crown-group age of the order being only ca 110.1 Ma (see also P. Soltis et al. 2019). Interestingly, stems of 18 of the 22 families of the order were ca 20 Ma or more, and over 50 Ma for Symplocaceae, with the exceptions of Ericaceae, with a stem of less than 2 Ma, and Clethraceae, Balsaminaceae and Sladeniaceae, all with stems of ca 10 Ma. Small, species-poor clades with long stems are commonly sister to more speciose clades (Rose et al. 2018). Rose et al. (2018) discuss many other aspects of diversification - clade ages (ibid.: Fig. 3, not all entered, Table 2), diversification rate changes (many) and biogeography - and their paper should be consulted for details. Ages in Bansal et al. (2021) also suggest rapid early diversification, although this was not the focus of that study; see also Schönenberger et al. (2005) and Larson et al. (2019/2020).
Schönenberger et al. (2005) examined character evolution in Ericales, which perhaps is beginning to make some sense, however, there is extensive homoplasy and relationships along the spine still need to be clarified (see below). Löfstrand and Schönenberger (2015b) looked at the evolution of characters around the Sarraceniaceae-Actinidiaceae area, but here, too, relationships are unclear (Larson et al. 2019/2020). For chemotaxonomy in the order from a phenetic point of view, see Rocha et al. (2015) and for pollen evolution, see Y. Yu et al. (2018).
Chartier et al. (2017: 36 characters, all 274 genera of the order) addressed the question of how clades (families, groups of families) occupied the total floral morphospace of the order, a morphospace (≡ morphological diversity, disparity, i.e. the difference in character states between two taxa) defined by variation in 37 floral characters. Somewhat to their surprise, they found that androecial characters contributed most to this disparity, androecial variation being particularly extensive in Lecythidaceae (see also elsewhere). The seven family groups (e.g. Ericaceae + Clethraceae + Cyrillaceae) plus three isolated families (e.g. Theaceae) that they found tended to be in different areas of this morphospace, although overall it was more or less continuous and these ten clades occupied adjacent areas of the morphospace (Chartier et al. 2017). Although only four families, Lecythidaceae, Ericaceae, Sapotaceae and Primulaceae (18% of the total), together contributed ca 50% to the total disparity in the order (Chartier et al. 2017), those families include ca 2/3 of all ericalean species, so although Chartier et al. (2021) found that disparity was decoupled from species richness, such figures do make one wonder. Overall, there is a latitudinal gradient of floral disparity here, disparity peaking in the tropics (see especially the categories trees, Africa), notably between 10 and 20o S, in part at least driven by plant-pollinator interactions (Chartier 2021). Parins-Fukuchi et al. (2021) saw a relationship between periods of morphological innovation - as in the androecium, meristic variation, etc. - and periods of phylogenomic conflict, and this was evident in the fossils, too. They dated this period to 90-70 Ma (see fossils) and 110-100 Ma (molecular ages).
Herting et al. (2023: 32 floral characters, 414 spp., all genera) returned to the issue of Ericalean floral diversity, looking at the rate of morphological evolution and disparity, one of its results, in the context of a more general comparison across all angiosperms (ibid: 18 characters, 792 spp.); the phylogeny/chronology of Ericales in Rose et al. (2018) framed the discussion. They found that staminal characters overall showed that highest rates of change of the three floral modules (perianth, androecium, gynoecium), both in Ericales and in angiosperms as a whole (but c.f. Magnoliidae and Malvidae), although individual staminal characters like "fusion of anthers" and "distal fusion of filaments" had very low rates of change and non-staminal characters like "flower length" and "flower diameter" the highest rates, and rates of corolla change were interestingly low in asterid groups (Herting et al. 2023: Figs 2, 5).
Ericales contain ca 5.9% of eudicot diversity (Magallón et al. 1999).
Ecology. Ericales are an important component of the understory in tropical rainforests, including ca 10% of the species and some 22% of the total stems (Davis et al. 2005a); families like Sapotaceae, Lecythidaceae and Ebenaceae are involved, however, one third of the order is made up of Ericaceae alone, not a noteworthy component of such rainforests, although they are quite common in tropical montane Malesian-American forests. This forest may have developed only early in the Caenozoic (Burnham & Johnson 2004), when the clades now making it up initially diverged; members of Malpighiales are the other main component of this vegetation. Lens et al. (2007b: characters of wood anatomy optimized on a tree - note topology), suggested that the ancestors of Ericales-Cornales grew under more temperate conditions in the present boreal-arctic area and later moved into tropical lowland rainforest.
Genes & Genomes. For a genome duplication, either for Ericales as a whole or still lower in the tree, see Larson et al. (2019/2020).
Viaene et al. (2009) discuss the complex history of PI gene duplication, sub- and neofunctionalisation, and loss in the clade. All taxa in which they found two copies of the PI gene have connate filaments; they thought that the PI gene may have facilitated floral diversification (Viaene et al. 2009). However, close attention should be paid to the nature of staminal connation and corolla tube formation which vary considerably in the order, and, as they noted, Primulaceae-Theophrastoideae have connate filaments but only a single copy of the PI gene.
Studies on the duplication of the RPB2 gene show that the I copy persists here almost alone in the [eudicots + Trochodendrales + Gunnerales] clade (and also in the lamiids: Oxelman et al. 2004). Many taxa lack the mitochondrial coxII.i3 intron, but it is known from Primulaceae-Maesoideae (and Balsaminaceae - plesiomorphic presence?) clades and also from Ebenaceae and Styracaceae (Joly et al. 2001).
Chemistry, Morphology, etc.. For leaf teeth that have a "?", their morphology is unknown. Schneider and Carlquist (2003) suggest that pit membrane remnants occur in some of this clade - perhaps mostly in some members of the terminal polytomy.
For details of ovary placentation, see Ng (1991). Truly parietal placentation does occur (e.g. Ericaceae-Pyroloideae), and although most other reports are incorrect, placentation at the apex of the ovary may be parietal (Löfstrand & Schönenberger 2015a). A stylar canal in the symplicate zone is common in the order (Löfstrand & Schönenberger 2015a).
For a summary of some chemical features of Ericales, see Grayer et al. (1999) and do Nascimento Rocha (2015: not easy to follow), for aluminium accumulation, see S. Jansen et al. (2004a, c), and for wood anatomy, see Lens (2005) and Lens et al. (2007b).
Phylogeny. Relationships within the order remain rather poorly understood; for early studies, see R. J. Bayer et al. (1996) and Morton et al. (1997a), both largely molecular data, and Anderberg (1992), morphological data. However, Polemoniaceae + Fouquieraceae, Myrsinaceae and relatives (i.e. Primulaceae s.l.), Ericaceae and relatives, and Balsaminaceae and relatives, these sister to the rest of the order, formed distinct clades, and a [Styracaceae + Diapensiaceae] clade was moderately (D. Soltis et al. 2000, 2007) or poorly (Albach et al. 2001b) supported, and these relationships are little different in Larson et al. (2019/2020). A study by Anderberg et al. (2002: five genes, both plastid and mitochondrial) had suggested a beginning of resolution of basal relationships within the order. B. Bremer et al. (2002) suggested a similar set of relationships, although the resolution (and sampling) is less extensive. Lecythidaceae, linked loosely with Sapotaceae in some earlier analyses (and versions 7 and earlier of this page) remain without a clear position (see also Wikström et al. 2015), indeed, Rose et al. (2018) found some support for a clade [Mitrastemonaceae + Lecythidaceae] that is sister to all other Ericales except Balsaminaceae and relatives; the latter are strongly supported as sister to all other Ericales although support for other relationships along most of the backbone of the order is weak. (The study by Rose et al. (2018) involved 4,531 ericalean taxa, 13 chloroplast, 9 mitochondrial and 3 ribosomal genes.) Interestingly, relationships within Lecythidoideae suggested by an analysis of 354 nuclear loci and of chloroplast genomes were rather different - Vargas et al. 2019.)
Geuten et al. (2004) carried out a Bayesian analysis of some 13 kb of nucleotide sequences, finding an inverted anther clade (Actinidiaceae, Ericaceae, etc.) perhaps sister to [[Theaceae s. str. + Symplocacaeae] [Styracaceae + Diapensiaceae]], all relationships with strong support in some analyses; Pentaphylacaceae and Primulaceae s.l. were sister taxa. However, Geuten et al. (2004) included only 16 terminals, for instance, the whole of the [[Sarraceniaceae [Actinidiaceae + Roridulaceae]] [Clethraceae [Cyrillaceae + Ericaceae]]] clade was represented by just two taxa. In a rather more extensive study employing some 59 terminals, nearly 20 kb of sequences, and a variety of analyses, Schönenberger et al. (2005) recovered a group differing only in some details from the Theaceae-Ericaceae-Sarraceniaceae clade just mentioned; they did not recover the [Pentaphylacaceae + Primulaceae] clade, rather, Primulaceae s.l. linked with Sapotaceae and Ebenaceae. However, Hao et al. (2010) note that the chimaeric nature of the mitochondrial atp1 gene in species of Ternstroemia (see Pentaphylacaceae) probably caused some of the odd findings in the Schönenberger et al. (2005) study. The relationships [Clethraceae [Actinidiaceae + Ericaceae]] were recovered in ML analyses of chloroplast genomes, but the positions of the first two reversed in Bayesian trees.
The peregrinations of taxa that used to be included in or near Theaceae are interesting. There were suggestions that Pentaphylacaceae, placed close to Theaceae by both Cronquist and Takhtajan, linked with Balsaminaceae, etc., in Ericales (Nandi et al. 1998). Prince (1998), although focussing on Theoideae, found that a) Theaceae were not monophyletic, and b) the two parts into which it split were associated with other Ericales included in the study. Thus Sladenia tended to associate with Theaceae s. str. in matK analyses, and although it was unplaced in morphological analyses, in these Ficalhoa was included in Theaceae s. str. (Pentaphylax was not included). Wei et al. (1999) compared the pollen of Pentaphylax with that of Clematoclethra (Actinidiaceae), another member of Ericales, and found the two to be similar. Pentaphylacaceae were associated with Gonocaryum (at that time in Icacinaceae) in an early study by Savolainen et al. (2000a), but this position has not since been recovered and Gonocaryum is now strongly associated with Aquifoliales as Cardiopteridaceae (e.g. D. Soltis et al. 2000; Kårehed 2001; Lens et al. 2008b). Tsou et al. (2016: Ficalhoa not included) placed Sladenia as sister to Freziereae in ITS analyses, yet it was sister to Pentaphylacaceae in trnL-F analyses, but support for the latter position in particular was very weak. Pentaphylax was placed sister to Ternstroemiaceae s. str. (Anderberg et al. 2001), and this is its resting place for now, but the limits of the family need confirmation. Rose et al. (2018) found a well supported clade of [Pentaphylacaceae + Sladeniaceae], but its position relative to the Sapotaceae-Primulaceae clade (poorly supported) was reversed when compared with that in the tree here, although support there, too, was low.
Both Pellicieraceae and Tetrameristaceae were in the Theales of Cronquist (1981). Pelliciera had been compared with Marcgraviaceae by Beauvisage (1920), while details of wood anatomy suggested relationships with Tetrameristaceae (Baretta-Kuipers 1976; see also above). Pellicieraceae and Tetrameristaceae formed a well-supported clade in the morphological analysis of Luna and Ochoterena (2004), but Marcgraviaceae did not join them, nor were other Ericales part of the clade. However, monophyly of balsaminoid clade is well supported, and it is probably sister to rest of Ericales (e.g. Källersjo et al. 1998; Nandi et al. 1998; Soltis et al. 2000, 2011; Savolainen et al. 2000a; Geuten et al. 2004). For relationships within the balsaminoids, see e.g. K. Bremer et al. (2004a) and Morton (2011: nuclear Xdh gene). If Balsaminaceae and Marcgraviaceae are sister taxa (Geuten et al. 2004; Janssens et al. 2009: ML bootstrap support 98%), there are no obvious synapomorphies for the family pair. Rose et al. (2018) and H.-T. Li et al. (2019: strong support, but only Pelliciera sampled in Tetrameristaceae) both recovered the relationships [Marcgraviaceae [Balsaminaceae + Tetrameristaceae]].
The position of the holoparasitic Mitrastemonaceae has been difficult to establish. Along with Cytinaceae and Rafflesiaceae, relationships of Mitrastemonaceae to Malvales have been suggested (Nickrent 2002). Barkman et al. (2004, also 2007: poor sampling) used mitochondrial sequences to place Mitrastemonaceae in Ericales, a position that also appeared in most analyses in Nickrent et al. (2004), and Hardy and Cook (2012) thought that Mitrastemonaceae were sister to most of Ericales except the Marcgraviaceae-Tetrameristaceae-Balsaminaceae clade. The cellular endosperm of Mitrastemon is certainly compatible with a position in Asterids, and its extrorse anthers are perhaps comparable with those of Ericaceae and their relatives, however, its parietal placentation and inferior ovary in particular are features found in many other parasitic angiosperms so they are not taxonomically particularly informative. Placing Mitrastemonaceae next to Ericaceae and their immediate relatives in the tree is partly for convenience; as mentioned above, Rose et al. (2018) found that it was sister to Lecythidaceae, albeit its position there was weakly supported.
Details of relationships in the tree used here up to early 2020 - [[Marcgraviaceae, etc.] [[Polemoniaceae etc.], Lecythidaceae [[Pentaphylacaceae, etc.], [Sapotaceae [Ebenaceae + Primulaceae]] [Mitrastemonaceae, Theaceae, [Styracaceae etc.], [[Sarraceniaceae [Actinidiaceae + Roridulaceae]] [Clethraceae [Cyrillaceae + Ericaceae]]]]]]] were adapted to follow those suggested by Schönenberger et al. (2005), etc., however, caution was clearly in order when using this phylogeny. Thus the tree in Duangjai et al. (2006b) also shows support for Lecythidaceae sister to most other Ericales, if with rather weak (73% bootstrap) support, but relationships in the order were not the focus of that study). The relationships just mentioned were largely recovered by Sytsma et al. (2006), and with strong support, but c.f. in part Soltis et al. (2011: ?sampling). Hardy and Cook (2012) recovered rather different relationships, thus Fouquieraceae were not sister to Polemoniaceae, Symplocaceae were sister to the Cyrillaceae-Clethraceae-Ericaceae clade. The grouping [Sarraceniaceae [Actinidiaceae + Roridulaceae]]] was recovered by Löfstrand and Schönenberger (2015b), but support for the position of Sarraceniaceae was not strong (see also Ellison et al. 2012). Relationships suggested by Z.-D. Chen et al. (2016: Chinese taxa) differed considerably from those above, but support was poor. X.-Q. Yu et al. (2017) recovered Primulaceae as sister to Pentaphylacaceae and other Ericaceae that they examined, but this may be a sampling artefact; Theaceae were the focus of their study. In a study of plastid genomes, M. Yan et al. (2018) found that Polemoniaceae were embedded in the Sapotaceae-Primulaceae clade, although support was weak and this, too, may be a sampling problem. H.-T. Li et al. (2019) found very low support for relationships along the backbone of the tree below a clade including [Theaceae [Symplocaceae [Styracaceae + Diapensiaceae]]], Ericaceae, etc., the relationships they found being rather different from those below, and there was a [Sapotaceae + Primulaceae] clade, but overall there was little support (see also Gitzendanner et al. 2019).
From the above confusing summary, it is evident that relationships in Ericales have been rather much up in the air. Indeed, S. A. Smith et al. (2013) had found that six of the top 25 seed plant clades with the lowest resolution were in Ericales, and relationships within the order were clearly in a bit of a mess (ibid., e.g. Figs 2-4). Larson et al. (2019/2020) verified such problems in a critical series of phylogenomic analyses using coding sequence data from 97 taxa; five small families, including Sladeniaceae and Mitrastemonaceae, were not included. Larson et al. (2019/2020) suggested that relationships in the order were best represented as [[Marcgraviaceae, etc.] [Sapotaceae, Ebenaceae, [Polemoniaceae + Fouquieraceae], Lecythidaceae, Primulaceae, [Pentaphylacaceae [[Theaceae [Styracaceae, etc.]] [Sarraceniaceae [Ericacaeae [Actinidiaceae + Roridulaceae]]]]]]], i.e., including a hexatomy, furthermore, support for the position of Sarraceniaceae was not very strong. Support along the spine of the tree recovered by C. Zhang et al. (2020: Fig. 1) was rather weak and there were alternative topologies possible. They recognized a clade, "Core Ericales", which included [[Sladeniaceae + Pentaphylacaceae] [Theaceae [the rest as below]]]], [Balsaminaceae + Marcgraviaceae] were sister to all the rest of the order, but families in between... Relationships recovered by W. J. Baker et al. (2021a) in the Seed Plant Tree, a preliminary analysis of Angiosperms353 nuclear data, are different yet again - Balsaminaceae, etc., still form the basal group, but then relationships are [Sladeniaceae [Theaceae [Lecythidaceae [[Mitrastemonaceae (v. low support) [[Polemoniaceae + Fouquieraceae] [Ebenaceae + Primulaceae]]] [[Pentaphylacaceae + Sapotaceae] [[Symplocaceae group] [Sarraceniaceae ...]]]]]]]. In the plastome analyses of H.-T. Li et al. (2021) the corresponding relationships are [basal group [[Pentaphylacaceae + Sladeniaceae] ... [[Theaceae [Symplocacaceae [Styracaceae + Diapensiaceae]]] etc.]]]; the branches on the spine between (...) have vanishingly little support and the exact position of Theaceae is unclear. Larson et al. (2023: Angiosperms353 data, focus on Primulaceae) recovered the following relationships: [Lecythidaceae [[[Polemoniaceae + Fouquieriaceae] [Ebenaceae + Primulaceae]] [Sapotaceae [[Sladeniaceae + Pentaphylacaceae] [[Theaceae [Symplocaceae [Styracaceae + Diapensiaceae]]] [[Roridulaceae + Actinidiaceae] [Clethraceae + Ericaceae]]]]]]] - some families were not included.
Our understanding of relationships here does seem to be in a mess. The ix.2024 version of the Seed Plant Tree did not include Mitrastemonaceae, but beyond that relationships are [[Balsaminaceae [Tetrameristaceae + Marcgraviaceae]] [Polemoniaceae [Primulaceae [[Lecythidaceae [Fouquieraceae + Ebenaceae]] [Sapotaceae [[Sladeniaceae + Pentaphylacaceae] [[Theaceae [Symplocaceae [Styracaceae + Diapensiaceae]]] [Sarraceniaceae [[Roridulaceae + Actinidiaceae] [Clethraceae [Cyrillaceae + Ericaceae]]]]]]]]]]]. On the other hand, relationships in Zuntini et al. (2024) are [Mitrastemonaceae [[Balsaminaceae [Tetrameristaceae + Marcgraviaceae]] [Lecythidaceae [[[Fouqieraceae + Polemoniaceae] [Ebenaceae + Primulaceae]] [[Sapotaceae [Sladeniaceae + Pentaphylacaceae]] [[Theaceae [Symplocaceae [Styracaceae + Diapensiaceae]]] [Sarraceniaceae [[Roridulaceae + Actinidiaceae] [Clethraceae [Cyrillaceae + Ericaceae]]]]]]]]]] - the last six families stand in the same relationships in the two, but that is about all that can be said. Indeed, C. C. Yu et al (2025: focus on Sladeniaceae and Pentaphylacaceae) found very different relationships in analyses of plastome (43 plastome, 17 families) and Angiosperms353 (278 gene trees, 25 genera) data sets which in part they suggested might be explained by ILS involving ancestral plastid polymorphisms. Relationships obtained from the analysis of nuclear data were [[Polemoniaceae + Primulaceae] [Lecythidaceae [Ebenaceae [Theaceae [[Symplocaceae [Styracaceae + Diapensiaceae] [Actinidiaceae [Clethraceae + Ericaceae]] [Sapotaceae [Sladenia [Ficalhoa + Pentaphylacaceae]], while those obtained from the plastome data were [Lecythidaceae [[Polemoniaceae + Fouquieraceae] [[Sapotaceae [Ebenaceae + Primulaceae]] [Actinidiaceae [Clethraceae + Ericaceae]] [Theaceae [Symplocaceae [Diapensiaceae + Styracaceae]]]] [[Sladenia + Ficalhoa] Pentaphylacaceae]]]]] (note that there is yet another topology in Yu et al. 2025 - see Fig. 6).
Previous Relationships. Theales of Cronquist (1981) included mostly families now in Malvales, Ericales, and Malpighiales, and Takhtajan's Theanae were largely equivalent (Takhtajan 1997). Ericales as here delimited are made up largely of Sarracenianae, Ericanae, Primulanae, and some families in Theanae, all adjacent groups in the Dilleniidae of Takhtajan (1997); its members are more widely scattered in Cronquist (1981). This is the asterid III group of some early phylogenetic studies.
Synonymy: Ericopsida Bartling - Ericidae C. Y. Wu et al. - Lecythidaneae Reveal - Barringtoniineae J. Presl, Empetrineae Link, Epacridineae Link, Ericineae Link, Primulineae Burnett, Pyrolineae J. Presl, Rhododendrineae J. Presl, Sarraceniineae Reveal, Scytopetalineae Engler, - Actinidiales Reveal, Aegiceratales Martius, Ardisiales J. Presl, Balsaminales Link, Barringtoniales Martius, Camelliales Link, Cyrillales Doweld, Diapensiales Engler & Gilg, Diospyrales Prantl, Ebenales Engler, Empetrales Martius, Epacridales Berchtold & J. Presl, Fouquieriales Martius, Gordoniales J. Presl, Halesiales Link, Lecythidales Martius, Lysimachiales Döll, Marcgraviales Martius, Mitrastemonales Makino, Monotropales Berchtold & J. Presl, Myrsinales Berchtold & J. Presl, Polemoniales Berchtold & J. Presl, Primulales Berchtold & J. Presl, Rhodorales Horaninow, Roridulales Nakai, Samolales Dumortier, Sapotales Berchtold & J. Presl, Sarraceniales Martius, Styracales Martius, Ternstroemiales Martius, Theales Berchtold & J. Presl, Vacciniales Dumortier
[Marcgraviaceae [Balsaminaceae + Tetrameristaceae]] / balsaminoid clade: non hydrolysable tannins [myricetin] +, ellagic acid 0; raphide sacs +, druses 0; vessels in radial multiples; paratracheal parenchyma +; ± branched sclereids +; lamina supervolute, elongating in bud, with obscure abaxial lines, toothed; inflorescence racemose; bracteoles immediately below the flower; cells with mucilage, tannins +; K and C rather similar in size and colour, K with a single trace at base; abaxial surface of C with stomata; nectary outside A; stamens = and opposite sepals, free from C, anthers (near) basifixed, thread-like structures along the stomium, filaments broad; mucilaginous secretion in the ovary, style short, stigma little expanded; ovules bitegmic; style persistent in fruit; endosperm at most slight.
Age. This node was dated to just under 65 Ma by K. Bremer et al. (2004a), to (66-)52, 49(-34) Ma by Bell et al. (2010), ca 55.4 Ma by Magallón et al. (2015), ca 51.1 Ma by Fu et al. (2019), and ca 43.6 or 45.2 Ma by Tank et al. (2015: Table S1, S2); the estimate was ca 81.7 Ma in Rose et al. (2018). With different topologies, the common ancestor of this clade was estimated at (52-)48, 44(-40) Ma (Wikström et al. 2001: Balsaminaceae sister to the rest) or (85-)59(-38) Ma (Wikström et al. 2015) and that of the [Balsaminaceae + Marcgraviaceae] clade was dated to the middle Palaeocene ca 58.9 Ma (Janssens et al. 2009).
Evolution: Divergence & Distribution. Looking at the floral morphospace of Ericales, Chartier et al. (2017) thought that this clade was morphologically rather homogeneous, although they noted that they did not include features like the colour and shape of perianth members in their analysis.
If Balsaminaceae and Marcgraviaceae are sister taxa (Geuten et al. 2004; Janssens et al. 2009: ML bootstrap support 98%), there are no obvious synapomorphies for the family pair, even when one examines floral development closely (see Schönenberger et al. 2010; Schönenberger & von Balthazar 2010; von Balthazar & Schönenberger 2013). Some of the similarities between Tetrameristaceae and Marcgraviaceae may be because they are both woody, but, depending on the topology, they could be synapomorphies/plesiomorphies for the whole clade, the corresponding features of Balsaminaceae being apomorphies for that family.
Geuten et al. (2006) suggest that heterotopic SEP3-like gene expression in bracteoles and calyx in members of this clade was present in its common ancestor; the gene is normally expressed in the corolla and other inner whorls. Indeed, most taxa in this clade have more or less petal-like sepals, bracts or bracteoles, although Tetramerista and Pentamerista, derived members, lack such expression patterns. Schönenberger et al. (2010: see also Schönenberger & von Balthazar 2010) suggest that thread-like structures bordering the stomium - they vary in their morphological nature - are an apomorphy for the whole clade. Schönenberger et al. (2010) emphasized that the nectary was situated in the periphery of the flower, that there are mucilage cells in the flower, etc., while von Balthazar and Schönenberger (2013) discussed these and other possible apomorphies of the clade.
Chemistry, Morphology, etc.. Beauvisage (1920) noted that both Pelliciera (Tetrameristaceae) and Marcgraviaceae have large air spaces in the cortex. The raphide sacs are white pockets in the stem; they are visible under the dissecting microscope.
For the wood anatomy of this group, see Lens et al. (2005b), that of Balsaminaceae is paedomorphic, for palynology, see Lens et al. (2005c: Marcgraviaceae) and Janssens et al. (2005: the rest), and for general floral morphology, see Schönenberger et al. (2010) and in particular von Balthazar and Schönenberger (2013).
MARCGRAVIACEAE Berchtold & J. Presl, nom. cons. - Back to Ericales
Lianes, climbing by weakly twining and/or roots, and/or hemiepiphytes; (vessel elements with scalariform perforation plates); rays broad [Marcgravia]; petiole bundle deeply arcuate, or annular with inverted adaxial plate; cells with essential oils; stomata staurocytic; lamina margin entire, with marginal to abaxial cavities (black dots); inflorescence often umbellate; bracts abaxially ascidiate, nectariferous, recaulescent on pedicel; K quincuncial; A many; G [2-20], opposite ?, placentation very intrusive parietal, stigma?, dry; ovules many/carpel; fruit with more or less irregular dehiscence, placentae fleshy; seeds many, small; exotestal cells ± enlarged, inner walls much thickened; endosperm with micropylar haustorium, cotyledons large to small; x = 18.
7 [list]/130 - two subfamilies below. New World tropics (map: from Heywood 1978). [Photo - Flowers, Fruits.]
Age. Diversification in Marcgraviaceae is estimated to have begun ca 23.9 Ma by Rose et al. (2018).
1. Marcgravioideae Choisy - Marcgravia L.
Leaves strongly heterophyllous, 2-ranked; apical flowers sterile, nectariferous; flowers 4-merous; C connate, calyptrate [calyptra thrown off at anthesis]; G ?; inner integument ca 3 cells across; embryo sac long; n = (?16, 19), Cx [diploid spp.] = 2.55-4.04 pg.
1/60. New World Tropics.
2. Noranteoideae Choisy —— Synonymy: Noranteaceae Martius
Also shrubs; leaves spiral; inflorescence also spike; nectaries associated with each flower, (bracts with paired, C-like appendages); flowers 5-merous; C basally connate, reflexed, deciduoud; (A 4-15), (style +), stigma wet [Souroubea]; (ovules 5<); n = (?16, ?17, 19), Cx [diploid spp.] = 5.55-6.2 pg.
6/70: Souroubea (19). New World tropics.
Evolution: Ecology & Physiology. Heterophylly is common in Marcgravia, the climbing form of the plant - it is a root climber - having orbicular leaves that are closely adpressed to the trunk. Little definite is known about the function of such leaves (Zona 2020).
Evolution: Pollination Biology & Seed Dispersal. The distinctive inflorescences with nectar secreted in the cup-shaped (ascidiate) bracts attract a variety of large pollinators including hummingbirds and bats (e.g. Dressler 1999; Tschapka et al. 2006; Fleming et al. 2009). Although individual flowers of Marcgraviaceae are polysymmetric, the inflorescences of Margravia and the flowers of taxa like Souroubea are monosymmetric from the point of view of their pollinators, which get pollen dusted on their heads (usually) as they take nectar from the modified bracts (see also Westerkamp & Claßen-Bockhoff 2007). Marcgravia evenia is a bat pollinated plant that has a concave bract above the inflorescence which reflects the signals of echo-locating nectarivorous bats, so helping them find the flowers more readily (Simon et al. 2011). Pollen of S. guianensis is associated with oil produced by the tapetum (Machado & Lopes 2000).
Seeds of Marcgraviaceae may quite often be dispersed by fruit-eating bats (Lobova 2009).
Genes & Genomes. Some polyploid species in The Rest have massive genomes of 2C = 14-21.5 pg or so (Schneider et al. 2014b).
Chemistry, Morphology, etc.. For the epiphytic habit in the family, see Zotz (2013). The black dots on the margin of the leaf blade make the leaf appear "serrate", but here that character is not so much about serrations per se, as the marginal glands, etc., that terminate any serrations that are present.
I do not know if the pollen grains are starchy (c.f. Balsaminaceae). Juel (1887) shows both integuments as being two cells across. Johri et al. (1992) described the seeds as being arillate.
For general information, see Dressler (2004) and Schneider et al. (2014b), for vegetative anatomy, see Beauvisage (1920), and for information on embryology in Marcgravia, see Mauritzon (1939c).
Phylogeny. Ward and Price (2002) suggest phylogenetic relationships within the family. Marcgravia, with its reversible heterophylly, two-ranked leaves, 4-merous flowers, calyptrate corolla, and nectaries adnate to abortive flowers, is distinct, but in the rest of the family both synapomorphies and generic limits are unclear.
[Balsaminaceae + Tetrameristaceae]: bracteoles apical; K ± C-like, with adaxial nectar glands; A with anthers latrorse, filaments postgenitally adnate to ovary; G [5], stylar canal +, 5-radiate, stigma with often inconspicuous commissural lobes; ovule 1/carpel, ?orientation, with chalazal constriction, funicle stout; K not persistent in fruit; endosperm with micropylar haustorium.
Age. K. Bremer et al. (2004a) dated this node to around 56 Ma, while around 50.5 Ma was the age in Magallón et al. (2015), (77-)51(-30) Ma in Wikström et al. (2015), ca 42.7 Ma in Tank et al. (2015: Table S2), and as much as ca 81 Ma in Rose et al. (2018).
Evolution: Divergence & Distribution. For possible apomorphies of this clade, see Schönenberger et al. (2010), Schönenberger and von Balthazar (2010), and von Balthazar and Schönenberger (2013).
BALSAMINACEAE A. Richard, nom. cons. - Back to Ericales
Fleshy herbs; non-hydrolysable tannins, naphthoquinones +; cork?; vessels single; sclereids 0; petiole bundle arcuate; mucilage sacs +; plant glabrous; nodes swollen; lamina vernation involute, extrafloral nectaries +/0 [paired glands or foliaceous lateral flaps on leaf base/stem]; inflorescence axillary; flowers vertically monosymmetric, resupinate [median sepal abaxial]; functionally abaxial sepal with prominent nectariferous spur, C 5; anthers postgenitally ± connate and forming cap over stigma, anthers polysporangiate, septae tapetal, filaments stout; tapetal cells 2(-4)-nucleate; (branched) cellulose threads from cell walls holding pollen grains to anther [?both genera], with raphides, starchy, pollen oblate, triangular in polar view, endexine lamellate; G [5], opposite C, stigma fairly broad, wet; ovules (hemitropous), apotropous, suprachalazal zone long, with elongated cells; embryo sac bisporic, 8-nucleate [Allium type], becoming very long; seed pachychalazal; endosperm also with chalazal haustorium, (xyloglucans +), cotyledons large; x = 9, nuclear genome [1 C] (0.079-)1.028(-13.407) pg; germination epigeal, radicle soon dies [?all].
2/1,100: [list]. Cool temperate to tropical, not Central and South America, the Antipodes, or Oceania. Map: from Hultén (1971), Meusel et al. (1978), Grey-Wilson (1980a), Hultén and Fries (1986), and Trop. Afr. Fl. Pl. Ecol. Distr. vols 1 (2003), 6 (2011).
Age. There are suggestions that diversification in Balsaminaceae began (39.3-)30.7(-22.1) Ma (Janssens et al. 2009), (41.0-)30.7(-21.2) Ma (Y.-F. Liang et al. (2025) or ca 69.6 Ma (Rose et al. 2018).
1. Hydrocera triflora (L.) Wight & Arnott —— Synonymy: Hydroceraceae Wilbred
Plant semi-aquatic; tapetum amoeboid; G loculi divided longitudinally into three parts; ovules apically bitegmic, micropyle endostomal, outer integument 8-9 cells across, inner integument 6-7 cells across; fruit a drupe, stones 5, separating, each with two longitudinal air sacs; testa sclerotic, 6-8 layers thickened cells, ca 5 layers unthickened; endosperm with micropylar haustorium several nucleate, protruding through the micropyle, chalazal haustorium uniseriate, embryo suspensor short [2-3-celled]; n = 8.
1/1. South India, Sri Lanka, Hainan to Peninsula Malaysia, Java, the Celebes.
2. Impatiens L. —— Synonymy: Impatientaceae Barnhart
Plant rhizomatous, (annual/pachycaul treelet/tuberous/epiphytic); (indumentum +); leaves (opposite); inflorescence (cymose), ± many-flowered, (bracteoles 0); (flowers not resupinate); K often 3, (spur 0), lateral C basally connate in pairs, (all four connate), adaxial C often with a K-like keel [esp when K = 1]; pollen (porate), 4(-3, 5)-aperturate, elliptic, rectangular in polar view, etc.; style at most short, stigmas 1-5; ovules (1-)several/carpel, uni- to biseriate, bitegmic, cuter integument 2-10 cells across, inner integument 2-6 cells across/unitegmic 9-15 cells across; (embryo sac unisporic, 8 nucleate); fruit a septifragal capsule, explosive [walls inrolling from base]; only exotestal cells thickened, surface smooth/verrucose/with unicellular hairs with reticulate or spiral thickenings/warty-glandular [unicellular]/clavate appendages, (mucilaginous); n = (3-)7-10(-12<); nuclear genome [1 C] (1125-)1922(-3182) Mb; (hypocotyl elongates).
1/>1,100. Cool temperate to tropical, not Central and South America, the Antipodes, or Oceania, most Old World, i.e. Africa (esp. Madagascar) to the mountains of Southeast Asia. Photo: Flower.
Age. Diversification within Impatiens may have begun (28.1-)22.5(-16.9) Ma (Janssens et al. 2009, 2012a) - there I. omeiana was sister to the rest of the genus, which began diversifying a little over 15 Ma, ca 59.3 Ma (Rose et al. 2018: [I. desmantha + I. gongshanensis] sister, i.e., corresponding to the 15 Ma age...) or (33.1-)24.8(-17.0) Ma (Y.-F. Liang et al. 2025).
2A. subg. Clavicarpa S. X. Yu & W. Wang
inflorescences racemose; sepals 4, lateral; pollen 3-aperturate; G [4]; one ovule/carpel; capsule hammerhead-shaped; seed ellipsoid, testa reticulate ornamentation.
30 spp.
Pollen 4-aperturate, oblong, circular, elliptic or quadrate; G [5]; many ovules/locule; capsule fusiform, linear, clavate.
[Semeiocarpidium + Racemosae]: .
Age. The age of this clade is (21.4-)15.6(-10.3) Ma (T. Xue et al. 2025).
2B1. Sect. Semeiocarpidium (Zollinger) S. X. Xu & W. Wang
4 lateral K in two pairs, lower pair enclsing upper pair and abaxial K, abaxial K with bifid spur; wing C connate; G [4]; capsule obconic (cylindrical); seed long-plumose or granulate.
60 spp.
2B2. Sect. Racemosae J. D. Hooker & Thomson
inflorescence racemose, many [>5]-flowered; lateral K 2 (4, inner 2 reduced); capsules linear; seeds ovoid, "digitate seed coat ornamentation".
70 spp. S.W. China and the Himalayas (to Central Asia).
Age. The crown group age of section Racemosae is (12.0-)8.7(-5.6) Ma (T. Xue et al. 2025).
[Fasciculatae [Tuberosae [Impatiens [Scorpioidae + Uniflorae]]]]:.
2B3. Sect. Fasciculatae S. X. Yu & W. Wang
Inflorescence fasciculate, 2--3-flowered; lower K large, saccate; inflorescence subumbelluate, peduncle very short; K 4, fully developed; pollen elliptic; capsule long-fusiform, ± straight; seeds subpherical/ellipsoid.
20 spp.
[Tuberosae [Impatiens [Scorpioidae + Uniflorae]]]:.
2B4. Sect. Tuberosae S. X. Xu & Wei Wang
Roots tuberous; inflorescences racemose, many-flowered; sepals 4, lateral, fully developed; pollen 4-aperturate; G (5); capsules clavate; seeds many/locule, ellipsoid.
[Impatiens [Scorpioidae + Uniflorae]]:.
Inflorescence 2-(many-)flowered.
90 spp., esp. Hengduan Mountains and Southeast China, some scattered in the Northern Hemisphere.
Age. The crown-group age of this section is (16.1-)11.8(-8.1) Ma (F. Qin et al. 2023).
2B6. Sect. Scorpioidae S. X. Yu & Wei Wang
Cymes subscorpioid; lower K large, saccate, inner 2 of 4 lateral sepals scale-like; capsule long, curved; seeds subspherical, testa minutely hummocky.
2B7. Sect. Uniflorae J. D. Hooker & Thomson
(Foliar extrafloral nectaries 0 - T.); inflorescence fasciculate, 2(–3)-flowered (racemose); (flowers greenish, brown to blackish or yellowish (never white, pink or purple), spur 0, lower sepals boat-shaped - T.); (anthers obtuse or truncate, apically dehiscing - T.); capsule short-fusiform, conspicuously turgid at middle, ca. 1 cm long (long-fusiform); seeds ellipsoid.
260 spp., esp Madagascar (the erstwhile sect./subg./genus Trimorphopetalum).
Evolution: Divergence & Distribution. Note that the ages above for diversification in the genus are wildly different.
Impatiens is most diverse in tropical and subtropical montane forests, and the imbalance in species numbers between it and the monotypic Hydrocera is striking. Within Impatiens diversification began only in the Early Miocene (28.1-)22.5(-16.9) Ma, but the speciation rate then much increased in the early Pliocene within the last 5 Ma, climate change causing much population fragmentation, isolation, and migration (Janssens et al. 2009).
The Southeast Asian/China region may be the centre of origin for the genus (Janssens et al. 2009; T. Xue et al. 2025). In Thailand in particular limestone is a favoured habitat, 45 of the 58 endemic Thai species growing there (Suksathan & Ruchisansakun 2020). Grey-Wilson (1980c) noted that species of Impatiens with long-linear fruits were Eurasian and American, species with fusiform fruits African and Malesian, with China to Thailand being the main area of overlap.
- There are over 260 species of Impatiens on Madagascar; monophyletic, it is the largest plant genus there, and ca 127 species belong to section Trimorphopetalum, the flowers of which lack spurs (Abrahamczyk et al. 2021) - see below for pollination. The stem group age of this section is ca 4.1 Ma, that of the crown group ca 3.8 Ma (Abrahamczyk et al. 2021, see also Janssens et al. 2009, but reduced sampling leads to a Pleistocene crown-group age).
- There is a basal split in section Impatiens between a clade from Southeast China and a much larger clade centred on the Hengduan Mountains (F. Qin et al. 2023); there has been subsequent dispersal around the Northern Hemisphere, especially in the former clade, while within the latter clade there has been subsequent dispersal to Southeast China, and so on... The original habitats of members of the section may have been relatively cool and at middle elevations (Qin et al. 2023, q.v. for more on diversification and biogeography here).
- The common ancestors of section Racemosae and also of the four main clades within it are likely to have originated in the Himalayas, with orogeny and climate change driving their diversification (T. Xue et al. 2025). However, there is quite extensive conflict between the plastome and nuclear trees in the relationships of the species in the two largest clades, but the nuclear tree was based on rather scant data (nuclear ribosomal genes - Xue et al. 2025), so the significance of the conflict is unclear.
The combination of non-hydrolysable tannins and raphides, both of which are found in Balsaminaceae, is rarely found in herbs (Fischer 2004a). However, the family is indeed likely to be primitively herbaceous. Increase in width of the stem is by expansion of pith cells (Troll & Rauh 1950), and although some species of Impatiens do produce a small amount of wood, it is derived and paedomorphic (Smets et al. 2011; Lens et al. 2012a).
Janssens et al. (2012a) included 13 palynological characters along with molecular data in their study of relationships within Impatiens; there was some phylogenetic signal in the palynological features. For possible apomorphies of the two genera and of major clades within Impatiens, see S.-X. Yu et al. (2015). Y.-X. Song et al. (2023) looked at the evolution of five pollen and nine vegetative and reproductive traits in Impatiens. There has been a fair amount of work on seed morphology and evolution. Song et al. (2022) optimized variation in a number of features of seed morphology, i.e. shape, primary and secondary ornamentation, and anticlinal and periclinal walls, onto five trees; there was some phylogenetic signal there. In a similar study, X. Zhang et al. (2025), with a focus on Chinese species, a somewhat more elaborate division of seed morphology, and a tree with a somewhat different topology, carried out a similar study. In both papers it was notable that seeds of species of subgenus Clavicarpa showed practically no variation in the characters studied and were similar in morphology to seeds of Hydrocera, thus suggesting the plesiomorphic state for the family as a whole (these papers should be read carefully for their justification of characters and states). Indeed, testal morphology here shows a considerable amount of variation, Utami and Shimizu (2005: p. 455) noting "The genus Impatiens has a most diverse and elaborately sculptured seed coat." This is something of an understatement, as a glance at the remarkable seeds of I. xanthina will confirm (see Song et al. 2022: Fig. 5E. For more on seed morphology, see Song et al. 2005; Cai et al. 2013; also Rewicz et al. 2025 for a bibliography).
Balsaminaceae are vegetatively quite variable, ranging from annuals to small and pachycaul trees; in a group of annual species from Thailand, the hypocotyl keeps elongating (Suksathan & Ruchisansakun 2020). Florally they are very diverse; there has been duplication and probable subfunctionalisation of the class B DEF gene in this clade (Janssens et al. 2006b; Geuten et al. 2006).
Ecology & Physiology. The invasive Impatiens glandulifera can have major effects on the aquatic communities in the places where it grows (it has been widely introduced) via the allelopathic effects of the quinone 2-methoxy-1,4-naphthoquinone that it produces; these effects are both direct via negative effects on the growth and survival of chironimid midge larvae, for example, and indirect, affecting other members of food chains that would otherwise eat those larvae (Hüftlein et al. 2024).
Pollination Biology & Seed Dispersal. For some time, rather little was known about pollination in Impatiens other than a few accounts of individual species (e.g. Tian et al. 2004), although recent work is beginning to address this absence. Thus Abrahamczyk et al. (2017b) carried out a morphometric analysis of some 86 species and recovered six clusters (so long as quantitative, not categorical, data were used) corresponding to pollination syndromes and to records of pollinators in the literature. Interestingly, there were two separate clusters made up of bird-pollinated species, clusters that included large and small birds respectively, another cluster seemed not to represent any particular syndrome, and one or two species like Impatiens glandulifera were isolated, in that case probably because it has over four times the amount of pollen than that of any other species examined (Abrahamczyk et al. 2017b; Lozada-Gobilard et al. 2019). Ruchisansakun et al. (2020) carried out a more extensive analysis of floral morphology, i. a. noting that 19% of the species they examined had asymmetrical flowers, and that shifts to such flowers were associated with bee pollination; they questioned the possibility of fly pollination suggested by Abrahamczyk et al. (2017b), but see below. Vandelook et al. (2019) found that there was some correlation between nectar composition and amount with pollinator in Impatiens, for instance, bird-pollinated flowers had copious sucrose-rich nectar and the one fly-pollinated species examined had nectar low in quantity but rich in amino acids and hexoses, even if bee-pollinated flowers also tended to have high amounts of amino acids. Pollen/ovule ratios, often correlated with pollination syndromes, were not linked here (Lozada-Gobilard et al. 2019).
Interestingly, species of the monophyletic Madagascan Impatiens section Trimorphopetalum (included in sect. Uniflorae), plants of the forest floor in eastern Madagascar, have small, spurless flowers that are dark dull purple, brownish, yellowish or greenish in colour, the plant lacks foliar extrafloral nectaries and quite often much in the way of floral nectar, etc. (illustrations in Rahelivololona et al. 2018; Abrahamczyk et al. 2021). Lozada-Gobilard et al. (2019) noted that the flowers had a fly pollination syndrome, and three of the species that they examined were selfers-cleistogamous. Abrahamczyk et al. (2021) observed that some species had greenish, cup-shaped flowers that were pollinated by small insects that also took nectar from the flower, while other species had more open, reddish to yellowish brown flowers that looked - and smelled - rather like fungal fruiting bodies and were visited mostly by female phorid flies, but also some other insects; phorids are elsewhere known to lay eggs on fungi. Abrahamczyk et al. (2021) noted that the number of pollen grains and ovules per flower were more or less strongly reduced in this clade, the former particularly in the cleistogamous species.
The flowers of Impatiens are protandrous. As the anther walls break down and then retract, the cellulose threads produced hold the exposed pollen over the stigma as if in a lattice (Vogel & Cocucci 1988); the pollinator picks up the pollen from there. As the flower ages, the stamens fall off and the stigma is exposed.
The common names for some Impatiens such as Busy Lizzie and Touch-Me-Not refer to the explosive dehiscence of the ripe fruits which can be triggered by gently squeezing them; the valves separate and incurve (rather unusual), flinging out the seeds as they do so. In I. glandulifera ca 80% of the septal area cracks before the explosive event itself, and the stored elastic energy efficiently used results in the seeds being dispersed with an initial velocity of up to 4 m/s, although not all species have exactly the same mechanism/energy efficiency (Deegan 2012; see Tian et al. 2021 for the application of Impatiens-type bending in the construction of dual-function electronic devices). Indeed, in I. capensis, only ca 0.5% of the stored energy is transmitted to the seed, the great majority of which are predicted to be thrown around 0.5 m or less (Hayashi et al. 2009). The seeds are not very dense, being rich in lipids, which also makes this initial dispersal less efficient, but they may also be secondarily dispersed by water (Hayashi et al. 2009), indeed, the plant, native in North America, is an effective competitor.
Genes & Genomes. There may be a genome duplication in Impatiens (Larson et al. 2019/2020; = INPAα, Leebens-Mack et al. 2019). Cytological variation in Impatiens is considerable; species with n = 9 often have a bimodal karyotype (Y. Song et al. 2003).
The very variable Impatiens morsei has a notably small plastome with the smallest Small Single Copy region of any Impatiens examined, NADH genes there being lost (H. Qiu et al. 2023).
Chemistry, Morphology, etc.. The paired glands or foliaceous lateral flaps on the leaf base or stem near the leaf base are at least sometimes vascularized (Colomb 1887). The area between the separate vascular bundles remains parencyhmatous even as secondary thickening develops (Cunha Neto 2023).
For an interpretation of floral morphology in which two of the sepals, the adaxial-lateral pair (non-inverted orientation), are perhaps better interpreted as prophylls/bracteoles borne immediately below the flower, as in other members of the balsaminoid clade, see von Balthazar and Schönenberger (2013). Given the evidence presented, this interpretation seems reasonable, and is followed above. The abaxial-lateral sepal pair is often reduced, perhaps becoming fused with the abaxial petal, or it is entirely absent (Caris et al. 2006a; see also Grey-Wilson 1980c). Interestingly, in Impatiens with three sepals there are four carpels, the adaxial carpel being larger than the other three (S.-X. Yu et al. 2010).
Janssens et al. (2012b) notes the variety of stamen form that is obscured by the bland statement "anthers connate and forming cap over stigma" (see above). The integuments are quite thick and are free only at the micropyle (e.g. van Tieghem 1898); L. L. Narayana (1970) also illustrates more conventional ovules. Studies by McAbee et al. (2005, see also 2006) show considerable variability in integument development in the family, many species having more or less well developed congenital fusion of the integuments - this has evolved more than once, and bitegmy may be derived. There is variation in the embryo sac, Hydrocera and at least some species of Impatiens having a bisporic, 8-celled embryo sac (Venkateswarlu & Lakshminarayana 1958); this may be an apomorphy for the family. The fruit type of Hydrocera is unclear (see Wood 1975), Venkateswarlu and Lakshminarayana (1958) describing a testa with the outermost ca 6 layers consisting of thickened cells, which one normally would not expect to find in a drupe (there the testa is often reduced), while Grey-Wilson (1980a) described the fruit as being a drupe with separate stones. The micropylar endosperm haustorium is massive and may invade the funicle and even the placenta.
For general information, see Sandt (1921), Grey-Wilson (1980c: African taxa), Fischer (2004a), Leins and Erbar (2010), S.-X. Yu et al. (2015: esp. Impatiens sections) and Suksathan and Ruchisansakun (2020: Thai taxa), for chemistry, see Szewczyk (2018), for lamina epidermis, see X.-X. Zhang et al. (2013), for information on floral anatomy, development, etc., see Grey-Wilson (1980b), for the gynoecium, see Shimizu and Takao (1982, 1985), for ovule variation and seed anatomy and development, see Guignard (1893), Chandresekhara Naidu (1985) and Boesewinkel and Bouman (1991), for some embryology, see Dahlgren (1934a) and Narayana (1963), for seed morphology and classification, see Y. Song et al. (2004) and Utami and Shimizu (2005) - variation here is considerable, and for germination, see Hofmann and Ludewig (1985).
Phylogeny. Hydrocera and Impatiens are sister taxa (Yuan et al. 2004; esp. Janssens et al. 2006a: atpB-rbcL), etc.. Taxa of Impatiens with three sepals are scattered through the genus, so that condition is apparently at least sometimes derived (Janssens et al. 2006a); see also Janssens et al. (2012a: I. omeiana sister to the rest) and especially S.-X. Yu et al. (2015: ITS, 2 plastid genes, 46 morphological characters) for relationships [C [[R + S] [[F + I], T [S + U]]]]. Ruchisansakun et al. (2015) explored the limits of section Semeiocardium. The monophyly of the spurless section Trimorphopetalum (now included in sect. Uniflorae) from Madagascar is strongly supported, and Impatiens from Madagascar plus the Comoro islands also forms a monophyletic group (Rahelivololona et al. 2018: outgroup sampling limited). H. Qiu et al. (2023: 26 taxa, 80 protein-coding chloroplast genes) recovered the two subgenera mentioned below, all four sections in the analysis with two or more species were monophyletic, and the relationships in Yu et al. (2015) were largely recovered as well - [C [[R + S] [I [F [S + U]]]]]. Y.-X. Song et al. (2022: 189 sequences, two markers, ITS and atpB-rbcL) recovered the relationships [Clavicarpa (subgenus) [W[Semeiocardium + SishRacemosae] [Fasciculatae Sish[Tuberosae W[Impatiens [Scorpioidae + Uniflorae]]]]]] (all sections) (W = weak, Sish = strongish support, other nodes all strong), overall rather similar to Yu et al. (2015). F. Quin et al. (2023) thought that the sister group of section Impatiens was [Fasciculatae [Scorpioidae + Uniflorae]] (age of common ancestor (21.2-)16.2(-10.8) Ma). X. Zhang et al. (2025) included 121 ITS, 95 atpB-rbcL and 76 plastome sequences, although the last seemed to have little to do with their character optimizations, and although the ITS tree played a major role there, support along the backbone of that tree was rather weak at all the nodes, one node being collapsed for the character analyses. Y.-F. Liang et al. (2025: 139 spp., all major clades, plastomes and mitogenomes) found that the same sections and subgenera were recovered in both plastid and mitochondrial analyses, but the relationships between the sections differed somewhat between the two sets of analyses, and even between different analyses of the plastomes - concordant quartet support values for these relationships tended to be (very) low. There was also an interesting clade, Longlinensis, in the Semeiocardium-Racemosae area (Liang et al. 2025).
Section Impatiens. F. Qin et al. (2023) looked at plastome variation of 82/ca 90 species of this section, recovering a number of well supported clades; the basal split in the section was between a clade centred on Southeast China and another, including the majority of the section, that was centred on the Hengduan Mountains.
Section Racemosae. T. Xue et al. (2025: 52/70 spp., plastomes, nuclear ribosomal DNA) exmained relationships here - all told some 87 species. Support for relationships in the plastome tree was quite strong, that in the nuclear tree disagreed quite extensively within the two main plastome clades recovered (in fact there were four main clades in the plastome tree, five in the nuclear tree, one of the main clades being paraphyletic). However, support values in the nuclear tree were low (Xue et al. 2025); one awaits the addition of more nuclear data.
Classification. The old infrageneric classification of Impatiens needed complete overhaul (Janssens et al. 2006a). S.-X. Yu et al. (2015: two subgenera, seven sections) suggested a new classification, but it, too, clearly has its limits (see e.g. Ruchisansakun et al. 2015). Two subgenera and 7 sections plus 1 unnamed clade were consistently recovered by Y. F. Liang et al. (2025) and the classification there has largely been followed above.
Previous Relationships. Balsaminaceae were included in Geraniales-Rosidae by Cronquist (1981; see also Takhtajan 1997).
TETRAMERISTACEAE Hutchinson - Back to Ericales
Evergreen trees; chemistry?; intervessel pitting opposite-alternate; petioles short [1> cm]; bracteoles rather large, ± caducous; fruit indehiscent; ?endosperm development.
3/5: [list], two groups below. W. Malesia, Central and N. South America.
Age. K. Bremer et al. (2004a) estimated the age of crown-group Tetrameristaceae to be 41 Ma; (31-)28, 25(-22) Ma was the age suggested by Wikström et al. (2001) and (53-)31(-13)Ma by Wikström et al. (2015) - other estimates, (42-)30, 28(-15) Ma (Bell et al. 2010) and ca 44.6 Ma (Rose et al. 2018) - but see below for Pelliciera.
Chemistry, Morphology, etc.. The lamina narrows gradually towards the base, and any petiole can be difficult to make out - it is at best short.
1. Pelliciereae Triana & Planchon - Pelliciera rhizophorae Planchon & Triana —— Synonymy: Pellicieraceae Bullock
Fluted buttresses made up of "adventitious" roots; vessels in multiples; petiole bundle more or less flat where it joins the stem, becoming annular; stomata cyclocytic; lamina vernation involute, base asymmetric, with paired glands abaxially, colleters?; flowers terminal, single, ca 10 cm across ["very large"]; bracteoles C-like; K with two traces at base, quincuncial; A extrorse, anthers very long [>4.5 cm long], connective prolonged into a point; pollen strongly verrucate; G apparently 2-carpelate, style long, stigma bifid, punctate; ovule 1/carpel, apical, pendulous, campylotropous; fruit ± dry, pointed, pericarp corky, K and C caducous; seed large, coat 0; endosperm 0, cotyledons large; n = 17, nuclear genome size [2 Cx] 2.52-2.56 pg; ± viviparous [embryo breaking through the pericarp while fruit still on the plant], germination phanerocotylar, hypogeal.
1/1 (?2). Central and N. South America, on both the Caribbean and Pacific shores. Map: see A. Graham (1977), also Duke (2020: Map 1) and Rull (2023: Fig 14A). Photo - Flower, Fruit.
Age. Z. He et al. (2022: Fig. 2b) date Pelliciera at over 60 Ma while Rull (2022, 2023) suggests an age of ca 45 Ma, some time in the mid-Eocene.
2. Tetrameristeae Hallier
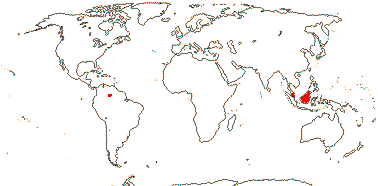
Cork inner cortical; (vessel elements with scalariform perforation plates); wood fluorescing [1 sp. tested]; nodes 3:3; stone cells [in stem] +, branched sclereids ?0; (bracteoles persistent); flowers 4- or 5-merous, rather small; filaments slightly connate at the base; G [(4)]; ovule basal, ?epitropous; fruit a berry; testa several layers thick, walls thickened; endosperm copious, cotyledons small; n = ?
2/4. Malesia (Tetramerista), Venezuelan Guyana (Pentamerista).
Evolution: Divergence & Distribution. Records of fossil pollen (see Lanagiopollis crassa, = Psilatricolproyes crassus) suggest that Pelliciera was once more widespread (A. Graham 1977; Rull 2023), and it may even have grown in the Old World. However, the fossil history of the genus is uncertain. Martínez-Millán (2010) did not accept New World records of the genus, which are based on pollen; Old World records are also based on pollen (see also Ellison et al. 1999; Plaziat et al. 2001). Manchester et al. (2015) thought that pollen of Alangium (Cornaceae) could be confused with that of Pelliciera (see also Duke 2020 for possible confusion with Hura-Euphorbiaceae). On the other hand, Rull (2022, 2023a, b: Fig. 3) based his detailed scenario of changes in the range and abundance of Pelliciera in the New World from the Eocene onwards on the palynological record. He thought that the genus predated most other mangroves there (but not Nypa), and in the Eocene it was abundant where it grew. However, after the Eocene-Oligocene climate transition (cooling, pCO2 decrease, etc.) some 34 Ma Rhizophora in particular became widespread and abundant while Pelliciera became very much less abundant, if somewhat more widespread than it had been, and since then its range has contracted and shifted - Nypa became extinct (see especially Rull 2023a). Rull (2023a) even suggests that Pelliciera might even do better growing with Rhizophora since the former does not tolerate high light and high salinity conditions, the latter being generally more tolerant, although why Pelliciera apparently flourished in the Eocene then becomes unclear (to me, at least).
Ecology & Physiology. For the evolution of the mangrove habitat, to which Pelliciera is restricted, see Rhizophoraceae.
Pollination Biology & Seed Dispersal. The fruits of Pelliciera can float for about a week, and may lose their pericarps as they are floating, germination beginning then; the seeds retain their viability for up to 70 days (Rabinowitz 1978; Rull 2023).
Genes & Genomes. For the cytology, etc., of Pelliciera, see Garzón-Bautista et al. (2018).
Chemistry, Morphology, etc.. There are no reports that Pelliciera accumulates aluminium, unlike Theaceae s.l., in which it has often been included. The nodal anatomy of Tetrameristeae is extrapolated from their petiole scars. The products of different marginal glands of the one leaf there may not be the same (Collins et al. 1977).
The floral diagram of Pelliciera in Tomlinson (1986) suggests that either the two carpels are oblique, or the bracteoles are not in the lateral position and the carpels are transverse. In Tetramerista there are glistening dots on the adaxial surface of both calyx and corolla. Although the gynoecium of Pelliciera appears to be bicarpelate at maturity, the stylar canal is five-radiate and so the gynoecium may be basically 5-carpelate (Schönenberger et al. 2010).
For general information, see Kubitzki (2004b: Tetrameristeae) and Kobuski (1951), Tomlinson (1986), Maas and Westra (1993), Kubitzki (2004b) and Duke (2020), all Pelliciera. For some vegetative anatomy, see Beauvisage (1920: Pelliciera).
The embryology, morphology and anatomy of Pellicieraceae s.l. are poorly known.
Previous Relationships. Pellicieraceae s. str. and Tetrameristaceae s. str. were included in Theales by Cronquist (1981) and Takhtajan (1997).
[[Polemoniaceae + Fouquieriaceae], Lecythidaceae, [Sladeniaceae + Pentaphylacaceae], Sapotaceae, Ebenaceae, Primulaceae, Mitrastemonaceae, [Theaceae [Symplocaceae [Styracaceae + Diapensiaceae]], [Sarraceniaceae [Roridulaceae + Actinidiaceae] [Clethraceae [Cyrillaceae + Ericaceae]]]]]: corolla connate, tube well developed; style long; Adβ/ACCHβ genome duplication?
Age. This node may be (95-)85, 82(-72) Ma in age (Bell et al. 2010), around 99.7 Ma (Magallón et al. 2015), (106-)99(-94) Ma (Wikström et al. 2015) - note topologies in all.
Genes & Genomes. The Ad-β/ACCH-β/Cm-α genome duplication event, dated at ca 85.6 Ma, may be placed at this node (Landis et al. 2018: ?Lecythidaceae; Larson et al. 2019/2020). This Ad-β event has also been dated to ca 75.9 or 101.4-72.9 Ma (Shi et al. 2010 and S. Huang et al. 2013 respectively); see also Soza et al. (2019) who suggested that it happened ca 85.6 Ma, C. Zhang et al. (2020) estimated an age of ca 108.6 Ma, while Q. Zhang et al. (2022: [Pentap. + Diap. + Symp. + Styr.]) suggested an age of (102.5-)99.7(-92.0) Ma - see also Theaceae, etc. below. For the older At-γ event mentioned by Zhang et al. (2022), see goings on at the base of the eudicots.
[Polemoniaceae + Fouquieriaceae]: kaempferol, quercetin +; cork cambium outer cortical; inflorescences terminal, determinate; K with scarious margins; A adnate to the corolla, thecal septum at most short and indistinct [septum 0]; gynoecial nectary +, with stomata; G [3], style hollow, style 3-branched; ovules in two ranks, apotropous, micropyle zig-zag [abruptly turned towards central axis]; fruit a loculicidal capsule, seeds on central columella; seeds winged; exotesta with circular/annular thickenings; endosperm scanty; mitochondrial coxII.i3 intron 0.
Age. The age of this node is estimated at (98-)51(-50)Ma by Wikström et al. (2015), (77-)65, 61(-48)Ma by Bell et al. (2010), ca 79.6Ma by Tank et al. (2015: Table S2), around 83Ma by Magallón et al. (2015), ca 99.4 Ma by Rose et al. (2018) and (95.8-)75.5(-60.3) Ma (De-Nova et al. 2018).
Evolution: Divergence & Distribution. Schönenberger (2006a, especially 2009) lists many other features occurring in this family pair, including free sepals, stomata on the abaxial surface of the calyx (also e.g. Ericaceae - ?general distribution of this feature?), anastomosing vascular bundles in sepals and petals, and details of gynoecial development. Chartier et al. (2017) commented that these two families were rather different, at least based on the 37 floral features that they examined.
Chemistry, Morphology, etc.. Both families have late corolla tube formation (Schönenberger 2009); for general floral morphology, see Schönenberger et al. (2010).
POLEMONIACEAE Jussieu, nom. cons. - Back to Ericales
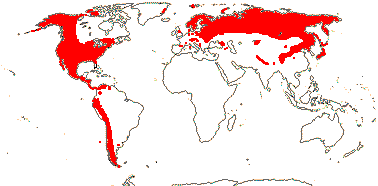
Fructan sugars accumulated as kestose and isokestose oligosaccharides [levans and inulins], cucurbitacins +, ellagic acid 0; cork cambium also pericyclic; (vessel elements with scalariform perforation plates); (wood rayless); (stomata paracytic); leaves opposite to spiral, lamina conduplicate, margins entire to deeply lobed; bracteoles 0; K connate, aestivation open, lobes with green midrib and colorless intermediate portion, tips terete/spine-like, C lobes usu. right-contortuplicate; K/C tube well developed, stamens = and opposite sepals, inserted at different levels or filaments of different lengths, anthers ventrifixed (basifixed); pollen pantoporate; nectary usu. not vascularized, prominent; G [(2-4)], median member adaxial, placentae protruding, stigma dry, the length of the arms; ovules 1-many/carpel, vascular bundle not reaching chalaza; seeds often mucilaginous when wetted, exotesta variously thickened, endotesta a pigment layer, radial walls ± thickened; endosperm nuclear/coenocytic, haustoria 0, embryo chlorophyllous or not; x = 9, nuclear genome [1C] (0.145-)3.011(-62.581) pg.
18/375: [list, to subfamilies] - three subfamilies below. N. temperate, W. North America, South America. Map: from Hultén (1971) and Meusel et al. (1978).
Crown-group Polemoniaceae are estimated to be (40-)36, 33(-29) Ma (Wikström et al. 2001), (47-)36, 31(-20) Ma (Bell et al. 2010) or ca 53.9 Ma (Rose et al. 2018: Acanth sister).
1. Polemonioideae Arnott
Herbs (annuals), (subshrubs); (short shoots +); leaves compound to simple; (inflorescence capitate); (flowers monosymmetric, median C ab- or adaxial); C veins usu. free or connected well above the base, (lobes with teeth); filaments usu. merged with corolla; (pollen 6-9 colporate); integument (3)7-20 cells across, hypostase +; seeds not winged (narrowly winged - Loeselia), (testa ± multiplicative); n = (6) 7 (8) 9, chromosomes "larger" [not Loeselia]; also sporophytic incompatibility system present.
13-22/340: Phlox (70), Linanthus (26), Navarretia (30), Polemonium (27), Gilia (25). Especially western North America, also a few N. temperate, southern South America. Several predominantly western North American genera have a few species in southern South America (see below). Photos: [Collection (all except Cobaea).]
Age. Gilisenium hueberi, perhaps close to Gilia, is known from deposits from the middle Eocene of Utah ca 40.4 Ma (Lott et al. 1998; Martínez-Millán 2010).
2. Cobaeoideae Arnott —— Synonymy: Cobaeaceae D. Don
Vines with leaf tendrils to small trees (herbs); (short shoots +): stem with endodermis [Cobaea]; leaves usu. unequal-pinnate (palmate, simple); flowers large, (mostly positionally monosymmetric - C.); K (basally connate), usu. herbaceous throughout, C veins connected at the base of lobe (and in upper lobe); A traces in two whorls, filaments often superficially adnate to C; pollen (exine verrucate), (100-220 µm long); ovules with nucellar cap; fruit septicidal and/or loculicidal; seed wing broad (narrow - Bonplandia); mesotestal cell walls thinly lignified; n = 15, 26, 27, [x = 7-9?] chromosomes "small".
4/34: Cobaea (18). Baja California, tropical America. [Photo - Flower & Fruits.]
3. Acanthogilioideae J. M. Porter & L. A. Johnson - Acanthogilia gloriosa (Brandegee) A. Day & Moran
Shrubby; short shoots +, leaves very dimorphic, spiny [persistent, branched on long shoots, unbranched on short shoots; pollen ≥4 equatorially colporate, exine coarsely verrucate; seeds few; n = 9, chromosomes 2-4 µm long.
1/1. Baja California.
Evolution: Divergence & Distribution. There are some 15 species of Polemoniaceae-Polemonioideae in western South America that have their closest relatives in North America; these disjunctions are up to 19.5 Ma and may well be the result of the mucilaginous seeds of the family sticking to migratory birds (L. A. Johnson & Porter 2017). Details of speciation patterns vary, but in no cases have there been extensive radiations in South America, in a few cases the South American representatives do not even represent distinct species, or they may be allopolyploids, hybridisation happening in South America or in North America, but in both cases with one or both parents extinct, and so on - very complex scenarios are possible in the examples discussed (Jonson et al. 2012; Johnson & Porter 2017). There are also clusters of similar events in Poaceae (Peterson et al. 2010b) and Boraginaceae (Guilliams et al. 2017).
Linanthus and some other genera show extensive infraspecific colour polymorphisms; Anghel et al. (2025) looked at relationships here and suggested that a clade of perennials was derived – rather unusual. They recovered a night-blooming (moth-pollinated) clade (night flowering was polyphyletic), and there is a variety of other pollinators here; polymorphism in flower colur is extensive. They found that at least 14 species co-occurred and co-flowered in the deserts of the S.W. U. S. A. (Anghel et al. 2025).
Schönenberger (2009) lists additional features that may be apomorphies for Polemoniaceae.
Ecology & Physiology. Polemoniaceae are well represented in drier habitats in western North America in particular.
Pollination Biology & Seed Dispersal. For floral variation in the context of different pollinators, see the classic work of Grant and Grant (1965). The pollen in bat-pollinated taxa tends to notably large (T. N. Taylor & Levin 1975), as has been noted elsewhere (Stroo 2000).
De Groot (2011) found remarkable infraspecific variation in floral orientation in Eriastrum eremicum (Polemoniaceae), and the flowers in this species also show considerable variation in their lobing (5:0, 3:2, 2:3, etc.); in both Eriastrum and Ipomopsis the median petal may be ad- or abaxial. Heterostyly is reported in Aliciella (Cohen 2019).
A number of Polemonioideae have myxospermous seeds (L. A. Johnson et al. 2004; Winter 2012).
Plant-Animal Interactions. For Polemoniaceae with glandular hairs that may trap insects on those hairs, see LoPresti et al. (2015), also elsewhere.
Genes & Genomes. For a genome duplication somewhere in the family, see Larson et al. (2019/2020).
Chemistry, Morphology, etc.. The cambium is sometimes storied; raylessness is frequent. In Cobaea the leaves are tendrillar and the basal pair of leaflets is foliaceous-stipuliform; development of the compound leaves of Polemonium is basipetal (Hagemann & Gleissberg 1996).
For inflorescence morphology, see Weberling (1989), for floral development, see Schönenberger (2009), for pollen, see Monfils and Prather (2004, and references), for some embryology, see Kapil et al. (1969), for seed morphology, see L. A. Johnson et al. (2004), for general information, see Grant (1959, 1998), Day and Moran (1986: esp. Acanthogilia), Johnson et al. (1996, 1999), Porter (1997), Porter and Johnson (1998) and Wilken (2004).
Phylogeny. Acanthogilia has been placed in its own subfamily (Porter et al. 2000), and it may be sister to the Cobaea et al. clade (Prather et al. 2000) or even sister to the whole of the rest of the family (Schönenberger et al. 2005, only four taxa included) - its position is unclear (L. A. Johnson et al. 2008 and references), although Rose et al. (2018) also found it to be sister to the rest of the family. It has very dimorphic leaves and short shoots and in this is like Fouquieraceae, its branched spines are reduced leaves like those found scattered in Polemonioideae, and its sepals have a green midrib, as in Cobaeoideae.
Johnson et al. (2008) suggest the following relationships within Polemonioideae - [[Polemonieae (one genus) + Phlocidae] [Gilieae + Loselieae]]. For the phylogeny of Phlox, see Ferguson et al. (2008). Relationships recovered in Polemonium depend on what part of the genome is being examined (Rose et al. 2020). For relationships in Linanthus, see Anghel et al. (2025; 25/26 spp., Angiosperms353 and double digest restriction-site associated DNA); topologies obtained from the two sets of analyses were very largely similar and well supported. Anghel et al. (2025) included two or more specimens/species and found that some of the species in the night-blooming clade and in the perennial group were not monophyletic.
Classification. Porter and Johnson (2000) suggest a phylogenetic classification for the family. The limits of the genera Ipomopsis (Porter et al. 2010) and Gilia (Prather et al. 2000; L. A. Johnson et al. 2008) need to be redrawn.
Previous Relationships. Polemoniaceae were included as Polemoniales in Solananae by Takhtajan (1997).
FOUQUIERIACEAE Candolle, nom. cons. - Fouquieria Kunth - Back to Ericales
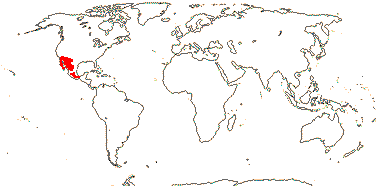
Woody, xeromorphic, with long and short shoots; flavonols only, ellagic acid, route I secoiridoids +, myricetin 0; root cork cambium outer cortical; stem cortex often with fibrous ridges, etc; cork cambium underneath; cuticle wax crystalloids 0; leaves heteromorphic, lamina (isobilateral), margins entire, petiolar spines on long shoots; K separate, ± scarious, imbricate; A 10(-23), only very shortly adnate to corolla; pollen with oil bodies; nectary tissue in base of ovary, vascularized; placentation intrusive parietal most of the length of the ovary, style with long branches, stigmas punctate (subcapitate); ovules 3-20/carpel, apotropous, bitegmic, micropyle endostomal, outer integument 3-4 cells across, inner integument (3-)7-8 cells across, suprachalazal zone massive; embryo sac tetrasporic, with lateral haustorium; (suspensor long, thin); testa and tegmen multiplicative, becoming crushed, testa hypodermis with banded thickenings; endosperm with micropylar and chalazal haustorium; n = 8, 12, x = 6 (7, 8), genome size (0.049-)1.358(-37.569) pg.
1/11: [list]. S.W. North America. Map: from Henrickson (1972b). [Photos - Habit, Branch, Flowers.]
Age. Diversification began here (20.9-)12.7(-7.8) Ma (De-Nova et al. 2018) or ca 11.2 Ma (Bell et al. 2018).
Evolution: Divergence & Distribution. Diversification began at the Miocene-Pliocene boundary; there is a ca 60 Ma stem, and molecular evolution in Fouquieraceae has been around twice that in Polemoniaceae (De-Nova et al. 2018). Soto-Trejo et al. (2022) also discussed the biogeography and evolution of the family; they thought that the more succulent taxa were plesiomorphic, the woody taxa derived. Schönenberger (2009) lists other features that may be apomorphies for Fouquieriaceae, including sepals and corolla lobes being similar in size and histology - but they are not notably similar in size...
Genes & Genomes. Soto-Trejo et al. (2022) suggested that the base chromosome number for Fouquieriaceae was n = 12.
Chemistry, Morphology, etc.. Layers of fibrous cells alternate with layers of cork cells in the stem cork, while the cork cambium in the root is described as being superficial (Henrickson 1969), the unusual position for angiosperms, although perhaps commoner in desert plants.
For an asymmetric phase in early floral development, see Schönenberger (2009); this may be connected with the fact that the perianth parts are borne in a distinct spiral. Members of the androecium have been described as being borne in a single whorl, but they are diplostemonous, and the antepetalous stamens are doubled in some species (Schönenberger 2009; Schönenberger & Grenhagen 2005). There is quite a lot of variation in the development of the embryo sac, but it always seems to be tetrasporic (Johansen 1936).
See Henrickson (1972b) and Kubitzki (2004b) for general information, for some root and stem anatomy, see Henrickson (1936a), for pollen, see Henrickson (1973) and Grigorjeva and Gabarayeva (2018), for ovule morphology and embryology, see Mauritzon (1936b), and for seeds, see Corner (1976).
Phylogeny. For relationships within the family, see Schultheis and Baldwin (1999), De-Nova et al. (2018) and Soto-Trejo et al. (2022).
Previous Relationships. Fouquieriaceae were placed in Violales by Cronquist (1981).
LECYTHIDACEAE A. Richard, nom. cons. - Back to Ericales
Trees (lianes); flavonols, ellagic acid +, kaempferol 0; (vessel elements with scalariform perforation plates); cortical vascular bundles +; (wood siliceous and/or with SiO2 grains); phloem stratified (with wedge-shaped rays); nodes 3 or more:3 or more; petiole with numerous arcuate or annular bundles in arcs, etc.; stomata usu. anisocytic; lamina margins short-toothed or entire, (tertiary veins subparallel, ± at right angles to midrib), stipules cauline, small or 0, colleters +; pedicels articulated; flowers large; K various; A latrorse, filaments not articulated, basally adnate to C, outer A staminodial; tapetum amoeboid, cells binucleate; pollen grains tricolpate/tricolporate, tricellular; nectary +; ovary inferior, [2-8], stigma ± capitate, wet or dry; ovules 1-many/carpel, bitegmic, micropyle endostomal, micropyle long [longer than embryo sac + chalaza]; antipodals ephemeral; K persistent in fruit; seeds often arillate, testa ?multiplicative, vascularized, exotestal cells variously thickened, palisade, or low with sinuous anticlinal walls, mesotesta sclerotic or not; endosperm nuclear/coenocytic, 0; x = 17/18 (?9); mitochondrial coxII.i3 intron 0 [?sampling].
31/348: [list: to subfamilies] - five subfamilies below. Tropical, especially America and W. Africa.
Age. Crown-group diversification in Lecythidaceae may have begun (65-)84, 46(-30) Ma (Bell et al. 2010), ca 82.9 Ma Rose et al. (2018), or (71-)65, 61(-55) Ma (Wikström et al. 2001).

1. Napoleonaeoideae (A. Richard) Bentham - Napoleonaea P. de Beauvois —— Synonymy: Belvisiaceae R. Brown, Napoleonaeaceae A. Richard
(Twigs green, winged); flowers single, axillary, ± sessile, "bracts" several; C 30-40 segments, connate, plicate, margin serrate [see discussion].
2/21.
1A. Napoleona P. de Beauvois —— Synonymy: Belvisiaceae R. Brown, Napoleonaeaceae A. Richard
Secondary xylem with crystal chains; ?stomata; leaves 2-ranked, lamina supervolute, with glands abaxially {submarginal; near base), margin ± serrate, stipules +/0; (inflorescence axis, pedicels, etc. well developed N. egertonii), (plant cauliflorous); G initiated before A; K 5, valvate, usu. with marginal nectariferous glands; 3 whorls of staminodia, members of outermost whorl many, subulate, free, ± horizointal-recurved, members of next whorl many, ± erect, connate except at apex, fertile A 10, paired, incurved, opposite C, extrorse, monothecal, alternating with pairs of incurved staminodes, all 20 members forming a single ring, connate except for windows at the midpoint; tapetal cells binucleate; nectary annular, at base of style; G opposite C, style short, very stout, stigma broad, pentagonal, flat; ovules 4/carpel, apical, collateral in pairs, outer integument 5-7 cells across, inner integument 9-11 cells across, integuments basally connate, ?micropyle length, endothelium 0; fruit a 1-4-seeded drupe; testal bundle single; endosperm 0; embryo curved; n = 16.
1/17. W. tropical and Central Africa. Map: from Liben (1971b) and Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003). Photo:
1B. Crateranthus Baker f.
K 3, imbricate; A with outer whorl staminodial, forming pseudo corolla; A ∞, bilocular, filaments slender; G semisuperior, [3-5], placentation axile-apical; style slender, stigma linear, on upper half of style; ovules "numerous"/carpel; fruit baccate, germination epigeal; n = ?
1/4. S.E. Nigeria to Gabon.
[Asteranthoideae [Lecythidoideae [Barringtonioideae + Foetidioideae]]]: A many [15-1200], initiated as ring primordium, development centrifugal (centripetal), in concentric series or not, basally connate; G opposite sepals.
Age. The age of this clade is ca 64.8 Ma (Rose et al. 2018).
2. Asteranthoideae Reveal (the old Scytopetaloideae) —— Synonymy: Asteranthaceae R. Knuth, nom. cons., Rhaptopetalaceae van Tieghem, Scytopetalaceae Engler, nom. cons.
Growth ?sympodial; plants Al-accumulators; nodes?, bundles leave stele two internodes before entering leaves; (cristarque cells +), sclereids +; ?crystal chains, crystals octahedral; leaves amphistomatous); leaves 2-ranked, stipules +, minute; inflorescence axillary, branched to 1-flowered, axis terminated by a flower; (pedicels not articulated); K connate, ([2-)3-lobed, imbricate), C [?= staminodes] thick, connate, (splitting into 6-16 segments), (thin, plicate, margins serrate - Asteranthos, Crateranthus [C.]); (anthers longer than filaments), (dehiscing by apical slit), (connective produced), (endothecium encircling whole anther - C.); tapetum glandular; (pollen tricolporoidate); (nectary 0); G superior (half-superior), style relatively long, slender, stigma punctate to lobed; ovules 2-4/carpel, (apical, campylotropous - C.), outer integument 5-8 (16-18 - C.) cells across, inner integument 5-9 cells across, (supra-chalazal zone long, narrow); fruit indehiscent (loculicidal capsule - Oubanguia); seeds usu. 1, with unicellular hairs or not, ruminate or not; testa cells often with crystals, testal bundle single or branched; endosperm +, ruminate (not Oubanguia), walls irregularly thickened, hemicellulosic, (embryo J-shaped), cotyledons accumbent, at least half the length of the embryo (ca 1/4 - Asteranthos); n = 11, 18, 21.
7/27: Rhaptopetalum (12), Scytopetalum (4). West tropical Africa, Asteranthos brasiliensis upper Rio Negro and Orinoco in Venezuela, Brazil, Colombia. Map: See Prance and Mori 1979), Heywood (1978) and Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003). [Photo - Fruit.]
[Lecythidoideae [Barringtonioideae + Foetidioideae]]: endosperm 0, embryo usually starchy.
Age. The age of this clade is ca 59.9 Ma (Rose et al. 2018) or (68.9-)62.7(-56.7) Ma (Vargas & Dick 2020).
3. Lecythidoideae Beilschmied —— Synonymy: Gustaviaceae Burnett
Secondary xylem with crystal chains; (colleters +); leaves two-ranked or spiral, lamina vernation involute; (inflorescence cauliflorous); (flowers monosymmetric via connate androecium); C (4-5) 6; A incurved, ?adnate to C, filaments contracted at the apex; pollen tricolp/oroidate, (fodder pollen +); nectary 0/+; G [2-8] carpelate, style short (long), stigma (lobed - Grias); ovules 3-many/carpel, (pendulous/erect), outer integument 5-25 cells across, inner integument 3-8 cells across, (outer integument with micropylar arilloid), (integuments basally connate), (micropyle short), outer epidermis 9-16 cells across, inner epidermis 5-9 cells across, (endothelium 0); fruit operculate (indehiscent); seed with swollen funicle, or aril (= wing), or neither, testal bundles (1-)2<; (endosperm sparse - Grias), (ruminate - Allantoma), embryo curved or not, macropodial [hypocotylar, cotyledons ± 0] or with long radicle and leaf-like (folded) or fat cotyledons; n = 17 (18); germination epigeal or hypogeal.
16/230: Gustavia (40), Scottmoria (23). Neotropical. Map, blue: from Prance and Mori (1979) and Mori and Prance (1990a). Photos: Flower, Fruit, Flower, Fruits.
Age. The estimated age of this clade is (55.9-)46.1(-36.5) Ma (Vargas & Dick 2020).
[Barringtonioideae + Foetidioideae]: cortical vascular bundles with inverted orientation; nodes 1:1; leaves supervolute; style slender, long; fruit indehiscent; seeds 1/fruit.
Age. This clade is ca 29.5 Ma (Rose et al. 2018).
4. Barringtonioideae Beilschmied —— Synonymy: Barringtoniaceae F. Rudolphi, nom. cons.
Secondary xylem without crystal chains, axial parenchyma diffuse-in-aggregates; leaves spiral, vernation?, glands in the stipular position; inflorescence racemose or paniculaate (flowers single); K 2-6, imbricate ofter circumscissile; C 3-5.imbricate; A inner whorls often staminodial; pollen syntricolp(or)ate, strong marginal ridge [thickened mesocarpial margin along colpi], polar cusion [thickening at polar junction of margins of adjacent colpi]; nectary annular; stigma punctate; ovules 1-many/carpel, (pendulous), (basal), endothelium 0 (+); seeds 1-5/fruit (many - Planchonia); (arilloid + [apex of outer integument] - Careya); testal bundles ≥2; endosperm at most scanty, embryo hypocotylar/macropodial, (radicle long, coiled), (cotyledons plicate - Planchonia), (embryo undifferentiated - Careya); n = 13; nuclear genome [1C] ca 1319 Mb; germination epigeal/hypogeal/hypogeal, with first leaves reduced.
6/91: Barringtonia (73). Paleotropical. Map above, Old World only, red: from van Steenis and van Balgooy (1966), Payens (1967), Liben (1971b) and Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003). [Photo - Flower]
Age. Wood that has been refered to Barringtonia or Petersianthus is reported from the Late Cretaceous/Early Palaeocene Deccan Traps (Manchester et al. 2015 and references; Wheeler et al. 2017).
5. Foetidioideae Niedenzu - Foetidia Lamarck —— Synonymy: Foetidiaceae Airy Shaw
Secondary xylem with crystal chains; leaves elongating in bud, petiole inconspicuous; K woody, C 0; A free, ∞, introrse, staminodes 0; nectary indistinct; style 3-5-fid; ovules in two rows, 5-30/loculus, anatropous to ± campylotropous, integument largely connate; testal bundles 4-5; K persistent, 1-2 seeds/loculus; ?cotyledons; n = ?
1/18. Madagascar, also E. Africa (Tanzania, Pemba), the Comores, Mauritius and Réunion.
Evolution: Divergence & Distribution. Note that there have been changes to the names of a number of South American Lecythidoideae - see Classification below. The discussion below does not reflect all these changes (for which, see Vargas et al. 2024).
For additional ages within the family, see Rose et al. (2018) and Vargas and Dick (2020).
Mori et al. (2017) discussed the biogeography of Lecythidaceae in some detail. Vargas and Dick (2020) suggested that the subfamiy originated in western Amazonia, but that the Guayana area has been important for diversification of the Bertholletia clade in particular; overall, speciation rates in American Lecythidoideae have held steady over the course of their existence. Chave et al. (2024) suggested that Miocene changes associated with the Andean uplift drove substantial and quite recent (124 species in some 10 million years) diversification in the clade including Bertholletia and its sisters; interestingly, Hoorn et al. (2023) depicted diversity in Eschweilera as being flat since the later Pliocene.
The [Allantoma + Cariniana decandra] clade (= Allantoma s.l., Lecythidoideae, see Y.-Y. Huang et al. 2008; Mori et al. 2007) may be an example of the derivation of polysymmetric flowers from a monosymmetric ancestor (there are many other examples in the gentianids), but the current phylogeny does not yet provide strong support for this. However, there is strong support for the hypothesis that the flowers of Lecythidoideae were initially polysymmetric, even if most extant species are monosymmetric (Mori et al. 2007).
Chartier et al. (2017, see also 2021) commented on the extent of androecial variation in Lecythidaceae compared to that in other ericalean families, even if overall there is great variation elsewhere in the order, and things get tricky when comparing floral morphology within Lecythidaceae. Thus in Napoleonoideae there are sepals, and then a large petaloid urceolate-tubular to flat-discoid structure with toothed margins; inCrateranthus this is called a pseudocorolla that is formed from connate stamens while in Napoleona it is called a corolla, and it is described as having 30-40 fused segments which are plicate; within this structure are numerous stamens, or three whorls of staminodes and ten functional stamens (Prance & Jongkind 2015). My immediate reaction when reading the descriptions, etc., was that the outer petaloid structure was the "same" in the two genera (see characterizations above), but developmental work is sorely needed here. Indeed, in general there is much floral variation, as is also evident e.g. when reading the descriptions of seeds and fruits (Prance & Mori 1978, 1979; Vargas et al. 2024: the Bertholletia clade).
Ecology & Physiology. 19 of the 107 species of Lecythidaceae-Lecythidoideae from Amazonian forests are in the 227 species that make up half the stems of trees ≥10 cm across, so are disproportionally well represented (almost 8.4% of the total) in those forests. They are second in terms of numbers of these species, although they are quite a small group, they are third in terms of numbers of individuals, and Eschweilera is the most abundant tree genus, E. coriacea the most abundant species (ter Steege et al. 2013: Fabaceae #1, Sapotaceae #2; see also Vargas & Dick 2020; Chave et al. 2024; c.f. in part Levis et al. 2017; Maezumi et al. 2018). 3 species of Lecythidoideae are in the top 20 in terms of above-ground woody biomass, where they make up 3.96% of the total (Fauset et al. 2015: Fabaceae are #1). Across the Atlantic, in eight Central African rainforests sampled, Oubanguia (Scytopetaloideae) and Petersianthus (Lecythidoideae) are described as hyperdominant species (Bastin et al. 2015); they are two of the 18 species that made up 50% of the above-ground biomass, both with ca 1.8% biomass, #7 and #10 on the list.
Zona and Christenhusz (2015) note that a number of Lecythidoideae are little-branched litter-collecting Schopfbaum.
Pollination Biology & Seed Dispersal. Monosymmetric Lecythidoideae are pollinated largely by euglossine bees. Several taxa have fodder pollen produced by the anthers in the hood (sometimes by some of the anthers in the ring), and as a result the pollen is heteromorphic; fodder pollen may even be in tetrads, unlike the pollen of the ring stamens (Ormond et al. 1981); nectar is also found in some of these taxa (Prance & Mori 1979; Mori & Prance 1990a). These monosymmetric flowers are unlike those of any other angiosperm, with the monosymmetry primarily being evident in the massive development of the abaxial part of the staminal ring that leads to the production of the sometimes complexly coiled staminal hood into which the bees force their way. A rather close evolutionary association between euglossine bees and monosymmetric Lecythidoideae has been suggested (e.g. Mori & Boeke 1987); divergence of crown-group euglossines occurred some 42-27 Ma (Ramírez et al. 2010), that of the relevant Lecythidoideae...? Polysymmetric Lecythidoideae are pollinated by a variety of bees other than euglossines. Details of floral development, incuding the origin of monosymmetry, are to be found placed in a phylogenetic context in Tsou and Mori (2007). Interestingly, the polysymmetric Allantoma is embedded in the monosymmetric clade (e.g. Y.-Y. Huang et al. 2015).
The pollination and floral morphology of Napoleonaea vogelii has been described in detail (Frame & Durou 2001). Despite the size of the flower, pollination by thrips is suggested; there are nectaries inside the flowers at the bases of some of the staminodes and also on the outside of the calyx.
The seeds of Lecythidoideae are large and are probably mostly dispersed by mammals, especially primates.
Economic Importance. Mori and Prance (1990b) dicuss various aspects of the biology and economic importance of the Brazil nut, Bertholletia excelsa.
Chemistry, Morphology, etc.. Lecythidoideae have characteristically fibrous bark. Gustavia has inverted cortical bundles in the stem (Metcalfe & Chalk 1950). There is banded apotracheal parenchyma (c.f. Sapotaceae!) and crystals in the axial parenchyma, the latter common in several other Ericales, but wood anatomy suggests little about groupings within Lecythidaceae (but see Mori & Prance 1990a) and relationships of the family (c.f. Lens et al. 2007b). In both Barringtonioideae and Foetidioideae the nodal anatomy appears to be 3:3 if one looks only at the base of the petiole, but the nodes are 1:1 in a t.s. of the stem. Ditsch and Barthlott (1994) suggested that the rather dimorphic wax platelets of Asteranthos differ from those of Scytopetalaceae, but such platelets also occur in some species of Barringtonia (c.f. their figs 26, 27, 29), so are not out of place in Lecythidaceae. In at least some species of Barringtonia there are little glands in the stipular position; these are perhaps to be compared with the minute "stipules" of Asteranthoideae (for which, see Breteler 2002). Cariniana is reported to have colleters on the leaf margin whose exudate lubricates the expansion of the young leaves (Paiva 2012; see also Fernandes et al. 2016); are these glands of Barringtonia, the teeth of other Lecythidaceae (in Napoleonaea insignis the margin is entire, but it also has tiny black spots), and the stipules, also colleters...?
The Cariniana ianeirensis clade is described as having obliquely monosymmetric flowers (Mori et al. 2007, 2017; Huang et al. 2008), but this refers to the apex of the staminal tube, not the orientation of the whole flower with respect to the vertical axis. Androecial variation in Lecythidoideae is extreme - note that the erect and reflexed stamens mentioned by Mori et al. (2017: esp. Fig. 5) are here described as being incurved - and there is both centripetal and centrifugal androecial development (Tsou 1994). The carpels are shown as being opposite the petals (Ronse de Craene 2010, 2011) or opposite the sepals (Frame & Durou 2001). Although Tobe and Raven (1983a) suggested that Lecythidaceae have a multicellular archesporium, this would seem to be a mistake (see the observations of Tsou 1994). Are the ovules apotropous (Baillon 1877)?
The exact nature of the petal-like structures in the flower, especially in those of Napoleonoideae and some Asteranthoideae, has been a matter of much discussion. Ronse de Craene (2010, esp. 2011) considers the former subfamily, at least, to have five petals that become fused and plicate, and there are more or less thread-like staminodes developing from all three androecial whorls. Prance and Jongkind (2015) suggest that the innermost incurved whorl of staminodes and stamens of Napoleonaea may represent two whorls, one staminal, the other staminodial; this would imply that there are four androecial whorls. The single "perianth" whorl of Asteranthos could be equated with either of the petaloid whorls in Napoleonaea (one the corolla, the other staminodial); more developmental studies on Napoleonoideae and Asteranthoideae in particular are needed. (If Napoleonaea is considered to lack petals, the basic condition of the corolla for the family is ambiguous - c.f. Ronse de Craene 2011.)
The diversity of embryo morphology, and hence details of germination, in Lecythidaceae is considerable (e.g. Payens 1967 and references). Thus Prance and Mori (1979, see also Mori & Prance 1990a) discuss embryo morphology in Lecythidoideae in particular. Here embryos may be macropodial, i.e. the embryo is made up largely of hypocotyl plus radicle, any cotyledons at most being inconspicuous; germinating seeds with such embryos may penetrate the seed coat at opposite ends - or opposite sides. Arillate seeds are also common in this part of the family, and the aril may be flattened and form a wing, whether on one side (Cariniana) or all around (Couratari) the seed.
For general information, see van Tieghem (1905b, as Rhaptopetalaceae), Prance and Mori (1979: monograph, 2004), Mori and Prance (1990a: monograph), Prance (2008: Foetidia), Prance and Jongkind (2015: African taxa), Mori et al. (2017), and Mori's The Lecythidaceae Pages, Appel (1996, 2004), Letousey (1961), all as Scytopetalaceae, Liben (1971a) and Prance (2004), both Napoleonaceae, also Endress (1994b: floral morphology), Tsou (1994b) and Thammarong et al. (2019), both pollen of Barringtonioideae, Mori et al. (2015a: placentation, Lecythidoideae), Mauritzon (1939a), Venkateswarlu (1952a), Vijayaraghavan and Dhar (1976: Scytopetaloideae) and Tsou (1994a: not Scytopetaloideae), all embryology, Tsou and Mori (2002: seed coat anatomy in Lecythidoideae), and Takhtajan (1992: endothelium and testal vasculature.
Phylogeny. The set of relationships [Napoleonaeoideae [Asteranthoideae [Lecythidoideae [Planchonoideae + Foetidioideae]]]] were recovered by Morton et al. (1998), Mori et al. (2007) and Rose et al. (2018). Initially there were no rbcL sequences for Crateranthus, but it was placed with Napoleonaea in joint analyses (see also this site prior to x.2014); matK analyses place it with Asteranthoideae (D. Kenfack, pers. comm.). The relationships of Asteranthos were uncertain (e.g. Prance & Mori 1979; Mori & Prance 1990). In chemistry, morphology, etc., including its connate, serrate-margined "petals", Asteranthos is similar to Napoleonaeoideae (but c.f. style, endosperm), yet sequence data align it with Asteranthoideae, and it is florally quite similar to Crateranthus, also in Asteranthoideae. For phylogeny, see also Morton et al. (1997c, esp. 1998).
Within Lecythidoideae, there is a terminal polytomy made up of four genera (with a total of 115+ species) that is only weakly supported, and so it may collapse into a larger polytomy that also includes the [Allantoma + Cariniana decandra] clade (= Allantoma s.l., see Y.-Y. Huang et al. 2008; Mori et al. 2007). Thomson et al. (2018) obtained a well-resolved tree in an analysis of 24 chloroplast genomes (e.g. Eschweilera polyphyletic), but a tree resulting from the analysis of the same taxa, but with 354 nuclear loci added to Thomson's data, resulted in trees with rather different topologies (e.g. Eschweilera monophyletic in nuclear analysis) (Vargas et al. 2019). Although the basal Lecythidoideae were always [[Grias + Gustavia] [ Couroupita ...]], there was otherwise substantial disagreement between the chloroplast and nuclear trees (Vargas et al. 2019). A morphological analysis of some 86 Lecythidoideae provided little phylogenetic structure, the biggest of the clades with over 50% bootstrap support (52%) containing only six species (Huang et al. 2011). Huang et al. (2015) found that Lecythis and Eschweilera were in eight clades among which Corythophora and Bertholletia were interspersed. There also may be polyphyly in Couratari (Vargas & Dick 2020). Confirmation of relationships would be comforting... Indeed, Vargas et al. (2024: 343 nuclear loci, ca 140 taxa) produced a concatenated nuclear phylogeny for the Bertholletia clade in which many relationships had good support and which allowed them to disentangle the limits of genera; there was some conflict with a plastid tree (10 loci) they also recovered. Chave et al. (2024: 86 spp., plastome data, 571 regions nuclear DNA) also noted conflict between nuclear and plastid topologies, nuclear results suggesting that Lecythis should be divided into four genera, and they documented substantial and quite recent diversification in the Bertholletia clade (see above).
Classification. Asteranthoideae (the old Scytopetaloideae plus Asteranthos) are placed as a subfamily in an extended Lecythidaceae, which can more or less be characterised, however, Lecythidaceae, as restricted to the last three subfamilies in the summary phylogeny above, cannot. Appel (1996) morphologically characterized two major groupings in Asteranthoideae. For the beginnings of a phylogeny-based classification of Lecythidoideae, see Y.-Y. Huang et al. (2015) and Mori et al. (2015b).
Vargas et al. (2024) has recently reworked the genera in the Lecythis-Eschweilera area, recognizing 6 more genera and providing some 60 combinations. The result is that the generic names of a number of these Lecythidoideae - a number ecologically very important - have recently been changed, Lecythis, for example, now barely merits a mention (Vargas et al. 2024.)
Previous Relationships. Scytopetalaceae were considered quite distinct until recently, e.g. by Cronquist (1981), placed in Theales, and Takhtajan (1997), placed in Ochnales, both members of Dilleniidae.
Botanical Trivia. Grias is a pachcaul and G. purpuripetala may have obovate leaves ca 2 m long (Mori et al. 2015b). The age of some individuals of Cariniana micrantha from lowland Amazonia has been estimated to be some 1,400 years or more (Chanbers et al. 1998).
[[Sladeniaceae + Pentaphylacaceae], [Sapotaceae [Ebenaceae + Primulaceae]], [Mitrastemonaceae, Theaceae, [Symplocaceae [Styracaceae + Diapensiaceae]], [[Sarraceniaceae [Roridulaceae + Actinidiaceae] [Clethraceae [Cyrillaceae + Ericaceae]]]]] - if this group exists: endothelium?
Age. K. Bremer et al. (2004a: note topology) estimated the age of this node at around 107 My and Fu et al. (2019: note topology) an age of ca 87.5 Ma; ca 69.1 Ma is the estimate in Tank et al. (2015: Table S1), but stem Pentaphylacaceae, which are included here, are 83.4 Ma (Table S2), so something has gone wrong.
Chemistry, Morphology, etc.. Vessel elements with vestured pits or walls are scattered, if uncommon, in this group - e.g. in some Symplocaceae, Theaceae, Ericaceae, Clethraceae, and Pentaphylacaceae (Ohtani 1983; S. Jansen et al. 1998 for general summary).
[Sladeniaceae + Pentaphylacaceae]: evergreen, woody; vessel elements with scalariform perforation plates; vessel-fibre pits bordered; nodes 1:1; petiole bundle arcuate; mucilage cells +; hairs unicellular; inflorescences/flowers axillary; C ± campanulate, only basally connate, white/whitish; A basifixed; pollen 14-30 µm long, surface usu. little ornamented; nectary 0; placentae becoming ± swollen; ovules bitegmic, micropyle endostomal, inner integument 3-4 cells across; fruit a loculicidal capsule, columella separate, persistent, K persisting; endosperm +, embryo long.
Age. K. Bremer et al. (2004a) estimated this node to be ca 102 Ma, while Fu et al. (2019: ?topology) gave it an age of ca 78.7 Ma, Rose et al. (2018) estimated an age of ca 95.1 Ma, (97-)92(-90) Ma is the estimate in Wikström et al. (2015) and (100-)87.5(-73.5) Ma in X.-Q. Yu et al. (2017: note overall topology).
Pentapetalum trifasciculandricus is a fossil ca 91 Ma from New Jersey, U.S.A., that has been placed either with Theaceae or in the Pentaphylacaceae area depending on the analysis (Martínez-Millán et al. 2009); the latter position was preferred by Martínez-Millán (2010). The analyses in López-Martínez et al. (2023a) suggested other possibilities as well.
Chemistry, Morphology, etc.. Sepals somewhat narrowed towards the base seem to be common here (see Jia et al. 2019: Fig. 7). The placenta is very well developed in Ficalhoa and many Ternstroemieae and Frezierieae.
Beauvisage (1920) remains a useful account of the vegetative anatomy - and general morphology - of the old Ternstroemiaceae. See Luna and Ochoterena (2004) and Martínez-Millán et al. (2009) for morphology.
Phylogeny. Luna and Ochoterena (2004) and Martínez-Millán et al. (2009) were unable to recover much in the way of strongly supported relationships in this area in morphological phylogenetic analyses. In some analyses in the latter paper Calophyllaceae (see Malpighiales) were included in Theales, and adding morphology in joint analyses tended to reduce support measures, perhaps especially bootstrap support.
A position of Pentaphylacaceae in Ericales seems reasonable from the gross morphological point of view. The anthers are superficially like those of Diapensiaceae, while Pentaphylax and Theaceae s.l. are generally similar. The seed is Ericalean (Huber 1991). For further discussion on the relationships of this clade, see the introduction to Ericales above.
Classification. Pentaphylacaceae had been recognised as a monotypic family for some time (see e.g. A.P.G. 1998; first versions of this site), and A.P.G. II (2003) suggested as an option recognising three families, i.e. Pentaphylacaceae, Ternstroemiaceae, and Sladeniaceae. However, the first two are quite similar phenetically, far more so than they are to Sladeniaceae, and so two families are recognised in A.P.G. III (2009), Pentaphylacaceae being expanded to include Ternstroemiaceae.
Previous Relationships. See Theaceae for a family that largely included this whole group in the past.
SLADENIACEAE Airy Shaw - Back to Ericales
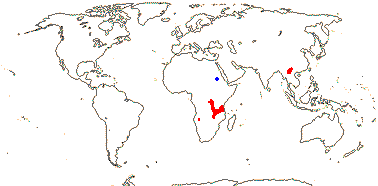
Chemistry?; cork cambium pericyclic; intervessel pitting opposite-alternate; lamina margins toothed; inflorescences cymose; pedicel strongly articulated with flower; flowers small [5³ mm long]; A (8-)10(-13), 15, anthers opening apically; style at most short, with relatively long pointed lobes; x = 12 (?6, ?24).
2/3: [list]. S.E. Asia, tropical E. Africa.
Age. Rose et al. (2018) thought that the two genera included here diverged ca 88 Ma.
1. Ficalhoa laurifolia Hiern
Exudate +; vessels not grouped; perulae 0; leaves 2-ranked, lamina vernation involute; C connate; A 15, in groups of 3 alternating with C, anther thecae dehiscing as slits across the apex; G [5], opposite K, style short, slightly impressed; placentation axile, placentae large, ovules many/carpel; fruit loculicidal capsule, smooth, K deciduous, style persistent; testa crustose, exotesta cells ± polygonal, little thickened; endosperm slight; n = ?
1/1. Tropical E. Africa. Map: see Verdcourt (1962), Trop. Afr. Fl. Pl. Ecol. Distr. Vol. 1 (2003) and C.-C. Yu et al. (2025).
2. Sladenia Kurz
Vessels in radial groups; petiole also with wing bundles; perulae +; leaves spiral, lamina (margins entire); C basally connate; A (8-)10(-13), anthers sagittate, dehiscing by pores, monocot anther wall development; microsporogenesis successive [tetrads tetragonal]; G [3], ridged, gradually narrowed to indistinct/short style; placentation apical, ovules 2/carpel, outer integument ca 3 cells across; embryo sac tetrasporic, 8-nucleate [Adoxa type]; fruit a ?schizocarp, longitudinally ridged, K persistent, endocarp crustaceous; seeds irregulary winged, testa thin, transparent; endosperm 0; n = 24.
1/2. S.W. China, adjacent Myanmar, Thailand and Vietnam. Map: see above, from Flora of China Vol. 12, also Jia et al. (2019: Fig. 1), for African fossil Sladenia [blue], see Giraud et al. (1992) and C.-C. Yu et al. (2025: Fig. 1, inc. Southeast Asian distribution and fossils).
Age. The wood of extant Sladenia is distinctive, and fossil wood Sladenioxylon african initially thought to be ca 100 Ma from the ?Cretaceous-Albian/Cenomanian deposits of northern Sudan match it quite closely (Giraud et al. 1992), although redating has suggested that this wood is only around 72 Ma (references in Atkinson et al. 2017). Fossils of Sladenia are reported from the Iberian peninsula/Balearic Islands, becoming extinct there in the Oligocene (Postigo-Mijarra et al. 2009). Inflorescences from Yunnan have been dated to 23-16 Ma (Jia et al. 2019).
Evolution: Divergence & Distribution. Sladenia is another case where past and present distributions are very different. Jia et al. (2019) thought that Sladeniaceae originated in Africa, and using climatic reconstructions C.-C. Yu et al. (2025) suggested that that Ficalhoa and Sladenia overlapped in Africa ca 66 Ma and again 14.9-4 Ma - i.a. this might help explain the similarities in the plastomes of the two.
Genes & Genomes.
Chemistry, Morphology, etc.. The pollen morphology and wood anatomy of Sladenia are very similar to those of Pentaphylacaceae, but there are no sclereids. Ficalhoa is very poorly known; it, too, lacks sclereids, but it was not associated with Sladenia in anatomical studies (especially Deng & Baas 1991). Li et al. (2003) have recently described a number of very distinctive embryological, etc., features for Sladenia, including monocot anther wall development; it will be interesting to see if Ficalhoa is similar in these respects.
For general information, see Stevens and Weitzman (2004).
Phylogeny. Sladenia was sister to Pentaphylacaceae (Ternstroemiaceae) in rbcL studies (Savolainen et al. 2000b), albeit the DNA was rather degraded. Sladenia and Ficalhoa come out as sister taxa in some recent molecular analyses (Anderberg et al. 2002); note, however, that Schönenberger et al. (2005) did not find support for this clade.
Previous Relationships. Sladenia has often been included in Theaceae, e.g. as Sladenioideae (see Takhtajan 1997).
PENTAPHYLACACEAE Engler, nom. cons. - Back to Ericales
Plants Al-accumulators; parenchyma apotracheal, diffuse or in short tangential lines; intervessel pitting opposite-scalariform; stomata mostly paracytic; lamina supervolute; flowers single, from axils of reduced leaves; first A whorl opposite K, anthers with crystals in the connective [?Pentaphlylax]; style hollow; ovules campylotropous to hemitropous, apotropous when few [?Symplococarpon]; testa multiplicative, mesotesta well developed; embryo U-shaped/curved; x = 12/23 (?22).
12/345: [list - as tribes] - three tribes below. Tropical and subtropical, but few in Africa.
Age. Rose et al. (2018) estimated the crown age of this clade to be ca 78.1 Ma.
Pentaphylax and Visnea are reported fossil from late Cretaceous (Maastrichtian, ca 69 Ma) and Eurya from Santonian (ca 85 Ma) deposits in Europe (Knobloch & Mai 1986). If the latter date is confirmed, it would suggest an age for this node of around 90 Ma or so.
1. Pentaphylaceae P. F. Stevens & A. L. Weitzman - Pentaphylax euryoides Gardner & Champion
Chemistry?; druses 0; buds perulate; lamina margins entire; A 5, thecae valvate, filaments very broad, narrowed and incurved apically; pollen smooth, tectum thin, columellae poorly developed, endexine thick; G [5], opposite ?sepals, stigmas shortly radiate; ovules 2/carpel, apical, apotropous, outer integument ca 2 cells across, inner integument ca 2 cells across, ?parietal tissue; fruit with midrib separating from rest of valve [= teeth], endocarp cells transversely elongated; seeds flattened; exotestal cells slightly thickened, elongated, mesotestal cells large, ± thin-walled; endosperm development?, slight, cotyledons longer than radicle; n = ?
1/1. China (Kwangtung and Hainan) to Sumatra, scattered.
[Ternstroemieae + Frezierieae]: ellagic acid +, iridoids 0; (pits vestured); pith often with diaphragms; sclereids +; perulae 0; C (whitish to yellowish to greenish); androecium initially with ± indistinct ring primordium, stamen formation centripetal; filaments to 2x longer than anthers, latter variable in length, connective usu. (much) prolonged; exothecium persistent, thickened or not; endothecium (not thickened), pseudopollen +; integument 3-4 layers across; fruit ± fleshy; mesotestal cells lignified, ± crystalliferous; endosperm +, ?nuclear, cotyledons incumbent, shorter than radicle.
Age. This node was dated to around 66.7 (Rose et al. 2018), (61-)55, 54(-48) (Wikström et al. 2001), (69-)54, 51(-35) (Bell et al. 2010) or ca 48.3 Ma (Fu et al. 2019).
2. Ternstroemieae de Candolle —— Synonymy: Ternstroemiaceae de Candolle
Sclereids much branched; leaves ± pseudoverticillate, lamina often with black spots, margins entire to crenulate and/or with black, deciduous setae; K opposite C, petaloid or not, C to 2/3 connate, (with narrowed then somewhat broadened erect apical portion - Anneslea); A fasciculate, fascicles opposite C, development centripetal, filaments shorter than anthers; endothecium thick; G [2-3], (inferior - Anneslea), style + (deeply divided)/± 0, stigmas punctate/peltate; ovules 1-12/carpel, apical-axile, outer integument 6-9 cells across; fruit becoming inferior [Anneslea], dehiscing irregularly/loculicidal/basally circumscissile/indehiscent; seeds few, ± dangling, ≥3 mm long, brown, sarcotestal [either exotesta or pockets of fleshy cells on either side of seed], unlignified exo-/mesotesta to 10 cells across, lignified mesotesta 7-15 cells across; n = 20, 25.
2/103: Ternstroemia (100). Tropics, esp. Malesia and Central to South America. Map: from Camp (1947), Trop. Afr. Fl. Pl. Ecol. Distr. vol. 1 (2003), Australia's Virtual Herbarium (consulted viii.2013) and M. Sosef (pers. comm.). [Photo - Flowers & Fruits © Nick Turland.]
3. Freziereae de Candolle
(Nodes 1:3, 3:3 [some Freziera]); sclereids usu. little branched; leaves scattered along shoot, two-ranked (spiral), lamina margins entire to serrate; plant dioecious or flowers perfect; inflorescence also fasciculate, (flowers from axils of expanded leaves); (K connate); C (urceolate), (orange-red - Balthasaria, purplish); A 5-30(-60), from ring primordium, in a single whorl, A opposite K initiated first [Cleyera], (filaments to 5x longer than anthers - Cleyera), (connective not prolonged); (theca epidermis lignified); G [(1-)3(-10)] (inferior - Symplococarpon), (placentation parietal/also free central - Adinandra), (styluli +); ovules 4-many/carpel, outer integument 3-4 cells across; fruit indehiscent, fleshy, a berry (drupe); seeds (1-)many, <4(-6) mm long, brown or black, inner walls of exotesta thickened and lignified or not, lignified mesotesta 1-5 cells across; (embryo curved); n = (12, 13(?), 15, 18) 21 (-23), etc.
9/255: Adinandra (80), Eurya (75), Freziera (75). Southeast Asia to Malesia, Hawai'i, Central to South America, E. (Balthasaria) and W. (Adinandra) Africa, and the Canaries (Visnea). Map: from Camp (1947), Verdcourt (1962), van Balgooy (1975) and Weitzman (1987). [Photo - Eurya Flower, Flowers & Fruits, Flower, Fruit.]
Age. The crown group age of Freziereae is ca 51.9 Ma (Rose et al. 2018: Visnea sister).
Fossils of Eurya are reported from Europe from the late Cretaceous ca 70 Ma onwards (H. Zhu et al. 2016) - which disagrees with the ages above, and of course Eurya is now no longer known from Europe.
Evolution: Divergence & Distribution. Visnea mocanera (Frezierieae), from the Canary Islands/Madeira, is very isolated (see map) from the rest of the family, whether as the result of vicariance as the North Atlantic opened 120-100 Ma (Grehan 2017: less likely), or dispersal from the East, Palaeo-Macaronesia being 60 Ma or more old (the oldest currently emergent Canary Island is ca 21 My: see Gelmacher et al. 2005; Fernández-Palacios et al. 2011).
Frost and Lagmarsino (2021) looked at diversification in the cloud forest Freziera, a genus with rather undistinguished flowers but a considerable amount of vegetative variation. This seems to have begun in the northern Andes ca 7 Ma (the phylogenetic fuse was ca 5.6 Ma), with species there moving into a variety of habitats. In a subsequent Angiosperms353 analysis Frost et al. (2024: 50 spp) recovered nine more or less well suppported clades, but relationships between/within them varied somewhat and there seems to have been introgression, etc..
Pollination Biology. Buzz pollination has been described in Ternstroemia. Here the partly connate petals, as in T. laevigata, form a tube with a narrow, fluted apical opening, i.e. the end result is rather like the anther cone of Solanum and other buzz-pollinated flowers, while the spreading sepals with their red margins appear to function as petals. The spreading sepals of T. rupestris are red, those of T. longipetiolata are greenish with white-pink margins, and so on, the petals closely surroung the stamens are whitish, or, in the case of T. longipetiolata they are purple (Bittrich et al. 1993; see also Amorim et al. 2019; Vieira et al. 2021, J. A. Vieira pers. comm.). However, T. gymnanthera and some other Old World species have spreading petals and exposed stamens. Anneslea has distinctive petals that become narrowed, but a somewhat broadened erect apical portion surrounds the protruding style - buzz pollination here, too? On the other hand, Cleyera, and also Eurya (for which see W. H. Brown 1938), secrete nectary from the basal part of the ovary wall, and in E. japonica in particular the flowers produce oligosulphides - the scent is described as "rather putrid" (Trees and Shrubs Online) - and are probably carrion or dung mimics (Okuyama et al. 2025).
Genes & Genomes. Hao et al. (2010) note that the atp1 mitochondrial gene in species of Ternstroemia is highly chimaeric, and transfer (?how), probably from Vaccinium, may have occurred ca 15-50 Ma; some "host" genes have been converted by Vaccinium mitochondrial genes...
C.-C. Yu et al. (2025) note that there appears to have been some transfer of genes fro Visnea to Ficalhoa and much more substantial transfer from Eurya to Freziera. For similarities in the plastomes of Ficalhoa and Sladenia, see above.
Chemistry, Morphology, etc.. Freziera shows considerable variation in nodal anatomy and stomatal morphology (Weitzman 1987). Although the leaves of Pentaphylax are entire, they, the bracts, and some sepals, are terminated by blackish, deciduous and probably glandular points, rather similar to those found in the rest of the Pentaphylacaceae. Its pericylic sheath consists of fibres alternating with lignified parenchymatous cells (Beauvisage 1920). Cleyera (Freziereae) lacks pericyclic fibres in the petiole.
The flowers are very often single in the axils of reduced leaves; if the shoot of Pentaphylax does not develop expanded leaves after the flowers appear, the inflorescence appears to be racemose. If the flower-bearing shoot is very much reduced, then the inflorescence is fasciculate. For variation in seed type and pollen surface of Freziera, see Weitzman (1987). As R. Zhang et al. (2020) describe the gynoecium of Adinandra latifolia, there is axile placentation apically, but at the base separate placental mounds develop and fuse, the result being free-central placentation ("deuterogenic placentation") that is separately vascularized from the apical placentae. The reports of an aril in Ternstroemieae (e.g. Keng 1962) are incorrect; there is a sarcotesta which may, by its expansion, aid in the irregular rupture of the fruit.
Many references are to Ternstroemiaceae: For general information, see Weitzman et al. (2004) and Y. H. Wang et al. (2006: phylogeny, focus on Theaceae), for floral development, see Tsou (1995), R.-J. Zhang et al. (2007), Zhang and Schönenberger (2014) and Ronse de Craene (2018), for pollen, see Lobreau-Callen (1977) and Wei (1997), and for some embryology, see Mauritzon (1936a).
Pentaphylax is particularly poorly known.
Phylogeny. For a phylogeny that includes Theaceae s. str. and a few other Ericales, see J.-B. Yang et al. (2006: mitochondrial matR analysis, relationships unclear) and Su et al. (2011: mostly Theaceae included); the latter found that Euryodendron was well supported as sister to Eurya (see also Ohi-Toma et al. 2025. The tribes above were recovered by Tsou et al. (2016), although relationships within Freziereae in part depended on the marker used, and also by Rose et al. (2018). Frost and Lagomarsino (2021) and Frost et al. (2024) looked at relationships in Freziera using the Angiosperms353 probe on herbarium specimens (50/75 spp.). After a careful analysis of the data they obtained a fair bit of resolution, nine main clades being evident (esp. Frost et al. 2024).
C.-C. Yu et al. (2025) looked at relationships between the old Sladeniaceae, Ficalhoa and Sladenia, and Pentaphylaceae (6 or 7 genera of the latter, mostly represented by one species) and found these differed depending on whether whole plastome or Angiosperms353 nuclear data were used. Analysis of the former suggested that Sladeniaceae were sister to Pentaphylacaceae, analysis of the latter suggested the relationships [ Sladenia [Ficalhoa + Pentaphylaceae]]; relationships between the Pentaphylacaceae included were congruent both with each other and with relationships previously suggested within Pentaphylacaceae.
Classification. Pentaphylacaceae-Frezierieae and -Ternstroemieae are morphologically amply distinct from Theaceae. The former have pollen 14-28.5 µm long (versus 36.5-54.5 µm), vessel-fiber pits bordered (versus unbordered), etc.. However, differences in the relative length of the radicle in the embryo (long radicle in Pentaphylacaceae, short in Theaceae) are not so clear-cut given the inclusion of Pentaphylax itself and Sladeniaceae in the mix. Inclusion of Sladeniaceae in Pentaphylaceae (see C.-C. Yu et al. 2025 and above) necessitates tribal status for both Ficalhoa and Sladenia.
Previous Relationships. Theaceae often included Ternstroemia and relatives; thus Ternstroemioideae were a subfamily of Theaceae in Takhtajan (1997). On the other hand, Beauvisage's (1920) Ternstroemiaceae included Theaceae, even if he also removed some seventeen separate elements from the family (including Pentaphylax), most of which he thought were unrelated to each other. They included genera now placed in Marcgraviaceae, Ochnaceae-Medusagynoideae, Calophyllaceae, Bonnetiaceae, Actindiaceae, Stachyuraceae, Strasburgeriaceae, and so on. Cronquist (1981) circumscribed Theaceae quite broadly, but the families he did remove he thought were close to them. Theaceae were a major linking family in evolutionary classifications - "It is generally agreed that the Theaceae are closely related to the Dilleniaceae..." (Cronquist 1981: p. 323).
Age. The age of a [[Sapotaceae + Ebenaceae] Primulaceae] node - if it exists - is around 84.3 or 82.3 Ma (Tank et al. 2015: Table S1 and S2), 94.4 Ma (Magallón et al. 2015), (101-)92(-79) Ma (Wikström et al. 2015), ca 102.3 Ma (Rose et al. 2018) or 80.2-73.5 Ma (H.-F. Yan et al. 2024). The age of a [Sapotaceae + Ebenaceae] node - if it exists - is estimated to be (116.3-)111.9(-106.9) Ma (Bansal et al. 2021), while the age of an again perhaps notional [Sapotaceae + Primulaceae s.l.] node - [Ebenaceae + Lecythidaceae] are sister to them - is ca 102 Ma (Bremer et al. 2004a).
Age.
SAPOTACEAE Jussieu, nom. cons. - Back to Ericales
Trees and shrubs; saponins, C-30 oxidised triterpenes, pyrrolizidine alkaloids, flavonols, leucodelphinidin, gutta, myricetin +, ellagic acid 0; nodes (1:1) 3:3; (medullary bundles +); petiole bundle arcuate, horizontal D-shaped or annular (wing bundles +); latex sacs +; sclereids +; hairs T-shaped, arms unequal or not, unicellular (not in Delpyodon), brownish; leaves (two-ranked, opposite), lamina vernation conduplicate, margins entire (toothed), secondary veins often rather close, (stipules +, cauline); inflorescences cymose, pedicels not articulated; flowers 5-merous; K ± connate at base, C connate; A = C, opposite C, introrse to extrorse, staminodes +, opposite K; tapetal cells multinucleate; pollen 3-6-colporate, infratectum ± granular; disc + (0); G with hairs on the inside of the ovary, placentation axile to axile-basal, (style short), stigma punctate or minutely lobed, dry; ovules 1/carpel, ascending, apotropous, integument single, "thick", (vascularized), hypostase 0; fruit a berry (drupe), K persistent; seeds large, hard, shiny, hilar scar large, white; testa multiplicative, outer part with isodiametric heavily lignified cells; endosperm nuclear/coenocytic; n = (10-)13(-14), x = 12 (?13), genome size [1C] (0.049-)0.608(-7.503) pg.
73/1,242: [list] - Three subfamilies below. Pantropical.
Age. Crown-group Sapotaceae have been dated to (105.0-)84.5(-67.1) Ma (Richardson et al. 2013: this age is given as ca 107 Ma by Phang et al. 2023), (126-)107(-88) Ma is the age in Armstrong et al. (2014), while Rose et al (2018) estimate an age of only ca 58.3 Ma.
1. Sarcospermatoideae Swenson & Anderberg
?
2/14. S. China, Indo-Malesia. Map: from Aubréville (1964).
Age. Hofmann (2018) recorded a diversity of sapotaceous pollen from western Europe; by around 56 Ma there was pollen she placed in Sarcospermatoideae; see also Chrysophylloideae and Sapotoideae.
1A. Sarcosperma —— Synonymy: Sarcospermataceae H. J. Lam
Calcium oxalate crystals, crystal sand +; fibre pits distinctly bordered; leaves opposite or not, (stipels +); inflorescence axis well developed; A basifixed, staminodes short, broad, scale-like; disc 0; G 1[-2], style stout; seeds not laterally compressed; endosperm 0, cotyledons ± connate; n = ?
1/11. S. China, Indo-Malesia. Map: from Aubréville (1964).
[Eberhardtia [Sapotoideae + Chrysophylloideae]] (if this clade exists): inflorescence fasciculate.
Age. Rose et al. (2018) suggest that this node is ca 41 Ma old.
1B. Eberhardtia Lecomte
Mineral inclusions 0; C members with three segments, centre segment narrow; A 5, staminodes with versatile inverted V-shaped apex; endosperm copious; n = ?
1/3. South China, Vietnam, Laos, Sabah.
Note: Sideroxylon may also be around here...
[Sapotoideae + Chrysophylloideae]: silica bodies +[?plesiomorphic]/0, prismatic crystals +/0; (vessel elements with scalariform perforation plates); (stipules well developed); (plants di- or monoecious); K (in two whorls), C variously lobed or not; A (in groups up to 6 opposite C), staminodes often ± C-like (0); G 1-[2-14(-30)]; ovules (2-5/carpel); endosperm +/0, (cell walls with xyloglucans [thick, pitted, amyloid]); nuclear genome [1 C] (274-)1979(-2513) Mb. [Photos - Collection, Fruit].
Throughout the tropics. Map: from Aubréville (1964).
Age. This node, containing the bulk of diversity in the family, is estimated to be a mere 30.4 Ma by Rose et al. (2018: note that species of Sideroxylon are sister to both subfamilies) but 75 Ma by Bansal et al. (2021).
2. Chrysophylloideae LuerssonA high in the tube, (staminodes outside/above the staminal whorl); G (style with separate stigmatic areas); endosperm 0 (+), cotyledons plano-convex/foliaceous, radicle (exserted below commissure)/not.
44/672: Planchonella (103), Pycnandra (66), Pleioluma (ca 40), Micropholis (41),Chrysophyllum (27). Pantropical.
Age. Crown Chrysophylloideae are some (105-)91.7(-79) Ma (Bartisch et al. 2010).
There was pollen from at least two clades in Chrysophylloideae, one now largely South American and another Australasian, by around 56 Ma in western Europe - see Hofmann (2018).
3. Sapotoideae Eaton
Stipules cauline/0; (C lobes each with three segments); seed with lateral? hilum.
27/546. Pantropical.
Age. An estimate of the age of crown-group Sapotoideae is ca 72.5 Ma (from Fig. 1 in Armstrong et al. 2014), ca 72.4 Ma (Boluda et al. 2022) or (59.3-)52.5(-45.7) Ma (Swenson et al. 2022). Note that Sideroxyleae are sister to Chrysophylloideae - (56.6-)53.6(-52.2) Ma is the age of the combined clade - in Richardson et al. (2013).
Late Cretaceous pollen, Sapotaceoidaepollenites rotundus, ca 71 Ma and from off South Australia, may be close to that of Sapoteae (see Richardson et al. 2013). Hofmann (2018) recorded a diversity of sapotaceous pollen from western Europe; by around 56 Ma or soon after there was pollen identifiable as Sapotoideae.
3A. Sideroxyleae Small —— Synonymy: Bumeliaceae Barnhart
(thorns +).
2-3/85: Sideroxylon (83), (Sub)tropical, Pacific Islands.
Age. Crown-group Sideroxyleae are estimated to be (73-)62(-52) Ma (Armstrong et al. 2014) or (50.3-)44.5(-38.3) Ma (Richardson et al. 2013).
[Gluemineae [Tseboneae [Inhambanella [Sapoteae + Isonandreae]]]]: ?
Age. This clade is ca 59.1 Ma (Boluda et al. 2022).
3B. Gluemineae - tribal name needed
fruit explosively dehiscent.
3/11: Lecomtedoxa (6). Tropical West and West Central Africa.
Age. Gluemineae are ca 50.5 Ma (Boluda et al. 2022).
[Tseboneae [Inhambanella [Sapoteae + Isonandreae]]]: ?
Age. This clade - but note topology - is ca 56.0 Ma (Boluda et al. 2022).
3C. Tseboneae L. Gautier & Naciri
?
3/34: Capurodendron (32 - prob. ca 46). Madagascar.
Age. Crown Tseboneae are (51.3-)45.7(-39.5) Ma (Boluda et al. 2022).
[Inhambanella [Sapoteae + Isonandreae]]: ?
Age.The age of Inh. Sap. is ca 51 Ma (Boluda et al. 2022).
3D. Inhambanella Dubard
?
1/2. Tropical Africa
Age. The two species diverged ca 23.8 Ma (Boluda et al. 2022).
[Sapoteae + Isonandreae]: (K in two whorls, 2-4 valvate members in each).
Age. The age of this node is estimated to be (48.3-)40.5(-32.6) Ma (Richardson et al. 2013) or ca 55 Ma (Phang et al. 2025).
3E. Sapoteae Reichenbach (Mimusopeae) —— Synonymy: Achradaceae Vest, Boerlagellaceae H. J. Lam
K in two whorls, C with lateral appendages (not); A = C.
Manilkara (ca 80), Mimusops (ca 40)
Age. Sapoteae are estimated to be (44-)43(-42) Ma (Armstrong et al. 2014), ca 38.8 Ma (Richardson et al. 2013) or ca 43.6 Ma (Boluda et al. 2022).
Pollen like that of Vitellariopsis marginata (Sapoteae), now ± tropical southern African, is reported from Austria in deposits only some 17-11 Ma (Hofmann & Lichtenwagner 2019).
3F. Isonandreae Hartog
?
6/265: Palaquium (110: polyphyletic), Madhuca (114: polyphyletic). Indo-Malesia, also S. China, Hainan, Australia and the West Pacific.
Age. Crown-group Isonandreae are (43.6-)36.5(-29.0) Ma (Richardson et al. 2013), ca 43.9 Ma (Boluda et al. 2014) or ca 47.7 Ma (Phang et al. 2025).
There is pollen similar to that of Palaquium and Madhuca in deposits from Europe ca 56 Ma (Hofmann 2018; see Phang et al. 2025).
Evolution: Divergence & Distribution. For ages in the Manilkara (Sapoteae) area, see Armstrong et al. (2014) and for those in Capurodendron (Tseboneae), see Bioluda et al. (2014).
Bartish et al. (2010) discuss the historical biogeography of Chrysophylloideae, and thought that long distance dispersal dominated when explaining the current distributions of members of the group. The southeast Asian Xantolis is sister to the rest, and early diversification of other Chrysophylloideae perhaps occurred in Africa in the Campanian 83-73 Ma, although much diversification in the subfamily is Caenozoic in age. It is possible that Australian elements arrived from America via an Antarctic land bridge (Bartish et al. 2010, q.v. for further discussion, dates, etc.). The New Caledonian Pycnandra which, with around 59 species, is the largest endemic clade on the island (and one of the oldest such clades), may have reached there in the latter part of the Oligocene ca 29.8 Ma (it is sister to Niemeyera with 3 species and from eastern Australia); its crown-group age is ca 16.2 Ma (Swenson et al. 2008c, see also Isnard et al. 2020; c.f. Ladiges & Cantrill 2007). For diversification within Pycnandra, which seems to include cryptic species separated by geography, soil and/or altitude, see Swenson et al. (2015). Overall, it is estimated that Chrysophylloideae have moved to New Caledonia around nine times since the emergence of the island around 37 Ma (Grandcolas et al. 2008; Swenson et al. 2014; see also Nattier et al. 2017). Swenson et al. (2022) discussed the history of Gayella, from Chile but embedded in an otherwise Antipodean clade; they thought that this might have been the result of vicariance, although long distance dispersal was also possible. Planchonella, with a crown-group age of (54.2-)44.6(-36.6) Ma, may have originally been from the Sahul Shelf area but with subsequent very extensive movement throughout the Pacific and to the Indian Ocean, with the very widely-distributed P. obovata (species limits?) being disjunct between Palau and the Seychelles, 8,900 km apart (!: Swenson et al. 2019). Judging by the ages of the species on various islands/archipelagoes, Tertiary metapopulation vicariance seems not to have been involved (Swenson et al. 2019).
Looking now at Sapotoideae, the descendents of a possible ancient hybridisation 43-36.6 Ma event between a basically African clade and a basically American clade in Sideroxylon (Sideroxyleae) were previously segregated as Nesoluma. They are now to be found on very young islands in the Pacific and may have been hopping from island to island for the last 40 Ma or so (Smedmark & Anderberg 2007), rather like Planchonella; for other taxa behaving similarly, see Hillebrandia [Begoniaceae], Psiloxylum [Myrtaceae], various Rutaceae, etc.. Isonandreae perhaps originated In Africa, crown Isonandreae diversifying in Sundaland where they show a W→E diversification pattern across Wallace's Line (there has also been movement to the Indian subcontinent, and so on), and there is also much dispersal here both across water and over land, especially from the Sundaland area (Richardson et al. 2013; Phang et al. 2025 - c.f. dates). Sapotoideae in general are quite old, but it is unlikely that drift can be invoked to explain their disjunct distributions, migration via the boreotropical route stopped around 33 Ma, and so long distance dispersal may well be responsible for disjunct distributions like those in Manilkara. This genus may have originated in Africa ca 29 Ma (Manilkarinae, also African, are ca 32 Ma), but it is now to be found throughout the tropics - and this may have involved perhaps five separate movements to Madagascar alone, from there moving to the East Malesian area (33-)28, 15(-10) Ma (Armstrong et al. 2014).
Morphological characters in general are highly homoplasious here and characters for the subfamilies are hard to come by; see Smedmark et al. (2006) for character evolution in Sapotoideae, while Swenson et al. (2023: Fig. 5) mapped 25 morphological characters on to a tree of Neotropical Chrysophylloideae. Beckers et al. (2025) discussed the variation and evolution of characters of wood anatomy in the family - overall, they thought there was some similarity with the wood of Lecythidaceae. However, they were hampered by the absence of a clean phylogeny for the family, not to mention the fact that there were 11 genera in their nuclear tree that were not in the plastome tree, and 24 genera in their plastome tree that were not in the nuclear tree. However, variation here is quite extensive and will repay further study - Beckers et al. (2025) noted that in a number of taxa there had been evolution of scalariform from simple perforation plates, and this was perhaps particularly notable in Palaquium. Although Sapotaceae are a homogeneous family from some points of view, Chartier et al. (2021) noted that their distinctive combinations of floral characters put them in the periphery of the floral morphospace that represented that of Ericales as a whole. Floral variation is considerable and most characters are very homoplasious (e.g. Swenson et al. 2008a, b, c). The flowers are sometimes described as being up to 6-merous, i.e. following the number of sepals in a single whorl, however, petals, androecium and gynoecium must then be considered to have doubled in number (see Pennington 2004: good summary of floral variation). Swenson and Anderberg (2005) suggest that the basic floral morphology of the family is K5, C5, A 5 + 5 staminodes, however, anisomery is scattered in Sapotaceae, with different numbers of parts in the different whorls (Swenson et al. 2008c; see also Wanntorp et al. 2011). Kümpers et al. (2016) looked at floral variation, particularly merosity, in Sapotaceae in considerable detail, and noted a variety of ways in which it could change, which led to increases in numbers of all or most parts of the flower, however, carpel and stamen number sometimes increased independently of any general changes in merosity. Swenson and Anderberg (2005) suggested that the staminodes common in Chrysophylloideae were perhaps derived within the clade, and so were mot immediately comparable with those of other members of the family; staminodes in the former are outside the staminal whorl while in the latter they are in the same whorl as the stamens. Genera like Omphalocarpum (near Chrysophylloideae) are remarkable in having up to 11-merous flowers with up to 6 stamens opposite each petal, the ovary has up to 30 loculi and develops into a very large drupaceous fruit the pericarp of which has a hard, woody layer well over a centimeter thick (O. procerum).
In an analysis using nuclear genes that included a number of accessions of Capurodendron (Tseboneae), a Madagascan endemic, Christe et al. (2021) suggested that there were over 20 undescribed species in that genus alone. Capurodendron, with 43(described)-55 species, is probably the second most diverse endemic Madagascan plant genus (Boluda et al. 2022, 2024: the palm Dypsis may be most diverse), but since the other two genera in Tseboneae are also from Madagascar the whole clade with some 58 species is endemic (Canarium (Burseraceae) has over 30 species in Madagascar). Note that in general Madagascan Sapotaceae are both diverse and poorly known - as of iii.2026 Kiedaisch et al. (2026) estimated that there were about 11 genera and 113 species there, plus ca 60 undescribed species. As genera have been revised, their species numbers may double (Boluda et al. 2022 - Capurodendron), or if species numbers have been little changed, as in Donella, then the limits of most species have had to be adjusted, and there one species, D. guereliana, may need to be placed in a separate genus (Kiedaisch et al. 2026).
Ecology & Physiology. Sapotaceae are notably common in terms of both numbers of species and individuals in the Amazonian tree flora, ranking second in the latter category (Fabaceae are #1) althouth they are not notably well represented in the 227 species there that make up half the stems 10 cm or more d.b.h. (ter Steege et al. 2008, 2013, but c.f. in part Levis et al. 2017; Maezumi et al. 2018). 22 species of Pouteria have been found in a single hectare there, and Sedio et al. (2017) suggest that such aggregations are allowed by extensive interspecific variation in secondary metabolites affecting herbivory. In eight Central African rainforests sampled, Lecomtedoxa is described as a hyperdominant species (Bastin et al. 2015), one of the 18 species that made up 50% of the above-ground biomass (ca 2%, #6 on the list).
The New Caledonian Sebertia (= Pycnandra) acuminata (sève bleue) is a nickel (Ni) hyperaccumulator, and its latex contains some 11% wet weight, 25.7% dry weight of the metal (Jaffré et al. 1976), the highest concentration of Ni known in any living organism. At least two other species of Pycnandra can also hyperaccumulate Ni, an ability that has evolved twice in the genus (but not at all in its Australian sister taxon, Niemeyera), and the latex may be involved in plant growth at the seedling stage (Isnard et al. 2020).
Pollination Biology & Seed Dispersal. Entomophily is common in the family, but bat pollination is known from some Sapotoideae-Isonandreae (Phang et al. 2025 and references).
Fruits of Sapotaceae tend to be eaten by unspecialized frugivores (Snow 1981). For the cautionary tale of the dodo and the tambalacoque (Sideroxylon grandifolium) on the Seychelles, see Herhey (2004); the dodo supposedly ate the fruits and simultaneously enabled the germination of the seeds, so when the dodo became extinct, the pant could no longer regenerate. However, no evidence was given that being eaten by the dodo was essential for seed germination, rather, it seems that introduced monkeys eat the young fruits in large numbers... Seeds of the argan tree, Argania spinosa, are dispersed when the goats that eat the fruit as they graze the canopy of the tree (the goats are arboreal) spit the seeds out while chewing their cud (Delibes et al. 2017). Bat dispersal of seeds is also known from some taxa (Phang et al. 2025).
Plant-Animal Interactions. Larvae of New Caledonian Apionidae weevils eat the embryos of Pycnandra, and this includes P. acuminata (its embryo is bright pink) despite the high concentrations of Ni in its latex; all told three genera of weevils and three genera of Sapotaceae - the other two genera are Planchonella and Beccariella (Wanat & Munzinger 2012; Isnard et al. 2020).
Genes & Genomes. The mitochondrial coxII.i3 intron is absent in Chrysophyllum, at least (Joly et al. 2001).
Economic Uses. Chicle, a complex rubber once used in chewing gum, is the exudate of Manilkara zapota.
Chemistry, Morphology, etc.. There is banded apotracheal parenchyma in Sapotaceae (c.f. Lecythidaceae). The 38 papers by Kukachka (1978-1982, up to 11 papers/year) deal with the wood anatomy of neotropical genera of Sapotaceae. Some species of Sarcosperma have paired stipels at the apex of the petiole, a rather unexpected character for a member of Ericales. Anderberg and Ståhl (1995) suggest that bracteoles are absent, Wood and Channell (1960) that they are present.
The calyx is sometimes inserted in two whorls, e.g. 3 + 2, 2 + 2, 2 + 3 in Chrysophylloideae (e.g. Swenson et al. 2023: Fig. 2), a derived feature. Penningon (2004) described the fruit of Eberhardtia as being a loculicidal capsule, but it appears to be a berry (e.g. S.-g. Li & Pennington 1996). Three genera do have capsular fruits, but there is only one seed, little or no endosperm, and they are African (Penningon 2004). Amyloid is also known from the seeds of Omphalocarpum, a clade that is close to sister to the rest of Chrysophylloideae (see Kooiman 1960).
For general information see Franceschi (1993), and Ng (1991), and especially Pennington (1991, 2004). For pollen, see Harley (1991), for the androecium, see Ronse De Craene and Bull-Hereñu (2016).
Phylogeny. Sarcosperma is sister to the rest of the family. Its seed has a shiny testa, albeit not as thick as that of most other Sapotaceae; the genus was placed in Sideroxyleae by Pennington (1991).Eberhardtia may well be sister to the remainder (Z.-D. Chen et al. 2016; Rose et al. 2018; Beckers et al. 2025).
Within the rest of the family there are two major clades, Isonandreae + Mimusopeae + Sideroxyleae], i.e. Sapotoideae, and [Chrysophylleae + Omphalocarpeae], i.e. Chrysophylloideae (e.g. Swenson & Anderberg 2005). These clades also appeared in a combined molecular + morphological analysis and after successive weighting, the latter still with only moderate support (79% jacknife) because of the inclusion of Xantolis, the rest of that clade minus Xantolis having 97% support, however, there was little in the way of well supported resolution of relationships beyond that, although Omphalocarpeae may be polyphyletic (Swenson & Anderberg 2005; see also Anderberg & Swenson 2003; Bartisch et al. 2010). Smedmark et al. (2006) provide a general discussion of relationships in Sapotoideae. However, in a study with a focus on Isonandreae (Sapotoideae), relationships are messier, and Eberhardtia was well separated from other Isonandreae and was sister to a clade that includes a Chrysophylloideae that is embedded in Sapotoideae, Sapotoideae-Sideroxyleae being sister to Chrysophylloideae (Richardson et al. 2013). In Phang et al. (2025) relationships were similar to those below, although such relationships were not the focus of that work. Beckers et al. (2025: c.f. Figs 1A, 1B) compared a plastome tree that was a composite of previously-published work with a tree that included 51 genera and was based on Angiosperms353 data; although the two subfamilies were found in both, there was extensive disagreement in generic-level relationships.
Sapotoideae. Christe et al. (2021) carried out analyses using 638 low-copy nuclear genes and with a focus on Sapotoideae; they recovered the relationships [Tseboneae [Inhambanella [Isonandreae + Sapoteae]]]; Inhambanella was separate from the three other genera of Gluemineae included - if these relationships hold up, two more tribes will be needed. In Boluda et al. (2022) relationships in the same area are [Isonandreae [Tseboneae [Inhambanella + Sapoteae]]].
- Within Isonandreae Richardson et al. (2013) found both substantial phylogenetic structure and substantial polyphyly, the latter perhaps paradoxically involving mostly members of smaller genera.
- Sapoteae. Phang et al. (2025: 95 species of Palaquium, 738 nuclear loci) found quite extensive generic-level polyphyly, although the bulk of Palaquium was holding together so long as Aulandra, a small genus, was included; Palaquium impressinervosum was sister to the rest of the genus (see also Phang et al. 2023). Randriarisoa et al. (2022) clarified relationships in Sapoteae-Manilkarinae.
- Sideroxyleae. For relationships in Sideroxylon, see Stride et al. (2014).
- Tseboneae. Boluda et al. (2022, see also 2024) found that analyses of 638 nuclear genes in Capurodendron confirmed the existence of ca 20 undescribed species; C. madagascariensis was very clearly sister to the rest of the genus.
Chrysophylloideae. Swenson et al. (2007a, 2008a, 2013) discuss generic limits in Australasian members of Chrysophylloideae; the whole lot are monophyletic, and Beccariella (= Pleioluma) is sister to the rest. In a tree of Chrysophylloideae from Asia and Oceania Magodendron may be sister to the rest (Swenson et al. 2020: support poor); Swenson et al. (2023) exmained relationships in Neotropical Chrysophylloideae in some detail. For relationships among the monophyletic group of the ca 80 species of the Pouteria complex on New Caledonia, see Bartish et al. (2005) - maybe three clades in Swenson et al. (2020). Pouteria sensu Pennington is polyphyletic (Triono et al. 2007), as is Chrysophyllum (Terra-Araujo et al. 2015), the former appearing in eight places in a tree of Neotropical Chrysophylloideae and the latter three times (De Faria et al. 2017). See Swenson et al. (2007b, esp. 2019) for Planchonella; the distinctive New Caledonian P. baillonii is sister to the rest of the genus. Swenson et al. (2008c) looked at relationships in the sister group of Planchonella, the largely New Caledonian Niemeyera complex. Borg et al. (2019) focussed on relationships in the Englerophytum/Synsepalum area, a clade from Africa; relationships along the spine of the tree had only weak support. The focus was the same in Potter and Uleh (2024: nuclear ITS and plastid trnH-psbA markers; species from the two genera were intimately mixed. Spiniluma is to be included in this subfamily (Stride et al. 2014). Swenson et al. (2023) returned to the fray here - 1 nuclear, 2 chondrome and 2 plastome markers, 122 terminals, Neotropical taxa - and proposed extensive reworking of generic limits. Swenson et al. (2025) revised Planchonella, the largest genus in Chrysophylloideae, and in their comprehensive phylogenetic analysis (184 terminals, 3 nuclear loci, indels) recovered four major clades, also two clades with but a single species, of which one, P. baillonii, from New Caledonia, was sister to the rest of the genus.
Classification. Generic limits have been notoriously fickle in Sapotaceae: "it is difficult to understand how two authors working on the same family could have come to such widely different conclusions" (Pennington 1990: p. 29). Pennington himself (1991) helped clarify things somewhat, but molecular data are providing much further information and generic limits continue to change, and will do so for the forseeable future (anno 2023). Thus the fate of the smaller genera in particular in Isonandreae (Sapotoideae) is unclear (see Richardson et al. 2013 for a phylogeny). For a classification of Chrysophylloideae in Southeast Asia/Oceania, see Swenson et al. (2013). Clade limits in e.g. New Caledonian Chrysophylloideae are also clear enough so that species can be described in their proper genera (e.g. Swenson et al. 2008b, esp. c), however, sampling in Neotropical Chrysophylloideae has to be improved before generic limits there can be addressed (De Faria et al. 2017). Borg et al. (2019) suggested that division of the Englerophytum/Synsepalum group into six genera was on the cards, i.e., generic limits would follow more A. Aubréville than T. D. Pennington, and Potter and Uleh (2024) inclined in the same direction, although the discussion there suggested lumping was a viable option, and mention of the polytomy there of itself has nothing necessarily to do with generic limits. In a recent study of Neotropical Chrysophylloideae, eleven genera were reinstated, Chrysophyllum and Pouteria were more narrowly circumscribed, yet much more was still to be done (Swenson et al. 2023) - note clade Q there, ca 100 species not yet assigned to genera, and some large genera in both Chrysophylloideae and Sapotoideae were polyphyletic.
For a checklist and bibliography, see Govaerts et al. (2001), but generic limits are dated. Swenson et al. (2020) provide a list of accepted - but not necessarily unproblematic - genera assigned to subfamilies.
EBENACEAE Gücke, nom. cons. - Back to Ericales
Bark and roots black; petiole bundle arcuate; sclereids +; leaves two-ranked, lamina margins entire, lower surface with flat glands; pedicels articulated; flowers imperfect, ?4-merous; K connate, C contorted, connate; staminate flowers: stamens adnate to corolla, in two series, basifixed, anthers long, connective prolonged, pistillode +; carpelate flowers: staminodes +; style ± divided; ovules 2/carpel, pendulous, apotropous; fruit a berry, K persistent/accrescent; testa vascularized; endosperm copious, hard, mannose-rich polysaccharides +, cotyledons foliacous, radicle long; x = 15, nuclear genome [1 C] (0.114-)1.169(-11.986) pg.
4/855: [list] - two subfamilies below. Tropical (to temperate).
Age. Crown-group Ebenaceae are estimated to be some (65-)54(-42) Ma (Turner et al. 2013a), ca 57.1 Ma (Rose et al. 2018) or (112.1-)106.5(-100.3) Ma (Bansal et al. 2021).
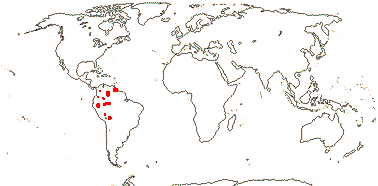 1. Lissocarpoideae Wallnöfer - Lissocarpa Bentham —— Synonymy: Lissocarpaceae Gilg, nom. cons.
1. Lissocarpoideae Wallnöfer - Lissocarpa Bentham —— Synonymy: Lissocarpaceae Gilg, nom. cons.
Trees; chemistry?; cork?; vessel elements with simple and scalariform perforation plates, mineral inclusions 0; (petiole bundle arcuate but with recurved edges and wing bundles); stomata anomocytic and cyclocytic; indumentum 0; plant dioecious; flowers axillary, or inflorescences subfasciculate; bracteoles large, apical, subopposite; flowers 4(-5)-merous; C right-contorted, corona +, 8-lobed, alternating with A, left-contorted [sect. Lissocarpa]; nectary?; staminate flowers: A 8, filaments basally connate; pollen 3-(4-)porate, ± smooth, 40-70 µm across; carpelate flowers: G [4], ovary inferior, [4], stigma clavate, hairy apically, hairs covering small lobes; ovules morphology?; seeds 1-2; testa?; endosperm very hard; cotyledons small; n = ?
1/8. Tropical N.W. South America. Map: from Wallnöfer (2004b: Fig. 4).
2. Ebenoideae Thorne & Reveal —— Synonymy: Diospyraceae Vest, Guaiacanaceae Jussieu, nom. illeg.
(Deciduous) trees; saponins, C-30 oxidised triterpenes, naphthoquinones [derivatives of 7-methyljugone and plumbagin], flavonols, leucodelphinidin, myricetin +, ellagic acid 0; vessel elements with simple perforation platesalone, silica bodies, prismatic crystals +; cambium storied; (cork cambium pericyclic); (nodes 1:3); SiO2 bodies + [not in Diospyros]; secretory cells common; petiole bundle also ± annular, with 0, 2 or more wing bundles; cuticle wax crystalloids 0; stomata usu. paracytic; hairs (T-shaped), unicellular; terminal bud aborts; leaves (opposite, spiral), lamina conduplicate, (margins slightly crenulate); inflorescence cymose, axis short; bracteoles alternate, ± median; plant di(mon)oecious, (flowers perfect), 3-7-merous; C urceolate to campanulate, rather small [usu. ≤1 cm long], (valvate); nectary +/0; staminate flowers: A (3-)12-20(-many), (inner anthers extrorse), often hairy; pollen 25.9±6.4 µm across, infratectum granular; pistillode?; carpelate flowers: (staminodes 0); G [2-8], opposite C or K, loculi often divided, stigmas little expanded, dry; ovules bitegmic, micropyle bi/endostomal, outer integument 3-7 cells across, inner integument 5-10 cells across [inner integument thicker than the outer]; K often accrescent; seed pachychalazal, often ruminate, testa multiplicative, radicle in pocket formed by testa (not - Diospyros), ± parenchymatous, or exotesta fibriform or mucilaginous, cells cuboid to palisade, endotesta crystalliferous or not, walls thickened or not; endosperm ± hard, cells thick-walled, cotyledons well developed; n = 15; nuclear genome [1 C] (1171-)1903(-2814) Mb; seedling phanerocotylar/cotyledons not photosynthetic, soon deciduous.
3/875: Diospyros (795+ - inc. 286 from Madagascar, 135 undescribed as of 10.vii.2024). Tropical (to temperate). Map: from Morley and Toelken (1983), Wickens (1976), White (1988) and Autralia's Virtual Herbarium (consulted iii.2014). Photos: Carpellate flower, Fruit, Collection.
Age. Crown-group Ebenoideae have been dated to (50-)42(-35) Ma (Turner et al. 2013a), ca 38 Ma (Rose et al. 2018), (57.8-)56.2(-55.1) Ma (Linan et al. 2019) or close to twice these ages, (108.6-)101.6(-94.0) Ma (Bansal et al. 2021).
Bansal et al. (2021: beautiful images) examined pollen found in two lignite mines in W.-N.W. India dated to ca 63 and 55-52 Ma and placed samples of this pollen found in the earlier deposits in all three genera of Diospyroideae. Along the same lines, fossil pollen from western Europe had suggested that diversification in Diospyroideae had begun by ca 56 Ma (Hofmann 2018).
Fossil flowers and associated leaves (Austrodiospyros) are known from Middle Eocene rocks of southeastern Australia ca 33.9 Ma (Basinger & Christophel 1985; Martínez-Millán 2010). For more on the fossil record of Diospyroideae - the identity of early macrofossils is suspect - see Bansal et al. (2021).
Evolution: Divergence & Distribution. Bansal et al. (2021) suggested that Ebenoideae might ancestrally have been in equatorial Africa, then they moved on to the Indian plate as it passed via the Kohistan—Ladakh Arc, and thence on to Southeast Asia 50-40 Ma - see also Arecaceae and Dipterocarpaceae. Although fossils identified as Euclea and Royena are reported from these Indian localities, those genera are today restricted to Africa (Bansal et al. 2021 - see also Euphorbiaceae—Crotonoideae).
Duangjai et al. (2009) found four separate lineages of Diospyros in New Caledonia, Turner et al. (2013a, b) noting that speciation there has occurred within the last 10 Ma. Resolution of relationships between species was often difficult, even when looking at a combination of whole plastid genomes and ribosomal DNA (Turner et al. 2016). In the largest of these clades, with some 24 species, initial divergence seems to have been mediated by climate or other features affecting the macrohabitat, but latterly differences in substrate/soils triggered speciation (Paun et al. 2015; also Samuel et al. 2019: summary. There are some 240 species of Diospyros on Madagascar (P. P. Lowry, pers. comm. 2019, ca 2/5 the species described), although little is known about their relationships; there are at least two separate clades on the island, and also two clades on the Mascarenes, and both the latter and some clades on the African mainland have originated in Madagascar (see also Buerki et al. 2013; Linan et al. 2019). Diospyros is about the most diverse genus in West Malesian l.t.r.f. (Davies et al. 2005).
Geeraerts et al. (2009) suggested apomorphies - especially palynological - for genera of Ebenoideae. The chemistry of Lissocarpoideae is unknown, so whether the presence of naphthoquinones is a synapomorphy of Ebenaceae as a whole or just part of them remains to be established.
Pollination Biology & Seed Dispersal. Ebenaceae-Euclea has been colonized twice by derelomine weevils which probably moved on to the genus from palms; brood-site pollination mutualisms are involved here (Haran et al. 2021).
Plant-Animal Interactions. Extrafloral nectaries occur widely in Ebenaceae (Weber & Keeler 2013).
Genes & Genomes. Akagi and Charlesworth (2019) discuss sex-determining genes in Diospyros and their evolution (see also Basinger & Christophel 1985; Henry et al. 2018). There seems to have been dioecy here for over 50 Ma, and it may be determined by a single-factor model. Epigenetic factors may allow the development of monoecy, and there is gynodioecy in the hexaploid D. kaki.
There is extensive variation in genome size in Diospyros, even within New Caledonia (Samuel et al. 2019).
Chemistry, Morphology, etc.. Ellagic acid may occur in Ebenaceae-Ebenoideae (Bate Smith 1962). Naphthoquinones are likely in Lissocarpa, given that the outer bark and roots are blackish. Vessels sometimes occur in radial multiples in the wood. See Sandratriniaina et al. (2023) for petiole anatomy, etc., of some species from Madagascar. Both Massart's model (rhythmic monopodial branches) and variants (e.g. Roux - continuous branching) occur in Diospyros. The terminal bud of the innovation frequently aborts. The flat foliar glands are extrafloral nectaries and they are quite often closely associated with vascular bundles; they probably occur in about three quarters of the species in the family (Contreras & Lersten 1984).
The morphology of the inflorescence of Lissocarpoideae is unclear; one interpretation is that the flowers are axillary, whether on short or long shoots. The flowers seem to be imperfect, at least some "perfect" flowers not producing fertile pollen despite having well developed anthers (Wallnöfer 2004a). In Diospyros s.l., both integuments appear to be very thick, although the inner is only three cells across at the endostome (van Tieghem 1898). There is variation in germination - foliaceous cotyledons and alternate subsequent leaves vs thick cotyledons and opposite leaves.
For general information, see Franceschi (1993), Ng (1991), and Wallnöfer (2001); for wood anatomy, see Jahanbanifard et al. (2020), for carpel orientation, see Baillon (1891), Eichler (1875) and Le Maout and Decaisne (1868), for some seed anatomy, see Quisumbing (1925). Some information on Lissocarpa is taken from Schadel (1978: leaf morphology) and Wallnöfer (2004a, b), but that genus is poorly known.
Phylogeny. Rather degraded rbcL sequences initially suggested that Lissocarpa was to be included in Rutaceae, in Sapindales (Savolainen et al. 2000a). However, it is well supported (rbcL only) as sister to Ebenaceae s. str. (e.g. Berry et al. 2001; Rose et al. 2018).
Duangjai et al. (2006a and especially b), sequencing six plastid genes, found extensive phylogenetic structure in Ebenoideae: the African(-Arabian) Euclea and Royena were sister to Diospyros, and within Diospyros there were a number of well-supported clades, although relationships between them were unclear. Duangjai et al. (2009: eight genes, 119 species) provided a more detailed phylogeny of Diospyros s. str. with good Bayesian support for relationships along the backbone of the tree. For relationships focussing on the Mascarene taxa, see Linan et al. (2019).
Classification. Lissocarpaceae have often been placed in or close to Ebenaceae, but they were unassigned in A.P.G. I (1998). Since the two have morphologically much in common, it is reasonable to combine them (see A.P.G. III 2009).
PRIMULACEAE Borkhausen, nom. cons. - Back to Ericales
Schizogenous secretory glands/canals throughout the plant [contents yellow, red, brown: tannins, etc.]); saponins common, ellagic acid 0; rays 1-3 cells across; nodes ?3:3; trichomes peltate; stomata anomocytic; inflorescence racemose; C and A from common primordia; C connate; A = C, adnate to and opposite C, staminodes +; nectariferous tissue on G; G [5], opposite C, placentation free-central, with sterile apical projection/not, style short, hollow, stigma usu. ± capitate; ovules apotropous, bitegmic, micropyle bistomal, outer integument ca 2 cells across, inner integument 3-4 cells across [inner integument thicker than outer], endothelium +, tanniniferous; seeds angled; seed coat 2-layered, inner layer crystalliferous; endosperm nuclear, copious, cell walls thick, pitted [amyloid], with xyloglucans; x = 12 (?11), nuclear genome [1 C] (0.146-)1.895(-24.564) pg.
58/2,615: [list, to tribes] - four subfamilies below. World wide.
Age. Wikström et al. (2001) suggested a crown-group age for Primulaceae of around (52-)49, 46(-43) Ma; this age is estimated to be (74-)61, 57(-45) Ma by Bell et al. (2010), ca 79.5 Ma by Rose et al. (2018), (77.7-)77.1(-76.3) Ma by Larson et al. (2023) and 80.2-78.5 Ma by H.-F. Yan et al. (2024).
1. Maesoideae de Candolle - Maesa Forsskål —— Synonymy: Maesaceae Anderberg, B. Ståhl & Kallersjö
Lianes, shrubs or trees; (vessel elements with scalariform perforation plates); hairs unicellular, also peltate; petiole bundles three, annular; ?stomata; leaves spiral or two-ranked, lamina vernation induplicate, margin toothed to entire [also with papillae]; inflorescence often branched; bracteoles apical (connate); flowers small [7> mm across], (4-merous); K imbricate, C urceolate, induplicate-valvate/quincuncial/imbricate, developing before A; A basally connate, adnate to C at middle; G [(3-4)5], semi-inferior, style hollow?, stigma truncate, capitate or 3-5-lobed; ovules partly/not embedded in placenta (uniseriate); fruit many-seeded, drupaceous/baccate, K persistent; seed ?hilum; n = 10.
1/192. Old World tropics to Japan, the Pacific, and Australia. Map: from Coates Palgrave (2002).
Age. The age of crown-group Maesa may be ca 10.2 Ma (Rose et al. 2018: ?sampling).
[Theophrastoideae [Primuloideae + Myrsinoideae]]: bracteoles 0; C imbricate, subrotate, developing after A; ovules not embedded in placenta; hilum sublinear to elliptic.
Age. The age of this node is estimated to be 42-40 Ma by Wikström et al. (2001), (67-)55, 51(-40) Ma by Bell et al. (2010), (73-)55(-35) Ma by Wikström et al. (2015), about 65 Ma by K. Bremer et al. (2004a), ca 72.1 Ma by Rose et al. (2018) and ca 69.7 Ma by Larson et al. (2023).
2. Theophrastoideae A. de Candolle
Bracts ± displaced up the pedicels [recaulescent]; trichomes on K and C; staminodes ± C-like, opposite K; endothelium?
6-9/105. Mostly New World and tropical, some also more temperate and Old World.
Age. Crown-group Theophrastoideae are ca 70 Ma (Rose et al. 2018), ca 65.6 Ma (Larson et al. 2023) or 61.4-56.5 Ma (H.-F. Yan et al. 2024).
2A. Samoleae Reichenbach - Samolus L. —— Synonymy: Samolaceae Rafinesque
Herbs (annual) to subshrubs; nodes 1:1; leaves (reduced), lamina margins entire; (inflorescence scapose); flowers small [7> mm across]; K connate, C rotate; A adnate to C at middle, (staminodes 0), (anthers with prolonged connective); nectary at base of G; G [5], semi-inferior, style impressed; ovules not immersed in placenta, inner integument 2-3 cells across, innermost layer ± palisade; fruit a capsule with 5 teeth; seeds many, angled, small [ca 0.5 mm long]; coat undistinguished, exotesta and endotegmen tanniniferous, the latter crystalliferous; endosperm cell walls thin to slightly thickened; n = (12) 13.
1/15. America, the Antipodes, Europe, tropical to temperate. Map: from Hultén (1971), Meusel et al. (1978), and FloraBase (2005), red is Samolus valerandi only, for which see Wanntorp and Anderberg (2011). [Photo - Flowers.]
Age. Friis et al. (2021) described the flowers of Miranthus from Campanian-Maastrichtian deposits in west Portugal dated to perhaps ca 72 Ma. These flowers have many of the features of those of Samolus, and an association with that genus was most parsimonius when the fossil was placed on a molecular backbone tree, although other positions in Primulaceae were not much less parsimonious (Friis et al. 2021).
Crown-group Samolus are ca 34.1 Ma (H.-F. Yan et al. 2024: c.f. sampling).
2B. Theophrasteae Bartling —— Synonymy: Theophrastaceae G. Don, nom. cons.
Woody, tending to be pachycaul, (Schopfbaum); rays ≥10 cells across; nodes also 1:1 [Jacquinia, dividing into three] 5:5 [Clavija]; glands, canals?; petiole bundle deeply arcuate or annular, with small adaxial inverted bundles; subepidermal fibres/bundles +, marginal sclerenchyma; perulae +; lamina vernation conduplicate, margins spiny-toothed to entire, apex pungent or not, plant dioecious or flowers bisexual; flowers medium-sized; C development and presence of A-C common primordia unclear; anthers extrorse, with calcium oxalate crystals, etc. at apex and base, initially incurved over stigma; ?nectariferous hairs on K/C (0); G?, style long, stigma dry or wet; outer integument 2-4 cells across, ?inner; fruit a (rather dry) berry, (drupaceous), placentae ± pulpy; seeds 1-few, rounded; testa multiplicative, exotestal cells flattened, thick-walled, (hypodermal cells with thickened anticlinal walls), other mesotestal cells crystalliferous; cotyledons usu. foliaceous; n = 18, 20, 24.
4/90: Clavija (50). New World tropics. Map: from Ståhl (1989, 1991, 1995). [Photos - Collection.]
Age. The age of this clade is estimated to be ca 22.3 Ma by Larson et al. (2023) or ca 16.1 Ma by H.-F. Yan et al. (2024).
[Primuloideae + Myrsinoideae]: herbs; distinctive triterpene saponin; tapetal cells uninucleate; (exotegmic cells elongated, with band-like thickenings); fruit a capsule, dehiscing by apical valves; two ndhF deletions.
Age. This node may be ca 57.6 Ma (Rose et al. 2018), (54-)44, 40(-30) Ma (Bell et al. 2010: note topology), ca 41.7 Ma (Fu et al. 2019), (50-)32(-16) Ma (Wikström et al. 2015), ca 42.6 Ma (X. Feng et al. 2020/2021), ca 58.2 Ma (Larson et al. 2023) or ca 55.2-51.5 Ma (H.-F. Yan et al. 2024).
3. Primuloideae Kostelesky
Perennial (annual) herbs; cucurbitacins +; ?cork; ray width?; (cork cambium deep seated); vascular bundles usu. separated; glands/canals ?often 0; trichomes articulated, plant (farinose); lamina vernation involute ("induplicate") or revolute, margins entire to dentate or serrate, (pinnatipartite); inflorescence scapose; (bracts 0); flowers medium-sized; K often connate, (one vascular trace/K member), C salverform, (lobes retuse/± serrate/deeply lobed); A attached at or above middle of C tube; pollen syn- or polycolpate/pantoporate, or tricolporate; style usu. long, stigma dry; fruit with 5, 10 apical valves, (apically circumscissile); seeds many, angled, (winged); exotesta ± persistent, walls thickened or not, (endotegmen crystalliferous); endosperm (cell walls somewhat thickened, thin); n = 8-12; nuclear genome [1 C] (460-)2777(-10352) Mb [?level].
9/900. Overwhelmingly from the northern hemisphere, scattered elsewhere. Map: from Hultén (1971) and Meusel et al. (1978). Photo: Dodecatheon flower © R. Kowal, Primula flower.
Age. The age of crown-group Primuloideae is around 51 (Rose et al. 2018), ca 37.5 (Strijk et al. 2014), ca 35.1 (Fu et al. 2019), 33 (Boucher et al. 2016a), 54.9 (Larson et al. 2023) or ca 48.5/ca 45.1 Ma (H.-F. Yan et al. 2024: Stimpsonia not taken into account in the second age).
3A. Androsaceeae Pax - Androsace L.
(Annuals; cushion plants); leaves (pinnatifid - A. filicula); C with ring appendages at throat; pollen surface perforate/rugate/microreticulate; (integument single, 6-10 cells across - Douglasia); (capsules circumscissile - A. fil.); n = 9, 10, 11, 15, 16...
1/166. Temperate-Boreal Eurasia (the annual A. umbellate in New Guinea), western North America, southern South America.
Age. Crown-group Androsaceeae are thought to be (35.3-)34.9(-34.5) Ma (Boucher et al. 2011), ca 47.3 Ma (Larson et al. 2023) or ca 33.2 Ma (H.-F. Yan et al. 2024).
[Hottonieae + Primuleae]: (nectary 0).
Age. The age of this clade is ca 50.5 Ma (Larson et al. 2023) or ca 39.1 Ma (H.-F. Yan et al. 2024).
3B. Hottonieae Endlicher
(Plant aquatic - Hott.); leaves (± whorled, lamina deeply divided - H.), (brown-punctate - Omph.); C (deeply lobed - Sol.), (staminodes + - Sol.); seeds reticulate; n = 10, 11, 19, 20, 24.
4/30: Soldanella (15), Omphalogramma (12). North Temperate.
Age. Ca 33.3 Ma is the age of crown-group Hottonieae (Larson et al. 2023), while H.-F. Yan et al. (2024) suggest an age of ca 25.1 Ma.
3C. Primuleae Dumortier
(Cushion plants, ± woody - Dionysia); (plant aromatic - D.); stem with endodermis; (foliar sclereids +); vernation various; heterostyly + (0); (ovules immersed in placenta - D.); endotesta with inner walls thickened; n = 9-10, (polyploids +).
4/622: Primula s.l. (553), Dionysia (61). Northern Hemisphere, esp. the Himalayas, to New Guinea, Ethiopia and the Horn of Africa, temperate South America (P. magellanica).
Age. Larson et al. (2023) suggested that the age of this clade is ca 45.5 Ma while the age in H.-F. Yan et al. (2024) is ca 32.8 Ma.
4. Myrsinoideae Burnett
Benzoquinones +; (vessel elements with scalariform perforation plates); wood rays 0 / multiseriate, (with breakdown areas); (cortical bundles +); glands/canals + (0); inner wall of epidermis ± mucilaginous/0, (mesophyll cells with invaginations); lamina also vernation supervolute (curved), margins entire/crenate to serrate, teeth cartilaginous; inflorescence often fasciculate/corymbose; flowers (3-)4-5(-7)-merous, small to medium-sized; (bracts foliaceous); flowers protogynous [?sampling]; C often right-contorted [?level], (developing before A), tube short, (margins ciliate); nectary 0; A with anthers dorsifixed or basifixed, sagittate, (dehiscing by pores), staminodes 0; (pollen loxocolporate [apertures oblique]); style (0), (stylar canal 0), stigma dry or wet; ovules immersed in placenta, outer integument 2-3 cells across, inner integument 2-3 cells across, endothelium + (0), parietal tissue ca 4 cells across; seed coat undistinguished, testa ?and tegmen multiplicative, becoming crushed; endosperm cell wall thickening variable; embryo suspensor massive.
41/1,435. Pantropical and N. Temperate. Map: from Hultén (1958, 1971) and FloraBase 2008: Southern Hemisphere a bit notional. [Photos: collection woody members, Cyclamen flower © H. Schneider, fruit © H. Schneider, collection of ex-Primulaceae.]
Age. Myrsinoideae began radiating ca 53.1 Ma (Rose et al. 2018), other estimates are 56.6 Ma (Larson et al. 2023) and 40.3 Ma (H.-F. Yan et al. 2024, Coris but not Stimpsonia included).
4A. Stimpsonia chamaedryadoides A. Gray
Herb, annual/biennial; lamina irregularly dentate; inflorescence terminal, racemose, bracts foliose; K valvate, C imbricate, lobes emarginate or not; G [5], stigma capitate; ovules many, micropyle exostomal; fruit a 5-valved capsule; n = ?
1/1. S. China, Taiwan and Japan.
[Corideae [[Ardisiandreae [Cyclamineae + Lysimachieae]] [Woody Myrsinoideae]]]: ?
Age. The age of this clade is ca 56.4 Ma (Larson et al. 2023).
4B. Corideae Dumortier - Coris L. —— Synonymy: Coridaceae J. Agardh
Ericoid sub-shrub (annual); wood rayless; inflorescence terminal, congested, ?racemose; flowers monosymmetric, inverted; epicalyx +, spine-tipped; C unilabiate [5:0], lobes bilobed; ovules usu. 1/carpel, embedded in placenta, micropyle endostomal; fruit with 5 apical valves; seeds papillate; n = ?
1/2. Central and western Mediterranean, N.W. Africa, the Sudan.
[[Ardisiandreae [Cyclamineae + Lysimachieae]] [Woody Myrsinoideae]]: ?
Age. This clade is around 53.1 Ma (Larson et al. 2023) or ca 31.3 Ma (H.-F. Yan et al. 2024, but c.f. topology).
[Ardisiandreae [Cyclamineae + Lysimachieae]]: ?
Age. This clade is 52.0 Ma (Larson et al. 2023).
4C. Ardisiandreae Schwarz - Ardisiandra J. D. Hooker
Herbs, perennial; hairs articulated [uniseriate-glandular/]; leaf vernation revolute, lamina ± dentate; inflorescence terminal, flowers 1-few in axils of foliaceous bracts; K valvate, C quincuncial, lobes ± rounded; anthers with connective protruding, forming a cone; stigma subcapitate, strongly papillate; ovules many, initially not immersed in placenta, exostomal; capsule dehiscing irregularly, wall translucent/with valves, wall lignified; seeds papillate; n = 17.
1/3. East African mountains.
Age. Larson et al. (2023) suggested an age of ca 7.8 Ma here.
[Cyclamineae + Lysimachieae]: ovules embedded in placenta.
Age. The age of this clade is ca 50.5 Ma (Larson et al. 2023) or a mere 26.8 Ma (H.-F. Yan et al. 2024).
4D. Cyclamineae Dumortier - Cyclamen L.
Herbs, tuberous [tuber hypocotylar]; stem pericycle not lignified; petiole with annular bundle; ?stomata; leaves radicular, lamina with palmate venation, base cordate; flowers large [ca ≥2 cm long], axillary, long-pedicelate; C developing before A, reflexed, epidermal cells not elongated, anticlinal walls straight or somewhat sinuous; anthers porose, usu. shorter than filaments, adnate to C; nectary and gland-headed hairs 0; style hollow, style punctate, stigma inside hollow tip; ovules campylotropous, integument 1, ca 2 cells across, inner integument ca 7 cells across, becoming digested, parietal tissue ca 4 cells across; pedicel coiling or otherwise bringing capsule close to the ground; capsule 2-layered; seeds relatively large [ca 2.8 mm long], covered in goo, hilum concave; endotesta thickening U-shaped; endosperm with thick and pitted walls, perhaps hemicellulosic, embryo slightly curved, ± transverse in seed, medium, cotyledon single; n = 10, 11, 15, 17.
1/23. Southern Europe and the Mediterranean Basin, to Iran, N.E. Africa (Somalia, 1 sp.).
Age. Perhaps ca 13.0 Ma (Larson et al. 2023) or ca 8.0 Ma (H.-F. Yan et al. 2024), but c.f. sampling in both cases.
4E. Lysimachieae Reichenbach —— Synonymy: Anagallidaceae Borkhausen, Lysimachiaceae Jussieu
Perennial (annual) herbs, (shrubs, e.g. Hawaiian Lysimachia); stem pericycle lignified; leaves (opposite), lamina (succulent); (heterosyly +); K basally connate, C (0), epidermal cells elongated, anticlinal walls sinuous; A basally connate or not, free or basally adnate to C (adnate the length of tube), anthers (forming a cone), (porose), shorter than filaments; pollen (4-colporate); oil flowers + (0), gland-headed hairs on G; (antipodal cells large - Lysimachia); capsule with apical valves, (circumscissile), disintegrating irregularly; seeds many, ± angled; n = 8, 10-12, 14, 15...
5/250: Lysimachia (180). Widely scattered, inc. Hawaii, esp. China, few in South America, Australia or Africa.
Age. Crown-group Lysimachieae are ca 47.1 Ma (Larson et al. 2023)
Shrubs to trees, (pachycaul); petiole bundle (incurved) arcuate/interrupted annular, stomata also paracytic [Cybianthus], anisocytic; lamina (with nodules), (vernation revolute - Ardisiandra); (plant dioecious); flowers 4-5(-7)-merous; K deeply lobed (± connate / free), C developing before A, right-contorted; anthers sagittate, (cone-forming, connate by thecal margins), (locellate, shorter than filaments - Ardisia); pollen (4-colporate); nectary 0; stigma punctate; fruit a drupe; seed 1, (subglobose), hilum depressed; testa 1-layered, crystals 0; endosperm +, (ruminate); n = 23.
23/1,528 or 30/1,870: Ardisia (734: polyphyletic), Discocalyx (147: inc. Tapeinosperma [87]), Oncostemum (100) - see Julius et al. (2021), Stylogyne (60). Tropics, inc. Pacific islands.
Age. The crown-group age of Woody Myrsinoideae is ca 32.6 Ma (Larson et al. 2023) or ca 16.2 Ma (H.-F. Yan et al. 2024).
4F1. Aegicerateae A. de Candolle —— Synonymy: Aegicerataceae Blume
Shrub to tree; vascular rays relatively narrow [A.]; lamina margins entire; plant dioecious; K and C left contorted [A.]; C imbricate, tube very short; filaments very short, anthers porose; G with gland-headed nectariferous hairs [A.], style ± 0; ovules numerous, unitegmic [A.], integument 4-5 cells across [A.], funicular vascular bundle 0 [A.]; fruits long, curved [A.], seed 1/fruit [A.]; endosperm 0 [A.], ruminate, embryo to 8 cm long [A.]; n = 23 [A.], nuclear genome [1 C] ca 841 Mb [A.]; plant viviparous [A.].
2/10: Monoporus (8). Madagascar (Monoporus), Aegiceras coasts and islands India and S. China to N.E. Australia and the Pacific.
Age. Crown-group Aegicerateae are ca 29.1 Ma (Larson et al. 2023).
[Embelieae [[Ardisia Old World] [[Elingamita + Tapeinosperma] [Ardisia New World]]]] / [Myrsinoids + Ardisioids]: G/C/filaments with gland-headed nectariferous hairs (0 - Ardisia, Myrsine).
Age. This clade is ca 28.7 Ma (Larson et al. 2023).
4F2. Embelieae Bartling / Myrsinoids / [Embelia / [Cybianthus [Myrsine [Pleiomeris + Heberdenia]]]] —— Embeliaceae J. Agardh, Myrsinaceae R. Brown, nom. cons.
Liane, shrub, tree; plant dioecious/flowers perfect; C valvate/imbrictae; stigma various, often ± expanded; endosperm ruminate/not.
Myrsine (304: inc. Rapanea, Suttonia, etc. - polyphyletic), Cybianthus (160), Embelia (139). Pantropical (warm temperate).
Age. An estimate for the age of Embelieae is ca 26.2 Ma (Larson et al. 2023).
Ardisioids / [[Ardisia Old World] [[Elingamita + Tapeinosperma] [Ardisia New World]]]: ?
Age. An estimate for the crown-group age of the ardisioids is (26.1-)24.7(-23.2) Ma (Larson et al. 2023).
Age. This clade is (24.8-)23.5(-21.5) Ma (Larson et al. 2023.
[Elingamita + Tapeinosperma] [Ardisia New World]: ?
Age. Larson et al. (2023) suggested that this clade started diverging ca 22.5 Ma.
Age. Larson et al. (2023) suggested that these two genera diverged ca 19.0 Ma.
4. Ardisia New World / Ardisieae Bartling —— Synonymy: Ardisiaceae Jussieu
Parathesis (85).
Age. An estimate for the crown-group age of this clade is (21.0-)20.0(-18.6) Ma (Larson et al. 2023).
Evolution: Divergence & Distribution. For clade ages throughout the Primulaceae, see Larson et al. (2023: esp. Fig. S9); these authors also discuss distributions of some of the groups here.
Wanntorp et al. (2012) discuss the evolution of a number of characters of floral development in Primulaceae. Although some taxa have glands of various colours throughout the plant it is unclear exactly where this character should be placed on the tree. Similarly, where the character "ovules embedded in the placenta" should be placed is uncertain; the placenta sometimes seems to envelop the ovules only after fertilization, as suggested by the image of the fruits of Stimpsonia (Myrsinoideae) in Wanntorp et al. (2012), but embedded ovules are usually absent in Primuloideae and Samoleae, at least.
Anderberg et al. (2001) thought that rosette herbs might be the plesiomorphic condition for the [Theophrastoideae [Primuloideae + Myrsinoideae]] clade. However, Lens et al. (2005a) found no evidence from wood anatomy that this was likely, apart from in a few Myrsinoideae. Many of these herbaceous taxa have capsular fruits with five apical teeth (see also Larson et al. 2023), presumably plesiomorphic, and herbaceous Myrsinoideae such as Stimpsonia, Ardisiandra and Coris are more basal on the tree than woody taxa. A bit complicated. Similarly, it has been suggested that a cauline endodermis is a feature of Primulaceae in the old sense (Seago 2020 for literature), but it is unclear what group this feature actually characterises. Indeed, Decrock (1901) mentioned a cauline endodermis in Samolus (Primulaceae in the old sense), so a comprehensive survey of endodermal-type structures in Primulaceae might be useful.
Morozowska et al. (2020) optimized a number of seed and fruit characters on a tree of the family; Maesa was not scored. In many taxa the "seed coat" consists of two layers, an outer more or less thickened layer and an inner crystalliferous layer, however, the origin of this latter seems to vary (Morozowska et al. 2011, 2020), hence its apparently somewhat redundant mentions in the characterizations above. Even in taxa which appear to have similar origins of the testa it is anywhere from 9.9 to 128.6 µm across (Morozowska et al. 2011). See also Oh et al. (2008) for the seed morphology of herbaceous taxa around Lysimachia. The family would repay a comprehensive survey of fruit and seed characters.
Species of Samolus from S.W. North America are sister to the rest of the genus, and Theophrasteae are a tropical New World clade (Wanntorp & Anderberg 2011), perhaps suggesting a New World origin for Samoleae. Rose et al. (2018) suggested a New World origin for Theophrastoideae, occurrences outside the Neotropics being more recent. The recent discovery of ca 72 Ma fossils of Miranthus, from Portugal and close to Samolus (Friis et al. 2021), make one think twice; interestingly, the sediments in which Miranthus was found had marine connections, suggesting that the plant was growing in a habitat like that of extant Samolus (Friis et al. 2021).
Primula, Androsace and Soldanella have all radiated in alpine habitats in Europe where allopatric speciation has been common, but the first two also grow elsewhere in the northern hemisphere, Androsace being notably diverse in the Himalayas while Southwestern China and adjacent regions harbour the bulk of the diversity of the speciose Primula itself (Comes & Kadereit 2003; L.-B. Zhang et al. 2004; Boucher et al. 2011, 2016a; Roquet et al. 2013; Hughes & Atchison 2015; Y.-J. Liu et al. 2015 and references). There has also been extensive dispersal between these isolated alpine areas (e.g. Boucher et al. 2011; Roquet et al. 2013), while P. magellanica, from southern South America, grows far to the south of other Primuleae, species in Java perhaps being the closest; nearly all species are north of the equator. There has been notably extensive hybridization in Soldanella, the plastome relationships reflecting neither taxonomy, morphology nor ecology - but some geography (Slovák et al. 2023). Boucher et al. (2011) suggest that the cushion plant habit may be a key innovation in Androsace - it evolved twice there - and the short-lived/annual habit may be plesiomorphic in the genus.
Within Myrsinoideae, the rare, monotypic Pleiomeris (P. canariensis), restricted to the Canary Islands, is sister to Heberdenia, which also grows on the Azores; both are shrubs; although the east-Asian Stimpsonia may also be close (Martins et al. 2003), the latter is a herb with very different flowers and inflorescence. Note that Palaeo-Macaronesia is likely to be 60 Ma or more old, the oldest currently emergent Canary Island dating to ca 21 Ma (Gelmacher et al. 2005; Fernández-Palacios et al. 2011). However, Appelhans et al. (2020) found that Pleiomeris and Heberdenia (they did not look at Stimpsonia) may be separately embedded in Myrsine, and this also contains an Australasian-Polynesian clade (and also some species from the Americas) within which the ca 20 species of Myrsine from Hawai'i form a clade. There has been extensive hybridization in these Hawaiian Myrsine (Appelhans et al. 2020 and references). Strijk et al. (2014) discussed the evolution of the Madagascan and Mascarene Oncospermun-Badula complex. Much speciation there is quite recent, but Badula on Rodrigues may be older than the island.
Lysimachieae in Eastern Asia are quite diverse, and this is ascribed to a combination of how long the clade has been there and high diversification rates (Yan et al. 2018).
Ecology & Physiology. Aegiceras (Myrsinoideae) is restricted to the mangrove habitat; its seeds tend to sink quite quickly (Clarke et al. 2001). Hardly surprisingly, Aegiceras has a number of anomalous anatomical and morphological features; X. Feng et al. (2020/2021) discuss the molecular basis of salt tolerance, etc., here, noting that there has been a genome duplication in this area. The seeds in particular are those that might be expected from a mangrove plant; they lack endosperm and contain a large embryo that breaks the seed coat before the seed falls from the tree (c.f. Rhizophoraceae-Rhizophoreae, Acanthaceae-Acantheae-Acanthus ilicifolius, etc.: Juncosa 1982). For more on the evolution of the mangrove habitat, see Rhizophoraceae.
Boucher et al. (2011; see also Roquet et al. 2013) discuss the evolution of life forms in Androsace (this includes the North American Douglasia, etc.), which turns out to be very labile in this respect. The plesiomorphic condition may initially have been annuals/short-lived perennials, the perennial cushion habit has evolved twice in alpine habitats since the Miocene, and it is perhaps a "key innovation" enabling life at high altitudes - cold and dry; note that the crown-group age of Androsace is over 35 Ma (Boucher et al. 2011), although some estimates are slightly younger or decidedly older (see above). Dionysia (= Primula s.l.) are also mostly cushion-forming plants, often with a woody stem, and some are chasmophytes (Trift et al. 2004; Lidén & Mehregan 2023). With over 60 species of cushion plants, Primuloideae include a disproportionally large number of them (Boucher et al. 2016b).
Zona and Christenhusz (2015) note that litter-collecting Schopfbaum are known from genera like Clavija (Theophrasteae).
Pollination Biology & Seed Dispersal. As will be clear, there is a considerable amount of variation in how nectar and other floral secretions are produced in Primulaceae, and this is described in detail by Cano and Ronse de Craene (2023). They document nectarstomata on the gynoecium (and sometimes base of the style) in all subfamilies, but only in basal Myrsinoideae. Capitate-glandular hairs are particularly common in Theophrastoideae (their function there is unclear) and in Myrsinoideae that do not have nectarostomata, although not in Ardisia and Myrsine.
Heterostyly is well known in Primula (e.g. Darwin 1877). Although common, it is unlikely to be an apomorphy for Primuloideae, however, it may well be an apomorphy for Primula s.l. (de Vos et al. 2014a). Heterostyly may reduce the chances of the extinction of older clades, at least over shorter time spans, homostylous clades may show accelerated diversification (de Vos et al. 2014b). Mora-Carrera et al. (2025) noted that flowers of diplpod plants were distylous, heteromorphic and self incompatible, while those of polyploids were homostylous, homomorphic and self compatible. Primula section Primula contains European species that have been subjects of many of the studies on heterostyly; relationships in this isolated section are complex and species limits unclear (Schmidt-Lebuhn et al. 2012). Thrum plants are heterozygous Ss, while pin plants are the homozygous recessive, ss, S being a supergene consisting of three (or five?) tightly-linked genes (e.g. Li et al. 2011; Cocker at al. 2018; Gutiérrez-Valencia et al. 2021; Potente et al. 2022), however, Cocker at al. (2018) accurately describe the thrum plants as being hemizygous. Heterostyly is sometimes lost, as in species of Primula with buzz pollination (Mast et al. 2001, 2006), and in a number of other small clades, e.g. genera like Soldanella have the anthers in a cone surrounding the style (de Vos et al. 2014a, b). For further details, see McCubbin (2008), Barrett and Shore (2008), Kappel et al. (2017) and Cohen (2019).
The speciose Ardisia has flowers that look as if they are buzz pollinated; there are no nectaries. Indeed, Otegui and Cocucci (1999) note the possibility of buzz pollination here, a vibrating tuning fork being able to empty the anthers of pollen, and buzz pollination has been observed in A. escallonioides (Pascarella 1997). Similarly, the erstwhile genus Dodecatheon with its distinctive floral morphology is from the point of view of evolution a buzz-pollinated Primula (Mast et al. 2004; Harder & Barclay 1994, see also Lysimachia below). Interestingly, although the flowers of Cyclamen also have the buzz-pollination syndrome, buzz pollination has only rarely been observed there, and in C. persicum, at least, the micropterid jawed moth Micropterix elegans is the main pollinator and appears to be restricted to that species of Cyclamen (Schwartz-Tzachor et al. 2006; Nevard et al. 2021). For more on buzz pollination, see elsewhere.
Vogel (1986, 1997) examined pollination in Lysimachia, a largely herbaceous group with a few woody species. Pollination of about 70 or more species with yellow flowers (the colour is actually bee-green) is by 16 species of Macropis (Mellitidae-Macropodini) oil bees (Michez & Patiny 2005: see also Simpson et al. 1983; Michez et al. 2008; Renner & Schaefer 2010: "co-evolution"; Schäffler et al. 2012; Possobom & Machado 2017 and references). The bees collect oil that is secreted by floral trichomes, as well as pollen; they pbtain nectar from a range of flowers (Renner & Schaefer 2010). There are also buzz-pollinated species, while white-flowered species have nectariferous hairs, which suggests a different pollination mechanism, perhaps generalist bees collecting nectar and pollen. Oil pollination has been lost more than once in Lysimachia, including in the Hawaiian endemic subgenus Lysimachiopsis while species of the South African subgenus Palladia are bird-pollinated, and so it goes; Renner and Schaefer (2010: summary) date the crown clade of Lysimachia to (41-)31(-8) Ma and the age of the stem clade to (52-)41(-28) Ma; Michez et al. (2007) described a fossil bee Palaeomacropis eocenicus, with hairs on its legs very similar to the oil-collecting hairs of Macropis itself, from France in amber from the early Eocene some 53 Ma (the somewhat younger fossil macropid Eomacropis glaesaria is not likely to have been an oil collector: Michez et al. 2008). Anderberg et al. (2007) suggested that Lysimachia with buzz-pollinated flowers and those with nectar-producing hairs formed separate clades and were both derived from oil-producing ancestors, but the pattern of gains and losses of oil flowers is complex (Schäffler et al. 2012; Renner & Schaefer 2010); there are also some selfers (e.g. Vogel 1986).
Otegui and Cocucci (1999) show that there is wind pollination in the dioecious Myrsine laetevirens.
The stigma of Cyclamen is wet, and is just inside the punctate tip of the hollow style (Reinhardt et al. 2007).
The pedicels of Cyclamen twist spirally as the fruits mature, the capsules ending up close to the ground, and the seeds are dispersed by ants attracted by the goo on the surace (?all species); most species of Cyclamen have rather local distributions (Yesson et al. 2009). Myrmecochory is common in Primula (Lengyel et al. 2010).
Plant-Animal Interactions. Primulaceae are not often eaten by butterfly larvae, but the 110 species of the largely Old World (there are two species from the Caribbean) Lycaenidae-Riodinidae-Nemeobiinae are known only from Primulaceae, especially Maesa, but so far they have not been found on members of Theophrastoideae (Espeland et al. 2015); Ehrlich and Raven (1964) noted other records.
Plant-Bacterial/Fungal Associations. About 35 species of Ardisia, probably most of the subgenus Crispiardisia, have pustules along the edge of the leaf blade inhabited by Burkholderia; the association is quite recent, ca 5 Ma, and the symbionts in Primulaceae may be close to leaf-nodulating bacteria in Rubiaceae (Lemaire et al. 2011b, c). The association involves a single strain/species of Burkholderia that has a much reduced genome, and there was a single origin of the bacterium—plant association; bacteria live in the apoplast (Parniske 2018; Pinto-Carbó et al. 2018), and they are also found in the shoot apex, flowers, embryo, etc.. Overall, it seems that transmission is likely to be vertical (Ku & Hu 2014; Carlier et al. 2016); the association is obligate and there is general phylogenetic concordance between the two members of the symbiosis, but strict cospeciation is unlikely and there seems to heve been some host switching (C.-J. Yang & Hu 2022). It was unclear what role the bacteria play (I. M. Miller 1990), although Carlier et al. (2016) suggest that some aspect of the secondary metabolism may be involved - perhaps the bacteria protection against herbivory (Pinto-Carbó et al 2018). For more information, see Miller (1990) and C.-J. Yang and Hu (2018).
Genes & Genomes. Smith and Donoghue (2008) found that the rate of molecular evolution of the herbaceous taxa they examined was much greater than that of the woody taxa. For a genome duplication perhaps to be associated with the [Primuloideae + Myrsinoideae] clade, see Larson et al. (2019/2020); a genome duplication in Aegiceras, but not involving Primula, was dated to ca 35 Ma (X. Feng et al. 2020/2021).
Chemistry, Morphology, etc.. Podolak et al. (2013) discuss a distinctive triterpene saponin that is at least scattered in the [Primuloideae + Myrsinoideae] clade. Interestingly, Maesa has less xyloglucans than other Primulaceae (Kooiman 1960).
For wood anatomy (not Primuloideae!) see de Luna et al. (2018). Vessels in Maesoideae are in radial multiples as is quite common in woody Theophrastoideae and in woody Myrsinoideae. There are small often peltate/glandular hairs; these may be stalked (Primuloideae) or more or less immersed (Theophrastoideae), while there are both kinds in Myrsinoideae (Große 1908). There may be groups of druses in the abaxial epidermis; the fibres are septate; and the lateral bundles arise about half an internode below the leaf they supply. In Theophrastoideae the stomata are anomocytic and there are several petiole bundles forming an arc which seem to diverge very soon from the leaf trace after it departs from the central stele (Ståhl 2004); the subepidermal fibres of Jacquinia may lack lignification. The peltate foliar trichomes of the halophytic J. armillaris secrete salt (Kuster et al. 2019). Discocalyx has three traces in the petiole base, and some other taxa may be trilacunar (e.g. unnamed taxon from the Atlantic Forest); nodal anatomy needs study. Indeed, Wheat (1980 and references) noted a few (three) cortical bundles in the stem of Ardisia and Myrsine that supplied the petiole, but these were absent at the bases of the axillary shoots and from herbaceous species of the latter genus; the nodes were unilacunar (1:1). For farina in Primuloideae, see Bourdon et al. (2021); farinaceous fibrils in Dionysia (= Primula) tapetodes are produced by largely unsubstituted flavones exuding through 1μm pores on the heads of trichomes; the function of farina here is unclear.
Wheat (1980) described the distinctive sylleptic branching common in woody Myrsinoideae. The foliar vernation of Theophrasteae and Myrsinoideae is often described as being involute (?supervolute, c.f. Cullen 1978) or conduplicate. The involute leaves of Primuloideae can be sharply bent rather than incurved (for vernation, see Conti et al. 2000; Mast et al. 2001). Solereder (1908) reported that secretory tissues occurred in Androsace lactea, which, along with a number of herbaceous Myrsinoideae, had "closed (schizogenous) secretory cavities" containing reddish material. Ståhl (2004) suggested that there was a secretory system in Theophrastoideae, if not always conspicuous in Samolus; for reports of glandular dots on the calyx and corolla there, see Mabberley (1997). There are breakdown areas in the rays of woody Myrsinoideae, and these may be filled with dark contents (Lens et al. 2005a); for secretory structures in Myrsinoideae, see de Luna et al. (2014 and references). However, the presence of coloured glands may well not be a synapomorphy of Myrsinoideae (Hao et al. 2004); clarification of the phylogeny is in order, as is more anatomical work...
The floral primordia of Theophrasteae may initially be quite strongly monosymmetric, as in Deherainia (Sattler 1962), even if the flower at anthesis is polysymmetric. Coris is particularly distinctive florally. Ronse Decraene et al. (1995) noted that monosymmetry there is expressed early in development by the calyx, but monosymmetry of the corolla becomes evident only later; they suggested that the median sepal is abaxial, i.e. that the orientation of the flower is inverted or oblique, and this is indeed evident in photographs. There are common stamen/corolla primordia in turn born on a ring primordium in Primulaceae, but the position/relative development of these primordia varies. In taxa like Cyclamen the stamens are initiated as adaxial outgrowths of a common primordium, i.e. the petal primordia are initially larger than the stamen primordia, as also in Myrsine and Aegiceras (see especially Ma & Saunders 2003), whereas in other taxa it is the stamen primordia that are initially larger, as in Samolus (e.g. Sattler 1962). Even in the late bud stage of some species of Lysimachia the still as yet tiny corolla encloses only the bases of the massive anthers. Saunders (1936) suggested that some of the lobing of the corolla of Soldanella might be staminodial. However, this is a tricky character, since there are really two variables, the relative positions of these primordia and the relative speed of their development, and, as with evicted terminal inflorescences, initial topological relationships between parts can speedily become disrupted by post-initiation growth. In Primuloideae the corolla epidermal cells are isodiametric, while in Myrsinoideae the epidermal cells of the corolla are often elongated; the latter is probably a derived feature within Primulaceae. Trientalis has anisomerous flowers (Swenson et al. 2008c). It has been suggested that the staminodes of Samolus and Theoprasteae are developmentally rather different (Caris & Smets 2004), while in some taxa lacking staminodes such as Maesa there is a staminodial vascular supply (e.g. Subramanyam & Narayana 1976; see also Saunders 1934). Wanntorp et al. (2012) discuss nectaries and nectar secretion around here; both buzz pollination and oil flowers are quite common and complicate the issue.
Information on floral development in Maesa is taken from Caris et al. (2000); the ovules there are often separated by and partly sunken in placental tissue (see also Warming 1913; Utteridge & Saunders 2001). In Samolus, the ovules completely cover the placenta, but fingers of placental tissue can poke up between them (but not seen in the material examined by Caris & Smets 2004); Ma and Saunders (2003) suggest that in Theophrastoideae in general the ovules are not embedded in placental tissue (which would then be a synapomorphy for it).
The number of carpels is difficult to ascertain (see especially Sokoloff et al. 2017a), but five seems to be a common number (see also the teeth on capsules). Although their orientation is often unclear, they might be expected to be opposite to the sepals. However, several of the diagrams presented by Dickson (1936) suggest that the carpels are opposite the petals (see also van Tieghem 1869b), and although in Primula, at least, the carpels appear to be opposite the sepals, this is not always so (Subramanyam & Narayana 1976). The capsule valves of Samolus are opposite the calyx (Caris & Smets 2004).
For general information, see Richards (2003: species of Primula), Anderberg (2004: Primulaceae), Firat and Lidén (2021: Evotrochis and relatives), Lidén and Mehregan (2023: Dionysia), Anderberg et al. (2000) and especially Ståhl and Anderberg (2004), both Myrsinaceae, Ståhl (2004: Samolaceae), Carey and Fisher (1932: embryology), Ståhl and Anderberg (2004) and Lens et al. (2005a), all Aegiceras, J. Sun and Wang (2024: Pomatosace) and Caris and Smets (2004: Theophrastaceae). For chemosystematics, see Harborne (1968: old-style Primulaceae), Colombo et al. (2017: Primula), for wood anatomy, see Lens et al. (2005a) and de Luna et al. (2018), for leaf anatomy, see de Luna et al. (2017), for vegetative and some floral anatomy, see Decrock (1901), for floral anatomy, see Dickson (1936) and Douglas (1936), for floral morphology, see Sattler (1962), Ronse Decraene (1992), Ronse Decraene et al. (1995), Otegui and Cocucci (1999: Myrsine, data matrix of this and related taxa), Endress (2019) and especially Ma and Saunders (2003), for anther crystals, see Pohl (1941), for pollen, see Nowicke and Skvarla (1977, 1979: old-style Primulaceae), Mast et al. (2001), Y. Xu et al. (2015), da Silva-Fourny et al. (2018, 2020) and Britskit and Grigoryeva (2021: Androsace s.l.), for nectaries and nectar/oil, variously produced, see Vogel (1986, 1997) and Caris and Smets (2004), for the hollow style, see Guéguen (1901), for embryology, etc., Warming (1913), and especially of the herbaceous taxa, see Dahlgren (1916), Subramanyam and Narayana (1976: anther wall development varies within Primula) and Woodcock (1926), Corner (1976) and Sundberg (1982a, b), all Cyclamen, and for seed and endosperm, for the most part poorly correlated with major clades, see Oh et al. (2008: herbaceous taxa).
Phylogeny. The monophyly of Primulaceae s.l. is not in doubt (see Anderberg & Ståhl 1995; Anderberg et al. 1998; Källersjö et al. 2000 in particular), although support values for Samolus as sister to Theophrasteae are reduced when morphological and molecular data are combined. In the morphological analysis of Anderberg and Ståhl (1995) herbaceous taxa grouped together, and Theophrastaceae were sister to the rest, i.e., relationships were basically old-style. Larson et al. (2023) recently looked at relationships between 133 species from 49/55 of the genera in the family using Angiosperms353 probes. What they found can be summarized as follows: [Maesoideae [Theophrastoideae [Primuloideae [Stimpsonia [Coris [[Ardisiandra [Cyclamen + Lysimachia]] [[Aegiceras + Monoporus] [[Embelia [Myrsine + Cybianthus]] [Ardisia - Old World [[Elingamita + Tapeinosperma] [Ardisia - New World]]]]]]]]] - for [Elingamita + Tapeinosperma], c.f. Yan et al. (2019) under Myrsinoideae below. Note that a number of genera are not monophyletic, even with the limited sampling, also, Larson et al. (2023) compared six trees, both analytic methods and sampling (close to Ericales as a whole vs. focus on Primulaceae), and all the trees might not support the relationships mentioned here - see Larson et al. (2023) for more details. Yan et al. (2024: four subfamilies, 28 genera, 67 species, plastomes) carried out a fairly comprehensive study of the family using plastome data; there are some differences from nuclear studies, including the position of Stimpsonia - Myrsinoideae (see above) or Primuloideae (Yan et al.)?
Maesoideae: Sumanon et al. (2023) looked at the phylogeny here - 126 taxa, the Angiosperms353 probe set - and recovered well supported relationships of the basal clades, although many of the nodes elsewhere in the tree had less support.
Theophrastoideae: For the phylogeny of Samolus, see Wanntorp and Anderberg (2011). Phylogenetic relationships in Theophrasteae suggested by Källersjö and Ståhl (2003) imply that some generic realignments are needed.
Primuloideae: For ITS-based relationships within the subfamily, see Martins et al. (2003), however, support for many nodes was weak. Overall relationships are also shown by de Vos et al. (2014a: suppl. Fig. 1), and for Chinese species by Z.-D. Chen et al. (2016), the latter recovering a quite well supported triad [Androsace etc. [Omphalogramma etc. + Primula s.l.]] (see also Larson et al. 2023). For relationships within Primula, see Trift et al. (2002), Mast et al. (2004, 2006), Yan et al. (2010) and Y.-J. Liu et al. (2015); for Dionysia (= Primula), see Trift et al. (2004); and for Soldanella, where S. villosa, from the Pyrenees, is sister to the rest of the genus, see Slovák et al. (2023: c.f. plastid and nuclear trees). Wang et al. (2004), Schneeweiss et al. (2004b) and Boucher et al. (2011: ITS plus trnL-F genes, 72 spp.) discussed relationships within Androsace, the latter recovering two main clades, one Central Asian and the other European-American. Yan et al. (2024) found that Stimpsonia was sister to other Primuloideae in their chloroplast analysis.
Myrsinoideae: The old Myrsinaceae included only woody taxa with fleshy, drupaceous fruits, but the clade that included these taxa (minus Maesa) was also found to include Anagallis (= Lysimachia), Ardisiandra, Asterolinon (?= Lysimachia), Coris, Cyclamen, Glaux (= Lysimachia, although it lacks a corolla), Lysimachia itself, Pelletiera, Stimpsonia and Trientalis (Anderberg et al. 2000, 2001), all of which are herbaceous and with capsular fruits - and used to be included in Primulaceae s. str.. However, the limits of this extended clade were not so clear in Martins et al. (2003: ITS data alone). Anderberg et al. (2007) was particularly interested in the relationships of the herbaceous taxa; the monophyly of Myrsinoideae s.l. had moderate support (72% jacknife), and Cyclamen, the other herbaceous taxa, and the woody taxa then formed a tritomy. Hao et al. (2004) also provide a phylogeny of much of the group, although with a focus on Lysimachia (for relationships here, see also Manns & Anderberg 2005). Yesson et al. (2009) found that herbs occupied basal pectinations within Myrsinoideae s.l., and Cyclamen (the focus of their study) was well embedded in the subfamily. The subshrub Coris was sister to the whole of the rest of the subfamily when Stimpsonia (moving between Primuloideae and Myrsinoideae) did not occupy that position - both genera have capsules opening by apical valves. The clade [Coris + Stimpsonia] was sister to other Myrsinoideae in the phylogeny recovered by Rose et al. (2018). For Chinese taxa, see Z.-D. Chen et al. (2016).
Relatively less work has been carried out on relationships in the woody Myrsineae. Aegiceras is often embedded in these taxa (e.g. Anderberg et al. 2008). For the phylogeny of Badula, recognition of which may make Oncospermum paraphyletic, see Bone et al. (2012), Appelhans et al. (2020) and especially Strijk et al. (2014). In an ITS/ETS phylogeny Appelhans et al. (2020) focused on Pacific Myrsine (and in a RAD-seq analysis, on the species from the Hawaiian islands), while J. Wang and Xia (2013) provide a morphological phylogeny of Chinese Ardisia. Otegui and Cocucci (1999) in a morphological analysis placed Aegiceras sister to other woody Myrsineae, while Appelhans et al. (2020) and Larson et al. (2023) found that the small and very heterogeneous clade [Aegiceras + Monoporus] was sister to the other woody Myrsineae. In a plastome analysis, Yan et al. (2019) recovered the relationships [[Aegiceras + Myrsine] [Embelia [[Elingameta + Parathesis] [Tapeinosperma + Ardisia]]]]. Julius et al. (2021) began a major assault on the whole group with an initial focus on Ardisia; 90 spp. were examined, also 15 species in six related genera from both Ardisieae and Myrsineae s. str. were included. Ardisia was very much paraphyletic, and its subgenera tended to be polyphyletic, while the American Ardisia opegrapha and Labisia were successively sister to the rest of the whole complex. Within Ardisia those species of Badula, Labisia, Afrardisia and Discocalyx that were studied formed separate clades, even if their positions might be ambiguous (Julius et al. 2021). In a more extensive analysis using Angiosperms353 data Larson et al. (2023) found that some 19 genera were part of a paraphyletic Ardisia (c.f. the topology there with that in Yan et al. 2019); for problems with the circumscription of Ardisia, see also C.-J. Yang and Hu (2022). Flickinger et al. (2026: 50 spp., ITS and trnL-trnF) looked at relationships within the Caribbean Myrsinoideae; Ardisia and Myrsine were not monophyletic.
Classification. Primulaceae were often placed in a separate Primulales in the past. The only question, particularly in light of the break-up of Primulaceae s. str., the removal of Maesa from Myrsinaceae, the placement/addition of Samolus as sister to the old Theophrastaceae, the herbaceous ex-Primulaceae that are sister to the old-style, woody Myrsinaceae rather than being in a clade with the other Primulaceae, and the numerous features shared by the group as a whole, was whether it was worth recognising separate families at all - ordinal status was certainly inappropriate. A broad circumscription of Primulaceae was proposed in A.P.G. III (2009); Primulaceae s.l. are well characterized, and available subfamilial and tribal names fit well with the phylogeny - but see Myrsinoideae below.
Primula could include Cortusa, Dionysia (see Lidén 2007 for a revision; Lidén & Mehregan 2023), and Dodecatheon, however, it is unclear where generic limits here will be drawn. If Dionysia is recognised, as provisionally by Lidén and Mehregan (2023), then it would entail the recognition of seven genera in place of Primula s.l.; perhaps there should be three tribes in Primuloideae (see relationships above). The limits of Androsace will have to be extended to include Douglasia and Vitaliana (Schneeweiss et al. 2004b; Boucher et al. 2011; Britskit & Grigoryeva 2021). Generic limits in the woody Myrsinoideae/Myrsineae in particular are in a mess; it may be that the large genus Ardisia will be divided (its old infrageneric classification has some problems, but it could be worse), although a great deal of work is needed before this is done (Larson et al. 2023) - note that the type of Ardisia, A. tinifolia, is from Jamaica. Flickinger et al. (2026) adjusted the limits of some Caribbean Myrsinoideae; their work represents a reasonable nomenclatural compromise given how much of the bigger picture of relationships here remains unclear.
There are no problems with the circumscription of Maesa, and Sumanon et al. (2023) divide it into five subgenera. The limits of genera like the herbaceous Lysimachia have seemed unclear (Anderberg et al. 2007), but Manns and Anderberg (2009) provide combinations and an infrageneric classification for an expanded Lysimachia.
Previous relationships. Plumbaginaceae (see Caryophyllales here) were often associated with Primulaceae, both having features in common like apparently similar placentation and stamens opposite the petals (see Cronquist 1981 for discussion; Nowicke & Skvarla 1977 for what pollen had to say).
[Mitrastemonaceae, Theaceae, [Symplocaceae [Styracaceae + Diapensiaceae]], [[Sarraceniaceae [Roridulaceae + Actinidiaceae]] [Clethraceae [Cyrillaceae + Ericaceae]]]] (if the clade exists): testa with outer periclinal wall unthickened.
Age. The age of this node is (98.4-)78.4(-55.8) Ma (Naumann et al. 2013: [Clethraceae + Ericaceae] sister) or ca 104 Ma (Rose et al. 2018: sister to Lecythidaceae, in turn sister to everything excpt balsaminoids...).
MITRASTEMONACEAE Makino, nom. cons. - Mitrastemon Makino - Back to Ericales
Holoparasitic, root parasites, plant endophytic; ?anatomy; leaf waxes hummocky; leaves opposite, scale-like; flowers single, terminal; P uniseriate, 4; anthers extrorse, completely connate and surrounding G except for small apical pore, polythecate; pollen 2-porate [?colpate], ektexine reduced to tuberculae; G 8-20, placentation intrusive parietal, style stout, stigma hemispherical; ovules many/carpel, unitegmic, integument ca 2 cells across, funicular obturator +; fruit berry-like, circumscissile; seeds dust-like [in size], funicle sticky; exotestal cells with massive U thickenings; endosperm 1-layered, 10-15-celled, embryo undifferentiated, 4-celled; n = 20.
1/2: [list]. N.E. India and Japan (Honshu to Papua New Guinea, scattered, S.E. Mexico to Colombia. Map: from van Steenis and van Balgooy (1966), Meijer and Veldkamp (1993) and Amoroso et al. (2018). [Photo - Habit © S. Hsiao.]
Evolution: Divergence & Distribution. Estimates to the stem ages of Fagaceae and Mitrastemonaceae may be similar, suggesting perhaps that the two evolved together, but other scenarios are possible (Teixeira-Costa & Suetsugu 2022).
Ecology & Physiology. The hosts of Mitrastemon appear to be mainly Fagaceae where the parasite lives in the roots; the xylem anatomy of the host is quite extensively affected (Teixeira-Costa & Suetsugu 2022). L. Cai (2023) discussed the evolution of parasitism in flowering plants.
Pollination Biology & Seed Dispersal. Information is available only for Mitrastemon yamamotoi. That species seems to be pollinated by a variety of insects including social wasps (most effective in northern locality), crickets and cockroaches, perhaps also birds, much nectar being produced by and collecting in the axils of the uppermost pair of leaves. Seeds are dispersed by birds (Suetsugu 2019; Teixeira-Costa & Suetsugu 2022).
Genes and Genomes. The rate of genome evolution in Mitrastemon is rather slower than that of other parasitic taxa ().
A mitochondrial gene has moved from host to the parasite (Barkman et al. 2007: atp1) and from the parasite to its host, Quercus (Systma et al. 2009). The mitochondrial genes cox1 and matR showed considerable divergence, but not the atp1 gene (Barkman et al. 2007).
Chemistry, Morphology, etc.. Watanabe (1936: V) talks a lot about a "Mitrastemon-Pilz" - c.f. the ectomycorrhizae of Ericaceae?
The pollen may have three or four pores - see Watanabe (1936: III). Cronquist (1981) and Meijer and Veldkamp (1993) describe the fruit as being a berry or berry-like and opening via a transverse slit, i.e., it is also some sort of circumscissile capsule, while Meijer and Veldkamp (1993) described the ovule as being unitegmic and the seed coat as as being formed from the inner integument (the latter following Watanabe 1937: VII).
For general information (including a more extensive list of hosts) and references, see Meijer and Veldkamp (1993), the Parasitic Plants website (Nickrent 1998 onwards)\, Heide-Jørgensen (2008) and Teixeira-Costa and Suetsugu (2022).
Previous Relationships. Mitrastemonaceae were included in the old Rafflesiales (e.g. Cronquist 1981). Cocucci and Cocucci (1996) suggested that Mitrastemonaceae had relationships with Annonaceae.
[Theaceae, [Symplocaceae [Styracaceae + Diapensiaceae]], [[Sarraceniaceae [Actinidiaceae + Roridulaceae]] [Clethraceae [Cyrillaceae + Ericaceae]]]]: cork?; vessel elements with scalariform perforation plates; lamina margin serrate; Ad-β genome duplication.
Age. This node (as [Theac + Styr] or equivalent) has been dated to 103 Ma (K. Bremer et al. 2004a), (101.5-)91.5(-80) Ma (X.-Q. Yu et al. 2017), (97.3-)89.8(-68.1) Ma (M. Rao et al. 2018) or (99.8-)92.1(-84.4) Ma (Yan et al. 2021), if including Ericaceae, etc., a date closer to 106 Ma is more likely; Magallón et al. (2015) estimated the age of this node to be around 94.9 Ma, (103-)96(-88) Ma is the estimate offered by Wikström et al. (2015), ca 101.9 Ma by Rose et al. (2018) and (102.5-)99.7(-92.0) Ma by Q. Zhang et al. (2022: [Theac + Symp]); there are also several estimates ([Theac + Actin]) of around 65-61 Ma (Xia et al. 2020; Zhang et al. 2022 and references). See also Genes & Genomes below.
Evolution: Ecology & Physiology. Plants in this clade have relatively low leaf nitrogen, and other major shifts in this part of the tree (see also Ericaceae, and [[Sarraceniaceae [Actinidiaceae + Roridulaceae]] [Clethraceae [Cyrillaceae + Ericaceae]]) suggest slow carbon and nutrient cycling (Cornwell et al. 2014).
Genes & Genomes. A genome duplication, Ad-β, that is somewhere around here has been dated to ca 75.9 or 101.4-72.9 Ma (Shi et al. 2010 and S. Huang et al. 2013 respectively); see also Soza et al. (2019) who suggest that the duplication occured around 85.6 Ma and Diaz-Garcia et al. (2021) who date a duplication - they also call it the Ac-β and Vm-α duplication, but I think they are talking about the same thing - to ca 61 Ma (they also give other ages). See also Polemoniaceae, etc. above.
THEACEAE Ker Gawler, nom. cons. - Back to Ericales
Trees or shrubs; plants Al-accumulators; myricetin, ellagic acid +; cork cambium pericyclic; (pits vestured); intervessel pitting opposite-scalariform; pericyclic fibres +/0; petiole bundle arcuate; sclereids +, mucilage cells +; stomata usu. anomocytic (anisocytic or cyclocytic); hairs unicellular; leaves spiral, lamina involute or supervolute (conduplicate), margins toothed; flowers single, axillary, bracteoles +; C ± free; A usu. 40<, development centrifugal, ± basally connate, anthers versatile, articulated, connective usu. not prolonged, filaments variable in length; anther epidermis ephemeral/partly persistent, middle layer 3-4 cells across; pollen tricolporoidate; nectary?; G [(3-)5(-10)], opposite C, (styles +, separate), stigma wet; ovules 2-few/carpel, (basal), bitegmic, micropyle endostomal, outer integument 4-10 cells across, inner integument 4-11 cells across, hypostase +; antipodals in mature embryo sac 0; fruit a loculicidal capsule, K persistent or not; seeds few, often >4 mm long, flattened; testa massive, exotesta lignified or not, mesotesta lignified (fibrous; with sclereids), endotesta lignified or not; endosperm nuclear/coenocytic, cotyledons longer than radicle, accumbent; x = 6, nuclear genome [1 C] (1.053-)3.098(-9.113) pg.
9/195(-460!): [list, to tribes] - three tribes below. Mostly warm temperate to subtropical India-South East Asia and Malesia, some in the Americas, Africa none. [Photo - Collection.]
Age. Crown-group Theaceae are some 49 Ma (M.-M. Li et al. 2013), (74.7-)57.3(-39.6) Ma (X.-Q. Yu et al. 2017, q.v. for several other dates), ca 63.1 Ma (Rose et al. 2018), (71.8-)59.6(-48.8) Ma (M. Rao et al. 2018), (73.4-)66.4(-60.0) Ma (Yan et al. 2021), (73.3-)64.9(-56.4) Ma (Q. Zhang et al. 2022) or ca 66.5 Ma (Zan et al. 2023).
Pentapetalum trifasciculandricus, a fossil ca 91 Ma from New Jersey, may belong to Theaceae or be in the Pentaphylacaceae area (Martínez-Millán et al. 2009: c.f. analyses). For other information on the fossil record of Theaceae, see Martínez-Millán (2010).
1. Stewartieae Choisy - Stewartia L.
(Plant deciduous); pith heterogeneous; C connate; androecium fasciculate; pseudopollen type?; embryology?; capsule lacking columella; seeds narrowly winged or not; exotesta cells ± polygonal, (surface with ± detailed sculpturing); integument vascularization?; endosperm +; x = 17, n = 15, 17, 18.
1/9. East Asia, E. North America. Map: from Hong (1993).
Age. Crown-group Stewartieae are estimated to be around 18.7 Ma (M. M. Li et al. 2013), (24.7-)14.8(-8.3) Ma (X.-Q. Yu et al. 2017), (28.7-)22.7(-15.1) Ma (Yan et al. 2021) or (21.7-)18.5(-15.6) Ma (Q. Zhang et al. 2022).
[Gordonieae + Theeae]: stomata anisocytic/cyclocytic; androecium whorled; capsule with persistent columella; endosperm slight to 0.
Age. This node has been dated to around 68 Ma (K. Bremer et al. 2004a), ca 60 Ma (Rose et al. 2018), (66.9-)49.8(-33.8) Ma (X.-Q. Yu et al. 2017), (58.3-)50.2(-43.4) Ma (M. Rao et al. 2018), (26.4-)25.6(-20.8) Ma (Yan et al. 2021), (62.8-)55.7(-48.1) Ma (Q. Zhang et al. 2022) or ca 45.1 Ma (Zan et al. 2023).
2. Gordonieae de Candolle —— Synonymy: Gordoniaceae Sprengel
(Plant deciduous - Franklinia); stem cork cambium superficial; lamina entire; androecium in 3-5 whorls [?is this a character], connective with stomata; pseudopollen with pores; (ovule campylotropous), inner integument vascularized; dehiscence also septicidal; seeds apically/marginally/not winged; testa proliferating, surface with irregular protrusions, tegmen also proliferating; (embryo curved - Schima); x = 18, n = (15), 18.
3/40: Gordonia (22), Schima (17). Southeast Asia, West Malesia, S.E. United States. Map: from Camp (1947) and Bloembergen (1952).
Age. Crown-group Gordonieae are about 11.1 Ma (M.-M. Li et al. 2013), (32.2-)26.2(-23.3) Ma (X.-Q. Yu et al. 2017) or (26.6-)25.1(-24.1) Ma (Q. Zhang et al. 2022).
Grote and Dilcher (1992, also references) described Gordonia lamkinense from rocks of the Claiborne Formation of Kentucky ca 40.4 Ma, as well as other Theaceae from there (see also Martínez-Millán 2010).
3. Theeae Szyszylowicz —— Synonymy: Camelliaceae de Candolle
(CAM photosynthesis - fungal infection); pith heterogeneous; pedicels multibracteolate; K and C intergrading; A in 2 whorls, obdiplostemonous; nectary at bottom of / whorl-shaped surrounding base of filaments; pseudopollen ribbed (0); (G opposite K - Camellia); outer integument becoming vascularized, (endothelium +), (hypostase 0); (embryo sac bisporic [chalazal dyad], eight-celled [Allium-type] - Camellia); seeds (apically winged); exotesta cells elongated to polygonal, surface smooth; (cotyledons much folded); x = n = 15.
5/230-420: Camellia (120-280), Laplacea, Polyspora (32), Pyrenaria (42-60). Southeast Asia, Malesia, tropical America. Map: from Camp (1947, approximate), see also Yan et al. (2021: Fig. 1). [Photo - Flower, Fruit.]
Age. Crown-group Theeae have been dated to (39.2-)27.1, 27(-18.4) (W. Zhang et al. 2014), (30-)19.4(-11.6) Ma (X.-Q. Yu et al. 2017) or (25.2-)22.1(-19.2) Ma (Q. Zhang et al. 2022).
Fossil leaves identified as Theeae are known from the Upper Eocene of Japan, woods (?= Camellia) from Lower Oligocene deposits ca 30 Ma from both Bulgaria and Washington, W. U.S.A. (L.-L. Huang et al. 2016), and Camellia-type pollen perhaps 50 Ma is known from Europe (Hofmann 2018). Note, however, that crown-group Camellia is estimated to be a mere (18.7-)15.9, 12.3(-7.3) Ma by W. Zhang et al. (2014).
Evolution: Divergence & Distribution. Pollen similar to that of the Southeast Asian Polyspora (Theeae) has been found in Austria in deposits only some 17-11 Ma (Hofmann & Lichtenwagner 2019).
For some other dates in Theaceae, see X.-Q. Yu et al. (2017), Zan et al. (2023), etc..
Y. Yan et al. (2021) suggested that if fossils were taken into account, Theaceae in the earlier Cenozoic could be shown to belong to the circumpolar boreotropical forest biome, trans-Pacific disjunctions resulting from movements over the Asian-American land bridge, and with subsequent extinctions at higher latitudes as the climate deteriorated (see also Q. Zhang et al. 2022).
Zan et al. (2023) thought that diversification in Camellia - crown age ca 25.2 Ma - was associated with the evolution of evergreen broad-leaved forests ca 23-19 Ma, although D. W. Zhao et al. (2022) suggest a crown-group age of ca 39.5 Ma for the genus. Q. Zhang et al. (2023) noted i.a. extensive gene tree discordance (noted by others, too) which they attributed largely to rapid evolution and ILS, perhaps hybridization. Zan et al. (2023) thought that there had been extensive hybridization in Camellia (Zan et al. 2023), although for Q. Zhang et al. (2022) there were just two reticulation events here and one between the common ancestor of Gordonia and Schima. H.-Y. Lin et al. (2019) suggested that there had been ancient introgression after diversification in Stewartia.
Ecology & Physiology. Theaceae, along with Magnoliaceae, Lauraceae and Fagaceae (Tang 2015), are a notably prominent component of the subtropical evergreen broad-leaved forests (EBLFs) of East Asia, over 55% (= 148 spp.) of the family growing there and another 33% being known from Tropical Asia (X.-Q. Yu et al. 2017). Species diversity of Theeae in particular was associated with low pH (and high aluminium) conditions (M. Rao et al. 2018).
Pollination Biology. Various kinds of Diptera including calliphorid and chironimid flies pollinate flowers of species of Eurya in Japan; the nectar tends to be concentrated, as in other fly-pollinated flowers (Tatsuno & Osawa 2024). Nectar of Camellia perpetua changes composition with the seasons, and is richer in sucrose in the winter - bird-pollination especially then (H.-d. Jiang et al. 2025). The function of the pseudopollen is unknown, but it does not appear to be nutritious and it may be deceit pollen (Tsou 1997; Iqbal & Wijesekara 2002); not all taxa, e.g. of Camellia, have it (Q. Zhang et al. 2017).
Seeds of Franklinia take about a year to mature, for although fertilization takes place soon after pollination in the autumn, the young fruits overwinter in a state where the endosperm is cellularized but the zygote is still undivided (Schoonderwoerd & Friedman 2015, esp. 2016).
Plant-Animal Interactions. For the remarkable interactions between Camellia japonica and its seed predator, the camellia weevil (Curculio camelliae), see Toju et al. (2011). The capsule of the plant may be the size of an apple, the pericarp being over 2 cm thick, but the snout of the weevil, used for boring into the fruit to make a hole for subsequent egg deposition in the seeds, may be over 2 cm long.
Plant-Bacterial/Fungal Associations. Yuan et al. (2012) noted that leaves of Camellia oleifera infected by Exobasidium vexans were fleshy - and rather remarkably carried out CAM photosynthesis, the stomata opening during the night, etc.. For more on Exobasidium, see Ericaceae.
Genes & Genomes. There may be a genome duplication, the Cmα duplication, in a clade including Gordonieae and Theeae, at least (Larson et al. 2019/2020), while Q. Wu et al. (2022) suggested that there had been a recent whole genome duplication in Camellia.
For the base chromosome numbers of the three tribes, see Hembree et al. (2019), also Horiuchi and Oginuma (2001: Stewartieae). Hembree et al. (2019) also determined monoploid genome sizes: The mean 1Cx in Gordonieae was 0.84 pg, while in Stewartieae and Theeae it was 2.50 and 3.00 pg respectively.
Chemistry, Morphology, etc.. The cotyledons of (?all) Theaceae have three or more traces from a single gap. The stomata are often described as being "gordoniaceous", i.e. cyclocytic to anisocytic (see e.g. Lu et al. 2008).
The androecium of Camellia japonicahas a basically obdiplostemonous construction (Sugiyama 1997). Although the carpels seem to be opposite the sepals in Camellia, this may be connected with the arrangement of the perianth, rather than that of the gynoecium per se; the basic orientation of the gynoecium with respect to the floral axis is the same as that of Gordonia, where the carpels are clearly opposite the petals (Eichler 1878).
For general information about Theaceae s.l., see Keng (1962) and Stevens et al. (2004b), also Beauvisage (1920) and Liang and Baas (1991), both anatomy, Zhang et al. (2009: sclereids in Camellia), Jiang et al. (2010: lenticels on Camellia leaves); also Leins and Erbar (1991: floral morphology), Wei (1997: pollen), Fagerlind (1939c), Yang and Min (1995a, b), Tsou (1997, 1998) and Q. Zhang et al. (2017), all embryology, Gunathilake et al. (2015: testa surface) and Wang et al. (2006: Apterosperma, chromosomes and morphology).
Phylogeny. Y. H. Wang et al. (2006) looked at a number of morphological characters in the context of a cladistic analysis that focused on Theeae-Apterosperma. An analysis of two chloroplast genes by Prince and Parks (2001) suggests that there are three major clades (see above) and that Polyspora and Laplacea should be separated from Gordonia (see also Airy-Shaw 1936; Yang et al. 2004: genes from all three genomes, 2006: mitochondrial gene only, study of Theaceae s.l., including Pentaphylacaceae). However, relationships between these three clades remain unclear. The clade [Gordonieae + Theeae] has some support (see also Prince 1999; Yang et al. 2004; Rose et al. 2018) and makes morphological sense, although an analysis of matK data alone suggested that Theeae were sister to the other two tribes, but there was a polytomy in the combined analysis (including rbcL data: Prince & Parks 2001); see also Su et al. (2011: Apterosperma sister to [Tutcheria + Camellia]), M.-M. Li et al. (2013: several chloroplast genes, 11/20 species sampled were Stewartia) and Z.-D. Chen et al. (2016: Stewartia sister to the rest, but support weak). W. Zhang et al. (2014) found that Polyspora was sister to other Theeae using the LEAFY gene (see also Rose et al. 2018), Camellia was paraphyletic, but there was little resolution along the spine of the tribe in a matkK + rbcL analysis; Apterosperma may be of ancient hybrid origin. Details of relationships differed again in the plastome and n-ribosome analysis of X.-Q. Yu et al. (2017); the relationships [Stewartieae [Gordonieae + Theeae]] were usually obtained, but in single-gene analyses there was weak support for Theeae and for Gordonieae being sister to the rest of the family. M. Rao et al. (2018: 10 chloroplast loci and nrITS) found the relationships [Frankjlinia [Gordonia + Schima]] and [[Pyrenaria + Laplacea] [[Apterosperma + Polyspora] Camellia]]] in Gordonieae and Theeae respectively. Y. Yan et al. (2021: plastome coding regions + nuclear ribosomal DNA) recovered the relationships followed above: [Stewartia [[Gordonia [Franklinia + Schima]] [Pyrenaria [[Apterosperma + Laplacea] [Polyspora + Camellia]]]]], but again, the support for the relative positions of the first two tribes was weak. In this study the old Gordonia was highly polyphyletic, appearing in four separate clades, and Camellia was almost monophyletic, but C. gracilipes tended to migrate (Yan et al. 2021: that species was not included by M. Rao et al. 2018 or Zan et al. 2023 below).
For relationships within Camellia, see Vijayan et al. (2009), who noted that current sectional limits needed overhauling, and M. Rao et al. (2018), who found that [C. lanceolata + C. hekouensis] were sister to the rest of the genus. There has been much recent work on the genus, but many problems remain. Pang et al. (2022), using four plastid markers to look at relationships among 161 species recovered 13 clades, but then-current sections did not fit very well into those clades. Zan et al. (2023: 87 spp., 1481 low-copy nuclear genes) found a number of well supported clades in Camellia, but relationships between these clades were unclear, differed when subsets of the genes were examined, and differed from the relationships obtained using plastome data. Other recent studies include those by D. W. Zhao et al. (2022: 99 taxa, 3 nuclear markers), Q. Wu et al. (2022: transcriptome data, 116 plants) and Q. Zhang et al. (2023: 55 spp., 3 data sets of up to 1,617 orthologous low-copy nuclear genes). The latter group found little support for many relationships in the genus and plastome data again suggested different relationships; C. hekouensis was sister to the rest of the genus and was separated from it by a long branch (it even moved outside the genus in some analyses; C. lanceolata was not included (Zhang et al. 2023). For the limits of Pyrenaria, a genus which is best broadly drawn, see R. Li et al. (2011). Within Stewartia, S. malacodendron is sister to the rest of the genus (X.-Q. Yu et al. 2017), however, relationships here may be obscured by ancient introgression (H.-Y. Lin et al. 2019). Laplacea has turned out to be wildly polyphyletic, L. grandis being sister to Gordonia lasiantha and L. fruticosa being embedded in Theeae (Yu et al. 2017).
Classification. Stewartia is to include Hartia (e.g. Prince 2002; M.-M. Li et al. 2013). Generic limits in other Theaceae are difficult, but for useful notes on the genera, see Prince (2007). Franklinia hybridizes with Schima, and perhaps Gordonia and even Camellia, i.e. taxa scattered throughout Gordonieae and Theeae (Ranney et al. 2003; Hembree et al. 2019); generic limits are being reworked.
Both species limits and infrageneric classifications in Camellia are unclear (see Vijayan et al. 2009; Hembree et al. 2019). Q. Zhang et al. (2023) and Zan et al. (2023) noted extensive problems with sectional limits, previous sections very largely lacking support. Species limits in Schima were also found to be difficult (Bloembergen 1952).
Previous Relationships. Theaceae s.l. have in the past been associated with Asteropeiaceae (e.g. Takhtajan 1997), now in Caryophyllales, and Bonnetiaceae (see Malpighiales) (e.g. Cronquist 1981). Pentaphylacaceae, Sladeniaceae and Pellicieraceae, also erstwhile Theaceae, are in separate families in Ericales, the last quite apart from the other two.
[[Symplocaceae [Styracaceae + Diapensiaceae]] [[Sarraceniaceae [Actinidiaceae + Roridulaceae]] [Clethraceae [Cyrillaceae + Ericaceae]]]] - if this clade exists: ?
Age. The crown-group age of this clade is around 101.6 Ma (Rose et al. 2018), 93.2 Ma (Magallón et al. 2015) or ca 83 Ma (Tank et al. 2015: Table S1).
[Symplocaceae [Styracaceae + Diapensiaceae]]: shrubs to trees; lignans +; inflorescence racemose; A (obdiplostemonous), often ± forming a tube; style hollow; endothelium 0; endosperm copious, haustorium 0.
Age. The age of this node is some (68-)63, 61(-56) Ma (Wikström et al. 2001), about 100 Ma (K. Bremer et al. 2004a), 99.3 Ma (Rose et al. 2018), 81.2 Ma (Magallón et al. 2015), (94-)92(-89) Ma (Fritsch et al. 2015) or (95.5-)84(-71.5) Ma (X.-Q. Yu et al. 2017).
Evolution: Divergence & Distribution. In the context of the floral morphospace of Ericales, Chartier et al. (2017) thought that this clade was morphologically rather homogeneous.
Characters like presence of ellagic acid and a vascularized integument could be optimized to this node, but they would later have to be lost. The stamens often have rather broad filaments, and they tend to form a tube around the gynoecium.
SYMPLOCACEAE Desfontaines, nom. cons. - Back to Ericales
Plants Al-accumulators, O-methyl flavonols, route II decarboxylated iridoids, ellagic acid +, myricetin 0; true tracheids +; crystal sand +; stomata usu. paracytic, also very large water stomata +; (leaves two-ranked), lamina ± supervolute; (plant dioecious); (inflorescence branched), pedicels articulated/(not - Cordyloblaste); K basally connate; A (= and opposite K)-many, in bundles, (connate), adnate to C; anthers globose; pollen angular, spinuliferous or not, (3-pororate); G [2-5], (half) inferior, median member abaxial, stigma ± capitate, wet or dry; nectary on ovary; ovules 2-4/carpel, pendulous, epitropous, endothelium +; fruit drupaceous, stone with as many pores as fertile carpels, K persistent; seed usu. 1; testa vascularized, (exotestal cells with inner walls thin); embryo large, (curved); n = 11 (12), x = 11; mitochondrial coxII.i3 intron 0.
2/320(-260): [list], Symplocos (318). Tropical to subtropical, inc. New Caledonia, not Africa. Map: see Nooteboom (1975), c.f. Fritsch et al. (2015) - nothing in Amazonia). [Photo - Symplocos chinensis Flowers.]
Age. Crown-group Symplocaceae are estimated to be (57-)52(-48) (Fritsch et al. 2015) or ca 47.8 Ma (Rose et al. 2018).
Symplocos fruits are common in the fossil record and have been dated up to ca 48.6 Ma ( Martínez-Millán 2010 and references).
Evolution: Divergence & Distribution. Symplocos is locally very abundant as both pollen and fruits in the Caenozoic fossil record of Europe; it is also known from western North America, the southern USA and East Asia (Krutzsch 1989; Fritsch et al. 2015), as well as New Zealand (Lee et al. 2001). The family may be Eurasian, not strictly South East Asian, in origin (Xing et al. 2016; Q. Zhang et al. 2021), and diversification of [section Palura + The Rest], i.e., most of the family, is dated to only (41-)38(-35) Ma (Fritsch et al. 2015, q.v. for much information on biogeography).
Pollination Biology. There are subapical lobes on the style just below and alternating with commissural "stigmatic" lobes in the ca 145 species of the New World Symplocos subg. Symplocos sect. Symplocastrum; the papillae on these lower lobes are rich in lipids that may help the pollen stick to the pollinators (Kriebel et al. 2007). Pollen germinates on these subapical lobes which are thus (and from their position) true stigmatic lobes (Kelly & Nicholson 2009).
Chemistry, Morphology, etc.. Although the placentation is described as being fully axile, in material seen it is parietal at the apex. The androecium is basically obdiplostemonous (Caris et al. 2002); Basak et al. (2023) discuss potae/pororate pollen here.
For general information, see Nooteboom (2004), for pollen, see Nagamasu (1989), and for testa anatomy, see Corner (1976) and Huber (1991).
Phylogeny. For a phylogeny of Symplocos s.l., see Y. Wang et al. (2004) and Fritsch et al. (2006, 2008, 2015); section Cordyloblaste, with two species, appears to be sister to the rest of the genus and section Palura sister to the remainder (one node up in Wang et al. 2004). S.-C. Kim et al. (2021) looked at the plastomes of seven species - nothing remarkable.
Classification. The infrageneric taxonomy of Symplocos needs reworking, and two genera should perhaps be recognised (Fritsch et al. 2008).
[Styracaceae + Diapensiaceae]: cork cambium pericyclic; glandular hairs 0; leaves spiral, (margins entire); anthers basifixed; nectary 0; style continuous with ovary; ovules many/carpel; fruit a loculicidal capsule.
Age. The age of this node is estimated at (60-)55, 45(-40)Ma by Wikström et al. (2001), (78-)54, 51(-39)Ma by Bell et al. (2010), about 52.7 Ma by Magallón et al. (2015), ca 73 Ma by Tank et al. (2015: Table S2), and as much as 93.2 Ma by Rose et al. (2018).
If the fossil Actinocalyx is stem-group Diapensiaceae (see below), then the age of this clade is at least 83.5 Ma.
Evolution: Divergence & Distribution. Scott (2004) and Fritsch (2004) suggest that there are embryological features in common between the two families; I do not know if any of them are really synapomorphies.
Phylogeny. There is also fairly good support for this clade in B. Bremer et al. (2002).
STYRACACEAE Candolle & Sprengel, nom. cons. - Back to Ericales —— Synonymy: Alniphyllaceae Hayata, Halesiaceae D. Don
Trees or shrubs, (deciduous); ellagic acid, myricetin 0, iridoids?; (vessel elements with simple perforation plates); wood siliceous; resin canals often +; petiole bundle arcuate or D shaped (medullary and/or wing bundles +; complex - Parastyrax); indumentum stellate or scaly; (buds perulate); lamina vernation conduplicate-plicate or supervolute; bracteoles 0, pedicels articulated/not [Styrax]; flowers (4-)5(-7)-merous; K ± completely connate, open, (1 trace), C valvate or not; A 2 (3) x C/= and alternate with C, adnate to C, often basally connate, anthers basifixed, usu. "long", connective well developed, running the length of the anther, (filaments as broad as anther - Styrax sect. Pamphilia); pollen surface ± smooth; G [2-5], ± inferior, alternate with K, median member ?abaxial, often with hairs inside, (long style branches +), stigma punctate or lobulate, dry; ovule (1/carpel, basal - Pamphilia), in two ranks, often apotropous, integument 8-15 cells across, (apically bitegmic, outer integument t 6-9 cells across, inner integument 3-4 cells across, micropyle endostomal - Styrax), micropyle quite long, placental obturator +; fruit drupaceous/nut-like/samara/loculicidal capsule; testa multiplicative, exotesta much thickened, thickening O- or U-shaped, (testa vascularized, outer mesotesta sclerotic - Styrax), ± crushed or not; n = 8, x = 6 (?7, ?8), nuclear genome [1C] (0.11-)1.251(14.276) pg.
11/160: [list], Styrax (120). Japan to Malesia, esp. Southeast Asia, Mediterranean and thereabouts, North America, Styrax alone in Central and South America. Map: from van Steenis (1949b), Sales and Hedge (1996) and Fritsch (1999). Photo Flower.
Age. Crown-group Styracaceae may be ca 56 Ma (Rose et al. 2018) and pollen of Styrax a little younger than this is reported from Europe (Hofmann 2018).
Evolution: Divergence & Distribution. For the early Caenozoic fossil history of Styracaceae that are now East Asian endemics, see Manchester et al. (2009). Pollen like that of Rehderodendron, now growing in mainland Southeast Asia, has been found in deposits only around 17-11 Ma from Austria (Hofmann & Lichtenwagner 2019).
Fritsch et al. (2001) suggest possible additional synapomorphies for Styracaceae.
Plant-Animal Interactions. Van Steenis (1949b) illustrates the remarkable galls found on Malesian species of Styrax. There is a rather close association between the aphids involved (Cerataphidinae) that cause some of these galls and individual species of Styrax; the morphology of the galls is ultimately determined by the aphids (Stern 1995; Stern & Foster 1996; J. Chen et al. 2014 for a phylogeny of the aphids). Cecidomyids also produce galls on Styrax.
Plant-Bacterial/Fungal Associations. Glands containing bacteria have been reported on the leaves of the Brazilian Styrax camporum (Machado et al. 2014; see also C.-J. Yang & Hu 2018).
Genes & Genomes. A 20kb inversion in the large single copy region of the plastome characterises the [Bruinsma + Alniphyllum] clade and the accD gene has become a pseudogene, which is quite common in this part of Ericales (M. Yan et al. 2016, 2018; X.-L. Cai et al. 2020).
Chemistry, Morphology, etc.. Benzoin or gum benjamin, which contains benzoic acid but not polysaccharides, is exuded from the resin canals of Styrax (c.f. storax, from Altingiaceae).
The floral vasculature suggests that although the stamens appear to be in a single whorl, they are basically obdiplostemonous (Dickison 1993; c.f. H.-C. Wang et al. 2010). Van Tieghem (1898) showed Halesia as having two ascending epitropous ovules and two descending apotropous ovules. Pterostyrax (and Styrax?) lack endothelium. There are no septal bundles, as in many Ericales (but details of the distribution of this character?).
For general information, see Fritsch (2004), for pollen, see Morton and Dickison (1992: little variation) and for embryology, see Shinke and Tokuoka (2020: esp. Table 2); Julio and Oliveira (2007) described the fruit, ovule, etc., of Styrax camporum.
Phylogeny. For relationships within Styracaceae, see Fritsch et al. (2001). The main phylogenetic structure in the family is [[Huodendron + Styrax] [[Alniphyllum + Bruinsmia] [The Rest]]]; both main clades, especially the second, are well supported. Members of the former clade have entire leaf blades, members of the latter have dentate blades, an inferior ovary, and bud scales, with the exception of the [Alniphyllum + Bruinsmia] clade which differs from the others on all three counts. However, the rather scanty sampling in Rose et al. (2018) suggests different relationships - [Styrax [Huodendron [[Alniphyllum + Bruinsmia] (all well supported) [Changiostyrax [Halesia + Pterostyrax]]]]]. In an analysis of plastomes M. Yan et al. (2018, 2021) obtained similar basal relationships. Both Pterostyrax and Halesia were poly-/paraphyletic, although Yan et al. (2021) found that the former was monophyletic in analyses using whole plastomes, while Parastyrax was sister to other Styracaceae. In another plastome analysis with greater sampling X.-L. Cai et al. (2020) found the same basal relationships, beyond that they recovered the topology [Styrax [Huodendron [[Alniphyllum + Bruinsmia] [Halesia [Melliodendron [[Perkinsiodendron + Rehderodendron][Changiostyrax [Pterostyrax + Sinojackia]]]]]]]] - they did not look at Parastyrax and included only one species of Halesia. Support was generally strong, except perhaps for the position of Huodendron and in particular Changiostyrax.
For relationships within Styrax see Fritsch (2001).
DIAPENSIACEAE Lindley, nom. cons. - Back to Ericales —— Synonymy: Galacinaceae D. Don
Shrublets or perennial herbs; ecto- and endomycorrhizae + [?ectendomycorrhiza]; plants Al-accumulators; ellagic acid +, iridoids?, lignans?; (stem cork cambium superficial); vessel elements with simple (scalariform) perforation plates; secondary wood rayless; pericyclic fibres 0 (+ - Shortia); nodes 3:3 (1:1 - Pyxidanthera); petiole bundle(s) arcuate to annular (medullary bundles +); (stomata anisocytic); lamina margins toothed or entire, secondary veins subpinnate to palmate; inflorescence scapose/flowers axillary; K free or connate, C forming tube along with filaments, lobes deeply serrate or not; stamens 5, opposite K, anthers ± incurved, (appendaged), thecae horizontal to vertical, filaments flattened, staminodes +/0, (A and staminodes connate - Galax); tapetal cells binuclear; (nectary +, inconspicuous), G [3], median member adaxial, style continuous with G, stigma shortly 3-lobed, wet; ovules (6-)many/carpel, apotropous, integument 5-10 cell layers across, endothelium ± 0, hypostase +; archesporium (2-3-celled), antipodal cells persistent (numerous); exotesta thick-walled, tanniniferous, rest of testa crushed; endosperm copious, embryo terete; n = 6, x = 6 (?7).
6/15: [list], Diapensia (5). Scattered N. temperate, esp. East Asia and E. U.S.A., Diapensia lapponica plus D. obovata circum-Arctic. Map: from Diels (1914), Wood and Channel (1959) and Hultén (1971). Photo: DiapensiaFlower, © J. Maunder; Fruit, © J. Maunder.
Age. Crown-group Diapensiaceae are ca 65 Ma (Hou et al. 2010) or 59.9 Ma (Rose et al. 2018). In Gaynor et al. (2020) these two ages are used as constraints to the age of the crown group, and the age of the fossil Actinocalyx, stem group, is consistent with this crown-group age.
Friis (1985) described Actinocalyx from Upper Cretaceous rocks in Sweden dated to ca 83.5 Ma (see also Martínez-Millán 2010). It has a number of similarities with extant Diapensiaceae, although the anthers are rather different, the pollen is much smaller (7-9.5 µm, versus 17-40 µm), and the styles are separate.
Evolution: Divergence & Distribution. For diversification in the family, which may have originated in the Nearctic, see Gaynor et al. (2020). Although Diapensia lapponica plus D. obovata are prominent members of the circum-Arctic flora, and the genus is also in the Himalayan—Hengduan Mountains area, it is likely that it moved north to south, diversification beginning ca 8.5 Ma, prior to the origin of the Arctic flora (Hou et al. 2016).
Plant-Bacterial/Fungal Associations. The mycorrhizal association in Diapensiaceae may be an ectendomycorrhiza like that of many Ericaceae (see Asai 1934: the distinctiveness of the ericaceous mycorrhizal association was not fully understood then; Brundrett & Tedersoo 2018).
Genes & Genomes. Servick et al. (2015) found 46 and 31 independent autotetraploidy and triploidy events respectively in Galax urceolata.
For extensive rearrangements in the plastomes of Diapensia and Berneuxia, see M. Yan et al. (2018).
Chemistry, Morphology, etc.. For ellagic acid, see Harborne and Williams (1973). The absence of rays from the family should be confirmed; there is only one old reference in Carlquist (2015b).
The integument seems to consist of an outer and inner part in some taxa, and an endothelium does not always develop (Samuelsson 1913; Diels 1914; Kapil & Tiwari 1978). Schnizlein (1843-1870: fam. 160) shows Galax with the median G abaxial.
For general information, see Scott (2004), also Palser (1963: floral morphology and anatomy), Xi and Tang (1990: pollen) and for summary of embryology, see Shinke and Tokuoka (2020: Table 2).
Phylogeny. Galax and Pyxidanthera are successively sister taxa to the rest of the family (Rönblom & Anderberg 2002; Rose et al. 2018; Gaynor et al. 2020: ...[Berneuxia [Shortia...); since this set of relationships seems to be holding, the presence of staminodes is likely to be derived within the family. For a morphological phylogeny, see Xi and Tang (1990).
Previous relationships. Diapensiaceae have often been considered close to Ericaceae, but the anthers of some genera of the former which appear to be inverted, are not.
[[Sarraceniaceae [Actinidiaceae + Roridulaceae]] [Clethraceae [Cyrillaceae + Ericaceae]]]: inflorescence racemose; K quincuncial; anthers inverting late during development, initially extrorse, opening by pores or short slits; pollen ± verrucose/rugulate ["cerebellar"], tectum and foot layer solid, infratectum with granular elements; G [3], median member adaxial, also [5], opposite C, style impressed, branched; ovules many/carpel, endothelium +; fruit a loculicidal capsule; testa with much thickened inner wall [?higher level], endosperm copious; mitochondrial coxII.i3 intron 0.
Age. Wikström et al. (2001) suggested a crown group age for this clade of around (72-)67, 59(-54) Ma, although relationships within the group are other than those shown here and Roridulaceae are way elsewhere on their tree, being sister to all other Ericales. Later, Wikström et al. (2004) noted the considerable difference between their estimate and the substantially older (ca 89 My) fossil-based estimate of Magallón et al. (1999). Bell et al. (2010: internal topology) estimated an age for this node of (68-)53, 51(-43) Ma, an age of ca 50.6 Ma is the estimate in Ellison et al. (2012), (95-)86(-77) Ma in Wikström et al. (2015), about 90.4 Ma in Magallón et al. (2015), ca 98.2 Ma in Rose et al. (2018), (63.3-)52.1(-52.0) or ca 61 Ma in Diaz-Garcia et al. (2021: [Actinidia + Vaccinium], a genome duplication event?) and 53-47 Ma in Jiao and Wang (2022).
Glandulocalyx upatoiensis, a small-flowered fossil from the Upper Cretaceous 86-84 Ma and found in Georgia, S.E. United States, has been placed near Actinidiaceae or Clethraceae (Schönenberger et al. 2012) or in a tritomy with Roridulaceae and Actinidiaceae (Martínez 2016). Florally it does seem quite a good match: It has extrorse anthers that may become introrse, placentation that becomes intrusive parietal apically, the placentae being pendant basally, hollow style branches, etc.; the 24-28 stamens are in a single whorl, but the dense perhaps glandular hairs on the outside of the sepals are rather unusual (Schönenberger et al. 2012; see also Crepet et al. 2013). Three rather older (98-94 My) species of Glandulocalyx have been described from New Jersey; the flowers are also very small, less than 2.2 mm long, and with five stamens and clawed petals, some species have nectariferous staminodes, one species has viscin threads, another stellate hairs, there is a single style and expanded stigma, and so on, again, there are usually remarkable glands on the abaxial surface of the calyx. These latter species come out somewhere in this part of the tree in morphological analyses, or associated with Diapensiaceae or even within Ericaceae (Crepet et al. 2013), but overall Glandulocalyx is both morphologically heterogeneous and rather odd.
Paleoenkianthus is another interesting Late Cretaceous fossil from some 90 Ma (Nixon & Crepet 1993). It, too, has tiny flowers; many of its features are those of a bee-pollinated flower, and bees are likely to have been around by then (Cardinal & Danforth 2013; Sann et al. 2018). Its floral morphology is odd - K 5 C (5) A 8 G (4), with "four, short slightly unequal styles/stigmas" (Nixon & Crepet 1993: p. 620), and viscin threads which, however, Friis et al. (2011) suggested might be fungal hyphae. In fact, Nixon and Crepet (1993) were not that sure of their identification, and while Friis et al. (2011) thought that this fossil was probably Ericalean, they were loath to place it more precisely.
Evolution: Divergence & Distribution. Anderberg (1992, 1993) carried out phylogenetic analyses that focussed on this clade, in the first paper drawing attention to uncommon secondary metabolites like the diterpene andromedotoxin, the triterpene ursolic acid (also in Roridulaceae), the flavonol gossypetin, and the non-carbohydrate + glucose + phenol arbutin, all rather uncommon elsewhere; his work has not been fully integrated into the phylogeny. Löfstrand et al. (2016) also suggest several apomorphies for/within this clade.
Ecology & Physiology. A considerable increase in leaf mass per area (SLA) can be placed at this node (Cornwell et al. 2014).
Chemistry, Morphology, etc.. There are pit membrane remnants in the perforations of vessels in several families here (Schneider & Carlquist 2003, 2004; Carlquist & Schneider 2005).
Bracteole presence is variable (Anderberg & Xiaoping 2002). For a summary of pollen variation, see X-P. Zhang and Anderberg (2002). The particular time during development that the anther inverts varies within this clade, and even the direction in which it occurs - thus in Sarraceniaceae the inversion is introrse → extrorse, but usually it is in the opposite direction (Schönenberger et al. 2012). How this character evolved - indeed, what this character "is" - are both unclear (Löfstrand & Schönenberger 2015a; Löfstrand et al. 2016).
Phylogeny. Family-level relationships within this clade are well supported (e.g. Löfstrand and Schönenberger 2015b; Löfstrand et al. 2016, etc.). However, Z.-D. Chen et al. (2016) found the grouping [Clethraceae [Actinidiaceae + Ericaceae]] in a study of Chinese taxa while Larson et al. (2019/2020: Clethraceae and Cyrillaceae not included) found some support for the position of Sarraceniaceae as sister to the whole group.
[Sarraceniaceae [Actinidiaceae + Roridulaceae]]: route I secoiridoids +; K unequal, C quincuncial, at base thicker than sepals; stamens many [?here]; nectary 0; G opposite K [when 5], stigma dry, papillate [?level]; ovule hypostase +; K persistent in fruit.
Age. The age of this node is estimated to be (53-)48.6[47](-43) Ma (Ellison et al. 2012), ca 70.1 Ma (Tank et al. 2015: Table S2), 88.1 Ma (Magallón et al. 2015) or ca 93.4 Ma (Rose et al. 2018).
Evolution: Divergence & Distribution. Chartier et al. (2017) thought that this clade was morphologically rather homogeneous in the context of the general floral morphospace of Ericales; this would obviously not have been the case if vegetative variation had been examined.
See Löfstrand and Schönenberger (2015a, b) and Löfstrand et al. (2016) for possible apomorphies. "Many stamens" is probably a feature derived independently within Sarraceniaceae and Actinidiaceae.
Pollination Biology. Buzz pollination is scattered throughout this clade, but nectaries are found in some taxa, whether on the ovary or anther (for buzz pollination and nectar production, see also Boraginaceae).
Chemistry, Morphology, etc.. For a detailed study of floral morphology of the whole clade, see Löfstrand and Schönenberger (2015a). They describe all three families as having hydrolyzable tannins in the floral tissue and condensed tannins in vesicles; I do not know what these are chemically.
SARRACENIACEAE Dumortier, nom. cons. - Back to Ericales —— Synonymy: Heliamphoraceae Chrtek, Slavíková & Studicka
Herbs, rosette-forming, carnivorous [insectivorous]; O-methyl flavonols only +, monoterpene sarracenin +; mycorrhizae 0; cork?; vascular bundles initially separate; nodes ?; leaves with broad bases, pitcher +, terminal [epiascidiate], with lid/nectar spoon, (adaxial drainage hole/slit - Heliamphora - H.); inflorescence scapose (flowers solitary), bracteoles + [H.]; K ± C-like, (3-6), quincuncial, (nectar spoon at apex - H.), C (0 - H., 4), free; A 8-10 [H.] or many, centrifugal [ten groups, two opposite each C - Sarracenia - S.], anthers introrse, inversion late/unclear, with slits (basal pores); pollen (3-)4+ colporate, surface verrucose-vermiculate, with small granules; nectary (on ovary wall - S.), G opp. K, placentation intrusive parietal apically [placentae long-stalked V-shaped], style (not impressed), apically hollow or not, apex divided or peltate-expanded, stigmas small, dry; ovules many/carpel, unitegmic, integument 9-10 cells across, (bitegmic, micropyle endostomal, outer integument 3-4 cells across, inner integument 4-5 cells across - H.), "incompletely tenuinucellar"; seeds small, with wings/hairs, exotesta ± thickened; endosperm haustoria?, embryo medium; n = 13, 15, 21, x = 14 (?15), nuclear genome [1C] (1.304-)3.834(-11.275) pg / 1110-3597 mbp.
3/32: [list], Heliamphora (23+). E. and W. U.S.A. and the Guayana Highlands. Map: from Uphof (1931) and Schnell (2002). [Photo - Inflorescence © J. Maunder, Flower, Flower.]
Age. The age of crown group Sarraceniaceae is estimated to be (44-)35(-25) (Ellison et al. 2012), ca 48.5 (Rose et al. 2018) or ca 55 (S. Liu &anp; Smith 2020) Ma.
Archaeamphora longicervia, described from rocks in northeastern China ca 124.6 Ma and thought to be the pitchers of a member of Sarraceniaceae (H. Li 2005a, b) - there were also supposedly seeds in the deposits that were associated with it - is actually more likely to be a gall of the conifer Liaoningocladus boii (W. Wong et al. 2015; see also Herendeen et al. 2017).
Evolution: Divergence & Distribution. Ellison et al. (2012) offered a vicariance-style explanation that integrates the relationships and distributions of the three genera; Sarraceniaceae originated in South America, perhaps hopping across the proto-Caribbean on islands. However, given the topology of relationships of the three genera (see below) and the general uncertainty of broader-scale relationships, the biogeographic here seem unclear. S. Liu and Smith (2020) looked at relationships, distribution, etc., in Heliamphora; the crown group age there is 18-17 Ma.
Ecology & Physiology. Carnivory: There are nectar glands on the pitcher - in the nectar spoon above the pitcher in Heliamphora - which attract insects that then fall into the pitcher, alternatively, the nectar may take up water increasing the possibility of an insect's hydroplaning into the pitcher (see Bauer et al. 2008); either way, death by drowning is the result, retrorse hairs on the inside of the tube preventing the prey from climbing out. Suggestions that the colouring on the flap of the pitcher may attract insects, that is, it is a kind of pseudoflower (Cresswell 1993), seem unlikely (Joel 1988; Ruxton & Schaefer 2011), however, the plants of both Sarracenia and Heliamphora are variously scented, so helping to attract potential prey (Fleischmann 2016); for more on pitcher morphology, see Thorogood et al. (2017). Martin-Eberhardt et al. (2025) suggested that there was a long-term stability in the colour polymorphism of S. purpurea, where red plants might catch fewer prey, but they also caught fewer pollinators, while green plants were much more likely to capture the pollinator Bombus impatiens. Such reddening was in fact common in carnivorous plants, and anthocyanin loss had pleiotropic effects. The situation is complex. Thus West-Eberhardt et al. (2026) found that in an experimental study using S. purpurea as a model the colour red and nectar (this reflects UV light) additively increased the diversity of prey and the numbers of the ant Myrmica lobifrons in particular that were captured, but the inclusion of volatile organic compounds had little or no effect.
The pitcher fluid varies in the amount of digestive enzymes it contains, although these enzymes seem to come from the organisms in it, not from the plant; this is a case of symbiotic digestion (Peroutka et al. 2008b), and in Sarracenia purpurea, for example, there are few enzymes. Rainfall in the area where Heliamphora in particular lives can be heavy and the pitchers are open, however, slits/holes on the adaxial surface allow water to drain from the pitchers, and appropriately-placed hairs keep particulate matter, e.g. decomposing insects, from being lost in the process (S. Liu & Smith 2020). Nutrients from the entrapped animals are made available to the plant by the activity of detritivores that break up the animals, further decomposition is carried out by bacteria, and these in turn are eaten by rotifers and protozoa and ultimately by mosquito larvae, and mites, ciliates and flagellates are also to be found here - all forming a microcosm in each pitcher, even if there can be considerable interpopulational variation in the particular denizens of a pitcher (Kitching 2000; Ellison et al. 2003; Butler & Ellison 2007; Adlassnig et al. 2011; Grothjan & Young 2019). Interestingly, Cueller-Gempeler et al. (2025) found that although the diversity of microbes in the pitchers of S. purpurea decreased with the age of the pitcher, the rate of breakdown of its contents increased. Actual uptake of nutrients by the plant is by endocytosis carried out by epidermal cells, the so-called digestive epithelium (Koller-Peroutka et al. 2019; Freund et al. 2022); there are no glands, common in other carnivores, including Byblidaceae. Schulze et al. (1997) noted that 76.4±8.4% of the nitrogen in the pitchers of Darlingtonia californica ultimately came from insects, about the highest figure in carnivorous plants. For general information on carnivory, see especially Lloyd (1942) and Juniper et al. (1989), and for recent developments, see Hatcher et al. (2020) and Adamec et al. (2021).
Pollination Biology. Flowers of Heliamphora have nectaries and are buzz pollinated; Sarracenia has ten nectaries on the ovary wall above the stamen fascicles.
Plant-Animal Interactions. Caterpillars of the moth Exyra fax drain the pitchers of Sarracenia by opening up a hole at the base, and they then eat the pitcher. Exyra occurs throughout the range of Sarracenia (see e.g. McPherson & Schnell 2011).
For the microcosm inside the pitcher, see Bittleston (2018 and references), also above. The small mosquito Wyeomyia smithii breeds in the pitchers of Sarracenia purpurea, and the larvae eat animal remains in the pitcher although they are not harmed by the fluid there (Bradshaw 1983; Istock et al. 1983). The recent range expansion of the mosquito as the climate warms and its adaptation to the changing daylengths it consequently faces have been much studied (Mathias et al. 2007 and references). Interestingly, the diversity of animals, whether invertebrates or bacteria, in the pitchers increases with increasing latitude, the reverse of the normal trend (see elsewhere), perhaps because the numbers of Wyomyia larvae that eat them decrease (Buckley et al. 2003; Kindlmann et al. 2007).
Plant-Bacterial/Fungal Associations. Sarraceniaceae lack mycorrhizae (Brundrett 2004 and references).
Vegetative Variation. Franck (1976) provides an evaluation of classical work on the nature of the pitcher. In Sarracenia the leaves have an adaxial flange, but the pitcher develops from the midrib area. Fukushima et al. (2015) suggest that patterns of cell division affect the development of the leaf and of the pitcher shape, K. J. I. Lee et al. (2019) noted that their mechanism for trap growth in Utricularia (q.v.) involved a polarity field propagating through a curved sheet rather than cell division, while Whitewoods et al. (2020) reconstruct the morphology of the pitcher following the principles they used when discussing the development of Utricularia traps.
Genes & Genomes. For genome size here, see Veleba et al. (2020).
Chemistry, Morphology, etc.. Jensen (1992) suggested that the family has route I iridioids.
Löfstrand and Schönenberger (2015a) thought that the perianth of Heliamphora was biseriate, being made up of calycine and corolline whorls. Outside the petaloid calyx of Sarracenia there are three "bracts". When there are many stamens, development is sometimes centrifugal from initially 10 primordia (Guo & Halson 2020b).
For general information, see Kubitzki (2004b), McPherson (e.g. 2006, 2008b, 2010), papers in Ellison and Adamec (2018), esp. Naczi (2018), and the Carnivorous Plants Database, for perforation plates, see Schneider and Carlquist (2004) and Carlquist (2012c: pores almost closed), for pollen, see Takahashi and Sohma (1981) and for stylar, etc. development, see Guo and Halson (2020a).
Phylogeny. R. J. Bayer et al. (1996), Neyland and Merchant (2006), Ellison et al. (2012), Rose et al. (2018) and S. Liu and Smith (2020) provide information about relationships within the family; the topology [Darlingtonia [Sarracenia + Heliamphora]] was well supported. Liu and Smith (2020) discuss relationships in Heliamphora and Stephens et al. (2015) relationships and species limits in Sarracenia.
Classification. McPherson and Schnell (2011) and McPherson et al. (2011) provide an account of the family. There is extensive interspecific hybridization in both Heliamphora and Sarracenia.
Thanks. I thank D. Hoekman for information.
[Actinidiaceae + Roridulaceae]: raphides + [in sacs]; mucilage in stylar canal/on placentae, inner surface of carpels secretory, lateral carpellary vascular bundles absent [no synlaterals].
Age. Stem-group Roridulaceae have been estimated to be ca 90 Ma old (Warren & Hawkins 2006) or ca 38.1 Ma (Ellison et al. 2012); the age for this node is about 84.8 Ma in Magallón et al. (2015), 79.5 Ma in Rose et al. (2018), and (85-)76(-72) Ma in Wikström et al. (2015).
Chemistry, Morphology, etc.. See Löfstrand et al. (2016) for a discussion on synlateral vascular bundles and gynoecial mucilage.
ACTINIDIACEAE Engler & Gilg, nom. cons. - Back to Ericales —— Synonymy: Saurauiaceae Grisebach, nom. cons.
Trees, shrubs or twining lianes; (vessel elements with simple perforation plates); (nodes 3:3); petiole bundle deeply arcuate with wing bundles [Actinidia] or annular (medullary bundles +); stomata anomocytic; hairs multiseriate, often ± (flattened) setose; leaves (opposite), lamina vernation conduplicate, apex of tooth expanded, clear, not deciduous, (secondary veins subpalmate); plant cryptically dioecious/monoecious; K quincuncial, C ± connate or not, (?nectary at base); A 10-many, centrifugal, (in groups opposite C), inflexed in bud (not - Saurauia), (basally connate), anthers extrorse, inverting, becoming introrse and dehiscing by pores or ± short slits, opening at morphological base of anther; pollen tectum ± psilate to rugulate and transversely ridged, (equatorial bridge of ektexine over endoaperture); (?nectary on G); G [(-20)], styluli + (very short - Saurauia staminate flowers), or style +, ± branched (unbranched), (grooved - Actinidia), (hollow), (stigma capitate, peltate, lobed); ovules 10</carpel, integument 6-9 cells across, parietal tissue ?0-ca 3 cells across, nucellar cap 0-ca 3 cells across; (megaspore mother cells several), embryo sac protruding from nucellus; fruit usu. a berry (loculicidally dehiscent); seeds embedded in placental pulp; integument multiplicative; endosperm haustoria?; n = 24, 29, 30, 39, chromosomes ca 1μm long [Actinidia]; x = 24, nuclear genome [1C] (680-)1242(-2176) Mb/(0.178-)1084(5.656) pg; (plastid transmission biparental); horizontal transfer of mitochondrial rps2 gene [Actinidia].
3/430: [list]. Saurauia (350), Actinidia (54), Clematoclethra (25). Largely tropical, esp. South East Asia to Malesia, but not Africa (map: from Soejarto 1980).
Age. The crown-group age of this family is ca 46.5 Ma (Rose et al. 2018).
Parasaurauia was described from flowers of Early Campanian (Late Cretaceous) age ca 80 Ma from the eastern USA. It has impressed, separate styles and numerous stamens and may belong to crown group Actinidiaceae (Keller et al. 1996; Herendeen et al. 1999; see also Martínez-Millán 2010).
Evolution: Divergence & Distribution. For the fossil record of Actinidiaceae, see Manchester et al. (2015).
Both separate styles and numerous stamens are probably derived within the family (Keller et al. 1996; Herendeen et al. 1999).
Genes & Genomes. For a the Ad-α/ACCHα genome duplication, perhaps in Actinidia only, see Shi et al. (2010: ca 28.3 Ma), S. Huang et al. (2013: ca 26.7 Ma), Leebens-Mack et al. (2019), Larson et al. (2019/2020) and Qiao et al. (2019), but c.f. Q. Zhang et al. (2022) and see e.g. Theaceae, etc. above. J.-P. Wang et al. (2018) suggest that there were two autotetraploidy events in Actinidia or its immediate ancestors, one 57-50 Ma and the other 20-18 Ma.
Hybridization has been common in Actinidia. Palaeopolyploidy is pervasive, and He et al. (2005) proposed that x = 12 in Actinidiaceae, but there are few counts for Saurauia.
Akagi et al. (2019) looked at sex determination in Actinidia, and found that it was controlled by two genes on the Y chromosome; see also Akagi and Charlesworth (2019), who noted the pleiotropic effects of the sex-determinig genes.
There is paternal transmission of the plastid genome in Actinidia (Chat et al. 2003), maternal in Clematoclethra, and the clP gene is missing in both (W.-C. Wang et al. 2018).
Economic Importance. For the vitamin C-rich kiwi fruit, Actinidia deliciosa, see H. Huang (2014).
Chemistry, Morphology, etc.. There seems to be some confusion over the dehiscence of the anthers. Dressler and Bayer (2004: p. 15) talk about "longitudinal slits or pores that become extrorse by the inversion of the anthers", but their Fig. 4 shows the opposite, while Soejarto (1969) was similarly unclear, suggesting that the result of anther movement in Saurauia was that the pollen would be dispersed away from the stigma - the reverse is the case as his Fig. 23A-F shows. Anthers of staminodes in Saurauia contain sterile pollen; in general, dioecy in the family is cryptic, although Soejarto (1969) notes that male flowers of Saurauia that he examined had very short styles, female flowers had long styles - heterostyly associated with sex (see also Henry et al. 2018). For androecium development in Actinidia, see van Heel (1987), the synascidiate carpels are in a single whorl, and there is a large, flat, residual floral axis.
For general information, see Dickison (1972), Dressler and Bayer (2004) and Wong (2017: Saurauia), for vegetative anatomy, see Beauvisage (1920), the floral anatomy of Actinidia, see Schmid (1978), for floral development, see Schönenberger et al. (2012), for pollen, see Dickison et al. (1982) and X.-P. Zhang and Anderberg (2002), and for ovules, see Guignard (1882) and Schnarf (1924).
Phylogeny. Rose et al. (2018) suggest the relationships [Saurauia [Clematoclethra + Actinidia]]. Y.-F. Liu (2017) and Tang et al. (2019) looked at relationships within Actinidia where there has been extensive hybridization; the former suggest there was a rapid radiation producing what is now a backbone phylogeny, hybridization occurring between the species that resulted. Within Saurauia, there are sister Neotropical and Asian-Pacific clades (Löfstrand & Schönenberger 2015b), but the genus has been little studied.
RORIDULACEAE Martinov, nom. cons. - Roridula L. - Back to Ericales
Shrubs, carnivorous [insectivorous]; unspecified iridoids +, ellagic acid?; cork?; pericyclic fibres 0; hairs dense, glandular; leaves ± U-shaped in t.s., sessile, lamina linear, margins entire or laciniate; inflorescence with a terminal flower [?always; sometimes looking racemose], bracteoles +; C free; stamens = and opposite K, connective swollen at apex of anther, conspicuous and nectariferous, anthers latrorse; pollen densely and minutely spinose and microreticulate, or surface irregular, infratectum subcolumellate; placentation apical, style unbranched, apical part of style/stigma clavate, densely long-papillate; ovules 1-4/carpel, pendulous, integument 10-11 cells across, funicle prominent; testa mucilaginous [?]; endosperm with micropylar haustorium +; n = 6, x = 6 (?7), nuclear genome [1C] (0.054-)1.128(-23.459) pg.
1/2: [list]. Southern Africa. Map: fossil locality in green). [Photo - Roridula Flower © M. Schmidt.]
Age. The age of crown-group Roridulaceae is estimated to be ca 10.7 Ma (Ellison et al. 2012) or ca 20.2 Ma (Rose et al. 2018).
Evolution: Divergence & Distribution. It was suggested that, given their age, ca 90 Ma, Roridulaceae must be very much a relictual (?Gondwanan) element in the Cape flora (e.g. Warren & Hawkins 2006). However, well preserved fossils in Baltic amber dated to 47-35 Ma were recently described by Sadowski et al. (2015), furthermore, the general distributions of families in this part of the ericalean tree are not Gondwanan. Indeed, Roridulaceae might even have originated in the northern hemisphere; see also also Cornales-Curtisiaceae for comparable extant/fossil distributions. In any event, this is definitely a low-diversification clade (Magallón et al. 2018)!
Ecology & Physiology. Although the family may not be carnivorous in a conventional sense, digestive enzymes not having been recorded from it (e.g. Hartmeyer 1997), its two species live in a very close mutualistic association with two species of the hemipteran mirid bug, Pameridea (see Wheeler & Krimmel 2015 for the bugs). These bugs eat the insects that get stuck to the hairs that cover the plant, the assisted carnivory of Obregon (2017: p. 74), and the plant absorbs nutrients from their excreta via tiny holes in the cuticle, a form of indirect carnivory (Ellis & Midgley 1996; Anderson 2005). However, Plachno et al. (2009) suggest that Roridula is directly carnivorous when they recorded mineral uptake from Drosophila stuck on the leaves. Indeed, it is not totally clear what is going on here, since although the glandular hairs of the plant produce a lipophilic resin that is sticky even underwater, there are no tanks or their like here - or maybe it helps in catching wet prey, furthermore, spiders may also be involved (see Freund et al. 2022 for literature).
Pollination Biology. The bug Pameridea may also be involved in pollinating Roridula (Ellis & Midgley 1996). Most pollination is by autonomous selfing or (25-68%) selfing or geitonogamy by immature bugs, buzz pollination being at most uncommon and unimportant despite the apparent buzz-pollination syndrome of the flowers (Anderson et al. 2003). See also Mabberley (2017) for suggestions about pollination mechanisms.
Plant-Bacterial/Fungal Associations. The plant may lack mycorrhizae (Brundrett 2004; Conran 2004 and references; c.f. Adlassnig et al. 2005).
For general information, see Vani-Hardev (1972), Dahlgren in Dahlgren and van Wyk (1988: endosperm haustoria 0), Conran (2004), McPherson (2008, 2010), papers in Ellison and Adamec (2018), and the Carnivorous Plants Database, also Wilkinson (1998: anatomy) and Takahashi and Sohma (1981) and X.-P. Zhang and Anderberg (2002), both pollen.
Previous Relationships. Roridula was included in Byblidaceae by Cronquist (1981); for further information on relationships, see that family (Lamiales!).
[Clethraceae [Cyrillaceae + Ericaceae]]: ellagic acid +; cork cambium ± pericyclic; pericyclic fibres absent; leaves spiral; bracteoles 0; stamens = 2x K, anthers extrorse, inverting, becoming introrse and dehiscing by pores or ± short slits, developing at morphological base of anther; nectary well developed, in basal part of ovary wall; placentation intrusive parietal apically, basal part of placenta free, pendulous, style hollow; embryo sac narrow at the chalazal end; endosperm with micropylar and chalazal haustoria, embryo terete.
Age. The age of this node is estimated to be ca 58 Ma (Naumann et al. 2013), ca 50.6 Ma (Ellison et al. 2012) or (89-)71(-44) Ma (Wikström et al. 2015); see sampling in all. Around 93.9 Ma is the estimate in Rose et al. (2018), 79.2 Ma in Magallón et al. (2015) and ca 70.4 Ma in Tank et al. (2015: Table S2).
Rariglanda jerseyensis, a 92-85 Ma fossil from New Jersey, has dense multicellular hairs on the abaxial surface of the calyx; in an analysis of 13 morphological characters ("anther apical protrusion" means what in the three species so coded?) it was placed with the single species of Clethra in the analysis (Martinez et al. 2016). If correctly placed, most molecular ages proposed in this clade will have to be reconsidered - but Ericales were only one of the eight orders in which the fossil ended up in the analyses of López-Martínez et al. (2023a: Table 3 - Malpighiales with strong support). Silvestro et al. (2021) estimated that the time-of-origin of Clethraceae was about 120.6 Ma - again, if correct, other ages are underestimates.
Evolution: Divergence & Distribution. In the context of the floral morphospace of Ericales, Chartier et al. (2017) thought that this clade was rather homogeneous. For embryological apomorphies here, see Maheshwari (1950) and Palser (1963).
Chemistry, Morphology, etc.. It is possible that the accumulation of sugars as ketose and isokestose oligosaccharides is of systematic significance; fructoses may be involved in membrane stabilization and cold- and/or drought tolerance in plants (Livingston et al. 2009). Bracteoles have to be regained somewhere in this clade, but I have not worked out where - perhaps it is not that important.... Van Tieghem (1903) noted that both Clethraceae and Ericaceae had ovules with an epistase.
Phylogeny. The relationships [Cyrillaceae [Clethraceae + Ericaceae]] are sometimes recovered (Morton 2011: nuclear Xdh gene).
Previous Relationships. Members of this group have often been associated in the past - the "true" Ericales. Thus Palser (1961) included the various families that now make up Ericaceae, along with Clethraceae, and perhaps also Diapensiaceae, Lennoaceae (neither now in this area), Empetraceae and Cyrillaceae.
CLETHRACEAE Klotzsch, nom. cons. - Back to Ericales
Plant (deciduous); mycorrhiza a modified ectendomycorrhiza?; fructan sugars accumulated as kestose and isokestose oligosaccharides [levans and inulins], iridoids?; (pits vestured); petiole bundle arcuate or annular with medullary bundle; stomata also paracytic and actinocytic; hairs stellate [Clethra sect. Clethra]; lamina vernation conduplicate-subplicate, margins toothed (entire); inflorescences terminal, branched or not; flowers spreading (pendulous), (bracts conspicuous - Purdiaea), pedicels articulated; C basally connate or free; A obdiplostemonous [?all], adnate to C or not, anthers ± sagittate, dehiscing by pores or short slits; pollen <20µm, oblate, psilate to rugulate, granules +/0; nectary +, not vascularized/0; G ?orientation, (placentation apical - Purdiaea), stigma lobed or not; (ovule 1/carpel, pendulous, straight - Purdiaea); K persistent in fruit; seeds winged or not, or fruit dry, indehiscent, testa undistinguished, ± disappearing [Purdiaea]; endosperm hemicellulosic; n = 8, x = 8, nuclear genome [1C] (0.051-)1.329(-34.633) pg.
2/75: [list], Clethra (65). E. Asia to Malesia, S.E. U.S.A. (sect. Clethra), Mexico southwards, Cuba, 1 sp. on Madeira (Clethra sect. Cuellaria); largely tropical montane to ± warm temperate (map: from Sleumer 1971d; Good 1974; Heywood 1978). [Photos - Fruits & Flowers © A. Gentry, Inflorescence, Purdiaea Inflorescence.]
Age. Crown-group Clethraceae are ca 82.8 Ma (Rose et al. 2018).
Evolution: Divergence & Distribution. Grehan (2017) suggested that the distribution Clethra arborea/Macaronesia — C. alnifolia/east North America reflects vicariance events as the North Atlantic opened 120-100 Ma. Dates around here seem particularly unclear (for more in Clethra, see Rose et al. 2018), however, Palaeo-Macaronesia is likely to be 60 Ma or more old, the oldest currently emergent Canary Island dating to ca 21 Ma (Gelmacher et al. 2005; Fernández-Palacios et al. 2011) so at least some role for dispersal of Clethra is likely.
Sorokin et al. (2025) looked at the fruit anatomy and development in species of Clethra, linking them with fruits in Ericaceae. They found that most species of Clethra had fruits of the Hamamelis type in which both the inner mesocarp and endocarp were sclerenchymatous; such fruits were also to be found in Enkianthus, Pyrola rotundifolia, Chamaedaphne and Oxydendrum. On the other hand, fruits of C. arborea had fruits of the Lilium type, the endocarp alone being sclerenchymatous, as in the fruits of the ericaceous Moneses and Monotropa (Sorokin et al. 2025).
Ecology & Physiology. Purdiaea nutans locally dominates in ridge-line vegetation in Ecuador (Mandl et al. 2008).
Chemistry, Morphology, etc.. Iridoids were described as being absent but scored as being present in Hufford (1992); they are not mentioned by Schneider and Bayer (2004). In Clethra, there is a prominent endodermis in the stem and the pith tends to be heterogeneous.
Some information is taken from J. L. Thomas (1960: Purdiaea), Sleumer (1967b: Clethra), and Schneider and Bayer (2004), all general, Sai 1934 (mycorrhiza), Giebel and Dickison (1976: wood anatomy), Kavaljian (1952) and Schönenberger et al. (2012), both floral morphology, and X.-P. Zhang and Anderberg (2002) and Anderberg and Zhang (2002) both pollen.
Phylogeny. See Fior et al. (2003) for a phylogeny of Clethra; they suggest that the Macaronesian C. arborea may be sister to the E. North American C. alnifolia.
[Cyrillaceae + Ericaceae]: myricetin +; colleters +; K ?imbricate, C connate; stigma wet.
Age. The age of this node is estimated to be ca 93.9 Ma (Rose et al. 2018), 69.3 Ma (Magallón et al. 2015) or ca 64.7 Ma (Tank et al. 2015: Table S2).
CYRILLACEAE Lindley, nom. cons. - Back to Ericales
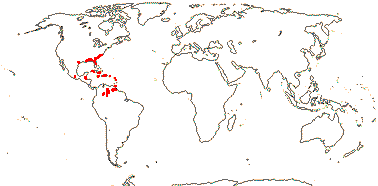
Shrub (deciduous), shooting from roots; iridoids?; sieve tube plastids with protein crystalloids and fibres; petiole bundle annular, complex or deeply concave; colleters + [Cyrilla]; lamina vernation supervolute, margins entire; flowers spreading, (weakly monosymmetric), (6-7-merous), pedicels articulated; K connate basally, C connate basally; A diplostemonous, or = and opposite sepals [Cyrilla], anthers ellipsoid, introrse, not inverting, dehiscing by slits; pollen >16µm, spherical, psilate [smooth], granules +; G [2-5], placentation apical, style 0 or short, continuous with ovary, ?solid, stigma lobed; ovules 1-3/carpel, pendulous, mostly apotropous, integument 4-7 cells across; fruit indehiscent, 1-4-seeded, dry drupe or 2-5-winged samara; testa undistinguished, ± disappearing; endosperm moderate; n = 20, x = 6 (?7).
2/2: [list]. S. U.S.A. to N. South America. Map: from Thomas (1960). [Photo - Cliftonia Habit].
Age. Crown-group Cyrillaceae may be ca 19.6 Ma (Rose et al. 2018).
Chemistry, Morphology, etc.. Goldberg (1986) notes the presence of small, scarious stipules; I have not seen them. J. L. Thomas (1960: p. 15) described "bright red, ligulate, glandular structures" associated with the axillary buds, and there were perhaps similar structures paired at the bases of the bracts and even sepals; he was unsure of their nature. Although it is unclear to what he was referring, it was probably Cyrilla.
Goldberg (1986) also shows a floral diagram in which the median K is abaxial. The sepals are small and do not overlap, except perhaps very early in development. Anderberg and Zhang (2002: see also Copeland 1953) draw the anthers as being introrse and also suggest that the stamens do not invert during development. Cyrilla was described as having its five stamens opposite the petals by Thomas (1960). There are stomata on the nectary, but apparently not on the nectaries of Ericaceae (W. H. Brown 1938).
For general information, see J. L. Thomas (1960: monograph, 1961), Anderberg and Zhang (2002) and Kubitzki (2004b), also Copeland (1953: floral morphology and anatomy), X.-P. Zhang and Anderberg (2002: pollen), Vijayaraghavan (1970: ovule morphology, etc.).
Previous Relationships. The old division between Clethraceae and Cyrillaceae was based on fruit type (dehiscent versus indehiscent fruits), but the new limits correlate better with general floral morphology.
ERICACEAE Jussieu, nom. cons. - Back to Ericales
Benzo- and naphthoquinones, route I secoiridoids +, ellagic acid 0; petiole bundle arcuate; pericyclic fibres in leaf and stem poorly/not developed; buds perulate [?level]; lamina vernation involute, margins entire to toothed, teeth associated with multicellular hairs; inflorescence terminal, flowers often ± pendent; K connate basally, C connate; A obdiplostemonous, appendages +, (resorbtion tissue/granular pouches +); tapetal cells uni- or binucleate; pollen >26µm, 3(-5) colporate; G [5], opposite C, style undivided, impressed in ovary [?level]. stigma expanded; integument 4-6 cells across; K persistent; exotesta with outer wall unthickened; x = 6 (?7), nuclear genome [1C] (0.102-)1.738(-29.75) pg; chloroplast infA gene defunct.
126/4,360 (4,426: Schwery et al. 2014): [list: to tribes] - nine subfamilies below. Boreal to warm temperate, also montane tropics, very rare in lowland tropics. Map: N. part of range, see Hultén (1971), also Meusel et al. (1978) and Luteyn (1995).
Age. The age of this node may be around ca 90.5 Ma (Rose et al. 2018), 98-90 Ma (Z.-W. Liu et al. 2014) or (123.9-)117.3(-109.4) Ma (Schwery et al. 2014), although many estimates of the age of core eudicots as a whole are similar to this last age...
Fossils of capsules from deposits in the Fort Union Formation in Wyoming have recently been described as Sandrawia scottii by Tiffney et al. (2025); although probably ericaceous, they cannot be identified more precisely.
Includes Andromedeae, Arbutoideae, Archerieae, Bryantheae, Cassiopoideae, Cosmelieae, Empetreae, Enkianthoideae, Epacrideae, Epacridoideae, Ericeae, Ericoideae, Gaultherieae, Harrimanelloideae, Monotropeae, Monotropoideae, Oligarrheneae, Oxydendreae, Phyllodoceae, Prionoteae, Pterosporeae, Rhododendron subgenera Rhodoreae, Richeeae, Styphelieae, Vaccinieae, Vaccinioideae.
1. Enkianthoideae Kron, Judd & Anderberg - Enkianthus Loureiro
(Plant deciduous); pith with small, thick-walled and lignified and larger and thin-walled cells mixed [heterogeneous]; leaves pseudoverticillate; inflorescence buds perulate; bracteoles 0; anthers dehiscing by slits broadening towards the apex, with paired awns; pollen grains tricellular, (hexacolporate), surface ± granulate/microspinose; ventral carpellary bundles in septal plane; suprachalazal nucellar tissue; megagametophyte with "ears"; n = 11 (?12).
1/16. South East Asia: China, Japan and environs. Map: from Kron and Luteyn (1995). Photo: Habit.
Age. Crown-group Enkianthus is ca 11.3 Ma (Rose et al. 2018).
For Paleoenkianthus, ca 90 Ma but in fact unlikely to be Ericaceae, see above.
[[Pyroloideae [Monotropoideae + Arbutoideae]] [[Cassiopoideae + Ericoideae] [Harrimanelloideae [Epacridoideae + Vaccinioideae]]]]: plant ectendomycorrhizal, [fungal sheath; Hartig net; hyphae with complex coiled intrusions in the exodermal cells], root hairs 0; K with single trace [?always]; anthers with exothecium, endothecium 0 [?here], dehiscing by pores; pollen in tetrahedral tetrads, tetrads calymmate, cohesion simple, surface ± rugulate; stigma dry to wet; vascular bundle in ovule absent; nuclear genome [1 C] (577-)735(-1252) Mb; duplication of complete chloroplast ndhH-D operon.
Age. This node is dated at around 91 Ma (Z.-W. Liu et al. 2014).
[Pyroloideae [Monotropoideae + Arbutoideae]]: root with hyphal mantle, Hartig net common, ECM-type fungi, commonly inc. Sebacinales-Sebacinaceae.
Age. The age of this node is about 86 Ma (Z.-W. Liu et al. 2014).
2. Pyroloideae Kosteltsky —— Synonymy: Pyrolaceae Lindley, nom. cons.
Perennial herbs, rhizomatous, (plant mycoheterotrophic; leaves as scales); fructan sugars accumulated as ?kestose and isokestose oligosaccharides [levans and inulins]; multicellular hairs on leaves 0; leaves pseudoverticillate; flowers (monosymmetric by style curvature, asymmetric); bracteoles 0; C free, ± rotate (± campanulate); anthers with short/0 tubules, appendages 0, (endothecium 0); (pollen in monads/polyads); nectary +/0; placentation intrusive parietal, (style 0 - Chimaphila), stigma ± peltate-lobed; integument 2-3 cells across; seeds small [≤1 mm long], testa cell walls thin; embryo short, hardly differentiated; germination mycoheterotrophic; n = 8, 11, 13, 16, 19; protein crystals in nuclei.
4/40: Pyrola (35). N. hemisphere, temperate to arctic, in N. Sumatra. Map: from Meusel et al. (1978) and Hultén and Fries (1986); the distribution in E. Asia is rather unclear. [Photo - Chimaphila Flower, Pyrola Flower.]
Age. The age of crown-group Pyroloideae is estimated to be (70.9-)50.7(-33.5) Ma (Z.-W. Liu et al. 2014) or ca 44.5 Ma (Rose et al. 2018).
[Monotropoideae + Arbutoideae]: ?
Age. The two clades diverged ca 70 Ma (Hardy & Cook 2012) or around 79 Ma (Z.-W. Liu et al. 2014).
3. Monotropoideae Arnott
Perennial herbs, roots fibrous/coralloid/forming a tight ball; plant echlorophyllous, holomycoheterotrophic [= hyperparasitic]; fungal hyphae with peg-like intrusions into exodermal cells; fructan sugars accumulated as kestose and isokestose oligosaccharides [levans and inulins]; (root cap 0); vascular cambium 0 [?all]; sieve tube plastids lacking both starch and protein inclusions; leaves sessile, ± scale-like; bracteoles 0; anthers with ± longitudinal dehiscence, etc.; pollen in monads; integument 2-3 cells across; (fruit a berry); embryo undifferentiated, with 30-50 cells; protein crystals in nuclei; chloroplast genome 35.1-45.1 kb, inverted repeat absent (very small); germination mycoheterotrophic.
11/13. N. hemisphere, largely temperate; to Colombia, Malaya and Sumatra. Map: from Wallace (1975) and Hultén and Fries (1986).
Age. Crown-group Monotropoideae are ca 65.8 Ma (Rose et al. 2018).
3A. Pterosporeae Baillon
Multicellular multiseriate glandular hairs +; inflorescences many-flowered; anther spurs paired/0; pollen 4-colpor(oid)ate; fruit rather irregularly dehiscent/loculicidal; seeds with large wing/subovoid, sticky; suspensor uniseriate; n = 8.
2/2. S.W. Canada, W. U.S.A. to Central Mexico, N.E. U.S.A. and adjacent Canada. Photo: Pterospora Habit, Flower.
3B. Monotropeae Dumortier —— Synonymy: Hypopityaceae Klotzsch, Monotropaceae Nuttall, nom. cons.
Plant usu. blackening on drying; multicellular hairs 0; inflorescences 1-many flowered, (inflorescence sessile); flowers 3-8-merous; (bracts petaloid), (bracteoles +?); K (0), (quite large), C (0), free (connate); anthers also hippocrepiform, thecae confluent, with apical slit, spurs 0; pollen 2-3(-4)colporoidate (3-4-porate); (placentation parietal), (style unvascularized); fruit a berry/loculicidal capsule; seeds minute, dust-like, (exotestal walls massively thickened); n = 13, 16, 24, genome (1 C - Monotropa) 29,242 Mbp.
10/17: Cheilotheca (3), Monotropa (4). N. hemisphere, largely temperate, to Colombia, Malaya and Sumatra). Photo: Monotropa Habit.
4. Arbutoideae Niedenzu —— Synonymy: Arbutaceae Bromhead, Arctostaphylaceae J. Agardh
Shrub to tree (deciduous); ellagic acid +, C-8 iridoid glucosides +; anthers with paired awns, filaments strongly dilated basally; exothecium +; pollen oblate; style continuous with ovary; ovules 10>/carpel, integument ca 5 cells across; fruit berry or ±dry, drupaceous, stony/fibrous layer well developed, stones separating or not; testa cells rather thick-walled; n = 13.
6/ca 90: Arctostaphylos (60). Warm (cold) temperate, esp. S.W. North America, the Mediterranean. Map: from Meusel et al. (1978), Hultén (1962), Hultén and Fries (1986) and Kron and Luteyn (2005).[Photo - Flower (x-sec), Inflorescence.]
Age. The age of the node [Arbutus + Arctostaphylos] is (25-)15, 14(-7) Ma (Bell et al. 2010), 17-12 Ma (Wikström et al. 2001), (53.5-)40.9(-28.5) Ma (Schwery et al. 2014) or ca 28.6 Ma (?taxa: Rose et al. 2018).
[[Cassiopoideae + Ericoideae] [Harrimanelloideae [Epacridoideae + Vaccinioideae]]]: ericoid hair roots + [± monarch, = endodermis, epidermis, a tracheid, sieve tube + companion cell - 40-70 µm across], association with non-ECM-type ascomycetes and basidiomycetes, inc. Sebacinales-Serendipitaceae; (toxic andromedane diterpenes +); (vessel elements with simple perforation plates [?here]); pericyclic fibres in leaf and stem ± developed; stamens early inverting, anther wall without endothecium; pollen oblate.
Age. Wagstaff et al. (2010) date this node to ca 65 Ma, but it is estimated to be 76.9 Ma in Rose et al. (2018) and 77 Ma in Z.-Y. Liu et al. (2014).
[Cassiopoideae + Ericoideae]: leaves opposite, lamina vernation revolute.
Age. Wagstaff et al. (2010: constraint age) give the age of this node as ca 40.5 Ma, while ca 74.7 Ma is the estimate in Rose et al. (2018).
5. Cassiopoideae Kron & Judd - Cassiope D. Don
(Vessel elements with simple perforation plates - C. mertensiana); pith with large thin walled cells surrounded by smaller thick-walled and lignified cells [= Calluna-type]; pericyclic fibres in leaf and stem poorly/not developed; hairs (± fasciculate/branched); buds not perulate; stomata transversely oriented, (adaxial only); leaves small, ericoid/sublinear/peltate; flowers single, axillary; anthers with paired awns; embryo sac bisporic [Allium type] [?always]; n = 13.
1/12. Circumboreal. Map: from Meusel et al. (1978), Hultén and Fries (1986) and Kron and Luteyn (2005). [Photo - Habit.]
Age. The age of crown-group Cassiope is estimated to be 37.2 ma (Rose et al. 2018: ?sampling) and that of [[C. ericoides + C. lycopodioides] The Rest] is estimated to be ca 19 Ma (Hou et al. 2015).
6. Ericoideae Link
Flowers erect to ± horizontal; stamen appendages 0; pollen with viscin threads [= sporopollenin; threads smooth, branched or not] (0), attached distally to the grains; capsule septicidal.
19/2,096. Widespread, if scattered, not lowland tropics. Map: from Meusel et al. (1978), Hultén and Fries (1986) and Kron and Luteyn (2005).
Age. This node may be ca 67.9 Ma (Rose et al. 2018).
For some literature on fossil pollen perhaps to be associated with this group, see Kumar (1995).
6A. Phyllodoceae Drude
(Plant deciduous); (grayanotoxins + [cyclic diterpenes] +); pith heterogeneous [?Bej.]; (stomata tetracytic); leaves (opposite), (ericoid); (flowers monosymmetric by stylar curvature); pedicels articulated [not Bej.]; C (3-7, free); A (held in C pockets - Kalmia), (endothecium +, local), resorbtion tissue +; (style continuous with ovary); exotestal cells isodiametric to elongated, (exotegmen - K. procumbens); n = 11, 12.
7/43: Bejaria (15). North temperate, Bejaria south to N. Peru and the Guianas. Photo: Flower.
Age. Kalmia-type pollen is known from western Europe in deposits at least 40 Ma (Hofmann 2018).
6B. Bryantheae Gillespie & Kron
Shrubs; pith heterogeneous; leaves opposite/spiral, ericoid/flat; flowers 4-9-merous; pedicel articulated [Bryanthus]; C ± free; A = or 2X C, resorbtion tissue +; (viscin threads 0 - Bryanthus); style ± articulated with ovary; n = ?
2/8: Ledothamnus (7). Japan, East Russia (Bryanthus), the Guayana highlands (Ledothamnus).
6C. Ericeae de Jussieu —— Synonymy: Salaxidaceae J. Agardh
Shrublets to small trees (trees); buds not perulate; leaves opposite (spiral), blade ericoid (flat); inflorescence axillary, (bracts and bracteoles recaulescent/0); A (3-)8(-10), (= and opposite K), thecae dorsi- to basifixed, (spreading), (pores elongated), paired spurs at base of thecae/on filament/0, (filaments ± connate/S-shaped); pollen (grains single), viscin threads 0; G (1-)4(-8), style (0), often continuous with or impressed in ovary, (excentric), stigma punctate to peltate to large and funneliform; 1-many ovules/carpel; fruit indehiscent [drupe, etc.]/loculicidal capsule, C usu. persistent (not - Daboecia); n = 8, 12.
3/860: Erica (853). Western Europe, tropical African mountains, Madagascar, esp. South Africa. Map: see Oliver (2000: Fig. 15 - Erica). Photo: Habit, Flower.
Age. Pollen from western Europe about 56 Ma has been identified as Erica (Hofmann 2018).
[Empetreae + Rhodoreae]: n = 13.
Age. The age of this clade is around 64 Ma (Xia et al. 2021: fossil calibration point).
6D. Empetreae Horaninow —— Synonymy: Diplarchaceae Klotzsch, Empetraceae Hooker & Lindley
Shrubs; buds not perulate; leaves opposite (spiral - Diplarche), blade flat/ericoid; (plant mono-/dioecious); (pedicels articulated - Diplarche); K 3-6, C 0 (5); A 3-10, (A 5, adnate to C - Diplarche); (tapetal cells multinucleate - Empetrum); viscin threads usu. 0; G [-9], style short [shorter than the ovary], (stigma deeply lobed); (1 ovule/carpel); fruit often a drupe; testa cells isodiametric.
4/7: Empetrum (3). Scattered: N. temperate to Arctic, Azores and the Iberian Peninsula, east North America, southern Andes, Tristan da Cuhna, eastern Himalayas, southwest China. Photo: Empetrum fruit, © J. Maunder.
6E. Rhodoreae Bremekamp - Rhododendron L. —— Synonymy: Azaleaceae Vest, Rhodoraceae Ventenat
Shrubs to small trees, plant (epiphytic); grayanotoxins [cyclic diterpenes] +; nodes (1:3≤/3:5≤); hairs glandular lepidote + eglandular multiseriate(-branched); leaf blade ± flat, vernation revolute (convolute); inflorescence buds perulate; flowers pendulous to erect, monosymmetric, at least by stylar curvature (polysymmetric); K (median member abaxial), (lacking vascular traces); A (= and opposite K), collapse tissue +; seeds also tailed/tuft of hairs at both ends, exotestal cells usu. elongated; plastid transmission biparental.
1/1,025: inc. Azalea, Ledum, Menziesia, Therorhodion,Tsusiophyllum. Predominantly North Temperate, the Arctic, to NE Australia, esp. montane Himalayas to Papua.
Age. This clade is around 65 Ma (N. Shrestha et al. 2018), 56 Ma (Xia et al. 2021), (58.1-)ca 56 Ma (Mo et al. 2022) or ca 62 Ma (Y. Ma et al. 2022: Fig. S16).
Rhododendron-type pollen from western Europe is dated to about 40 Ma or more (Hofmann 2018) while fossil seeds (R. newburyanum) from the same area are ca 54.5 Ma (Collinson & Crane 1978).
6E1. Rhododendron subg. Therorhodion (Maximowicz) A. Gray
Plant (deciduous); indumentum glandular hairy; bracteoles foliaceous; C ± rotate; x = 12.
3 spp.: N.E. Asia (N.E. China, Korea, Japan, E. Siberia), W. Alaska.
[Azaleastrum [Rhododendron [Pentanthera + Hymenanthes]]]: ?
Age. This clade is about 53 Ma (Y. Ma et al. 2022: Fig. S16).
6E2. Rhododendron subg. Azaleastrum inc. subg. Tsutsusi —— Synonymy: Menziesiaceae Klotzsch
inflorescences axillary; corolla ± funneliform.
102 spp.:
[Rhododendron [Pentanthera + Hymenanthes]]: ?
Age. The age of this clade is estimated to be ca 50 Ma (Y. Ma et al. 2022: Fig. S16).
6E3. Rhododendron subg. Rhododendron —— Synonymy: Ledaceae J. F. Gmelin, Rhododendraceae de Jussieu
indumentum lepidote; A (5-)10(-18).
440 spp.: Malesia (to N.E. Australia).
[Pentanthera + Hymenanthes]]: ?
Age. This clade is perhaps ca 47.5 Ma (Y. Ma et al. 2022: Fig. S16).
6E4. Rhododendron subg. Pentanthera.
Plant deciduous; A 5.
18. Esp. E. North America, also Poland and Austria to the Caucasus, China, Japan. Sect. Rhodora has the upper three lobes joined into a single three-lipped lobe
6E5. Rhododendron subg. Hymenanthes (Blume) K. Koch
; C ± campanulate.
302 spp.: North Temperate (scattered), esp. mountains of southwest China.
Age. Diversification here began ca 35 Ma (Y. Ma et al. 2022: Fig. S16).
[Harrimanelloideae [Epacridoideae + Vaccinioideae]]: K in fruit not withering.
Age. The age of this node is about 77 Ma (Z.-W. Liu et al. 2014) or 71.7 Ma (Rose et al. 2018).
7. Harrimanelloideae Kron & Judd - Harrimanella Coville
Leaves acicular, lamina margins entire; flowers single, terminal; anthers with paired spurs; style short, continuous with ovary; n = ?
1/2. Interruptedly circumboreal. Map: from Hultén and Fries (1986) and Kron and Luteyn (2005).
Age. Crown-group Harrimanelloideae are estimated to be ca 51.4 Ma (Rose et al. 2018).
[Epacridoideae + Vaccinioideae]: (multiseriate rays, wide and high).
Age. Wagstaff et al. (2010) suggest that the age of this node is ca 37.8 Ma, but it is around 65.3 Ma in Rose et al. (2018) and 67 Ma in Z.-Y. Liu et al. (2014).
8. Epacridoideae Arnott (Styphelioideae)
Axial parenchyma usu. diffuse (in aggregates); rays exclusively uniseriate; lamina epidermis ± lignified, cells ± rectangular, in longitudinally parallel  ranks, ad-/abaxial epidermal cells sinuous anticlinally, stomata parallel to leaf axis, ± circular, epicuticular waxws amorphous and/or as platelets; vascular bundles embedded, with well-developed abaxial fibrous tissue, no adaxial cap; leaves xeromorphic, pungent, >3 veins from base, midrib evident; inflorescences often axillary, usu. spikes/axillary flowers, flowers often lacking a clear pedicel, bracteoles several; flowers often pendulous; A = C, opposite K, epipetalous, anthers bisporangiate, monothecal, exothecium +, dehiscing by slits, appendages 0; C persistent in fruit.
ranks, ad-/abaxial epidermal cells sinuous anticlinally, stomata parallel to leaf axis, ± circular, epicuticular waxws amorphous and/or as platelets; vascular bundles embedded, with well-developed abaxial fibrous tissue, no adaxial cap; leaves xeromorphic, pungent, >3 veins from base, midrib evident; inflorescences often axillary, usu. spikes/axillary flowers, flowers often lacking a clear pedicel, bracteoles several; flowers often pendulous; A = C, opposite K, epipetalous, anthers bisporangiate, monothecal, exothecium +, dehiscing by slits, appendages 0; C persistent in fruit.
35/545. Australasia, Chile, Hawaii. Map: see Sleumer (1964), Kron and Luteyn (2005), FloraBase (2006) and Australia's Virtual Herbarium (consulted xi.2012). Photo: Flower, Fruit & Flowers.
Age. Crown-group Epacridoideae are 89.3-68.7 (Schwery et al. 2014) or ca 65.3 (Rose et al. 2018) Ma.
8A. Prionoteae Drude —— Synonymy: Prionotaceae Hutchinson
Stomata paracytic; lamina with three veins from the base; flowers solitary; anthers dehiscing by two slits; n = ?
2/2. Chile, Tasmania.
[Archerieae [Oligarrheneae [Cosmelieae [Richeeae [Epacrideae + Styphelieae]]]]]: multicellular hairs 0; stomata anomocytic; lamina with parallel veins, margin lacking serrations; anthers dehiscing by a single slit; (pollen tetrads not fully fertile).
8B. Archerieae Crayn & Quinn - Archeria J. D. Hooker
Rays also biseriate; abaxial epidermis plus hypodermis detached from mesophyll; inflorescence terminal, flowers axillary, bracteoles 2; n = ?
1/4: Australia (Tasmania), New Zealand.
[Oligarrheneae [Cosmelieae [Richeeae [Epacrideae + Styphelieae]]]]: main veins ± equal.
8C. Oligarrheneae Crayn & Quinn
Abaxial surface of lamina lacking ribbon wax and papillae; flowers 4-5-merous; A 4, 2; C glabrous, lobes valvate to induplicate-valvate; (A 2-5); G [2], style continuous with ovary, short [shorter than ovary]; ovule 1/carpel, apical; fruit a nut; n = ?
3/7: Dielsiodoxa (5). Western Australia.
[Cosmelieae [Richeeae [Epacrideae + Styphelieae]]]: ?
8D. Cosmelieae Crayn & Quinn
Vessels up to 500 mm2; axial parenchyma scanty paratracheal; "adaxial leaf stomata tetracytic"; leaves sessile, bases sheathing, not leaving a scar; (anthers dithecal?), anthers strongly curved; n = 12, 13.
3/27: Andersonia (22). Southwest and east Australia, Tasmania.
[Richeeae [Epacrideae + Styphelieae]]: ?
Age. Wagstaff et al. (2010) date this node to (37-)34.3, 33.4(-26.9) Ma, (29.8-)22.3(-15.4) Ma is the age in Schwery et al. (2014) and ca 54.6 Ma in Rose et al. (2018).
8E. Richeeae Crayn & Quinn
Vessels up to 500 mm2; axial parenchyma scanty paratracheal; crystals in ray cells only; rays also ≥20-seriate; nodes 3:3-several; lamina bundles transcurrent ad- and abaxially by fibrous bundle sheath extensions, stomata brachyparacytic, (elongated [length:width ratio ≥1.2]), (epicuticular waxes also as tubes); leaves sessile, often large [to ≥15 cm (-1.5 m) long], bases sheathing, leaving a scar [cortex also exfoliates]; bracteoles 2-6; (C connate, operculate, circumscissile above base); (A adnate to C); nectary of separate scales; n = 13.
2/68: Dracophyllum (63). East (central) Australia, New Zealand, New Caledonia.
Age. Wagstaff et al. (2010) date crown Richeeae to (23.5-)20.6, 16.5(-8.7) Ma.
Fossils from New Zealand described as Richaeaphyllum waimumuensis are perhaps ca 23 Ma, but these are unlikely to have anything to do with the Dracophyllum currently on the islands (Jordan et al. 2010).
8F. Epacrideae Dumortier —— Synonymy: Epacridaceae R. Brown, nom. cons.
Rays usu. also to 10-seriate; (stomata transversely oriented - Lysinema); (midrib ± prominent); (ovules apotropous - L.); n = 12, 13.
5/55: Epacris (45). East Australia, Tasmania.
8G. Styphelieae Bartling —— Synonymy: Stypheliaceae Horaninow
Crystals in axial parenchyma only (0); rays to 20-seriate; lamina bundles adjacent to abaxial epidermis/one layer of lignified cells; ad-/abaxial epidermal cells often not sinuous anticlinally, stomata elongated [length:width ratio ≥1.2], often papillae over stomata, (elsewhere on leaf surface), (epicuticular waxes as ribbons); bud scales 0 [?level]; leaves (opposite); (plant dioecious); inflorescence with several bracts, terminated in rudimentary flower; flowers sessile, bracteoles +; C valvate, (lobes densely hairy); pollen not all members of tetrads fertile/(grains single, three cells of the meiotic quartet not developing), [pseudomonads], (4-pantoporate); G loculi 1-11, style continuous with ovary; one ovule/carpel, apical; fruit a drupe; n = 4, 6-14.
Ca 16/384: Styphelia (146, + 74 undescribed), Leucopogon (80). Australia to Malesia and Southeast Asia, Fiji, New Caledonia, Hawaii.
Age. Cyathodophyllum novae-zelandiae, leaf fossils from New Zealand some ca 23 Ma, are probably closest to the Tasmanian Cyathodes (Jordan et al. 2010).
9. Vaccinioideae Arnott
(Plant deciduous); axial parenchyma scanty, paratracheal; (stomata paracytic); apical bud aborting; (lamina entire), (with marginal or surficial glands); inflorescences axillary; pedicel articulated, bracteoles +; anthers with dorsal disintegration tissue; stigma truncate; x = 12.
Age. The age of crown-group Vaccinioideae is estimated to be ca 46.2 Ma (Rose et al. 2018).
50/1,615. N. hemisphere, Malesia and montane Central and South America, Australia (Queensland), few in Africa. Map: from Meusel et al. (1978), Hultén and Fries (1986), Fl. China vol. 14 (2005), Kron and Luteyn (2005) and Australia's Virtual Herbarium (consulted i.2014).
Age. The age of [Oxydendrum + the rest] is estimated to be ca 46.3 Ma and that of [Lyonia + the rest] some ca 44 Ma (Rose et al. 2018).
9A. Lyonieae Kron & Judd
(Plant deciduous); (grayanotoxins [cyclic diterpenes] +); (lamina vernation revolute); secondary phloem stratified; lamina epidermis ± lignified; (anthers with paired spurs); filaments S-shaped (straight).
4/82: Agarista (39), Lyonia (35). Scattered: temperate E. Asia, N. America, the Caribbean, South America, esp. S.E. Brazil, and Africa, esp. Madagascar.
9B. Oxydendreae Reveal - Oxydendrum arboreum (L.) de Candolle
Deciduous tree; hairs eglandular; stomata paracytic; inflorescence terminal, branched, initiation to fruit <1 year; pedicel not articulated; traces to floral organs all leave floral axis separately; stamen appendages 0; testa cells elongated; n = 12.
1/1. Growing in a southwest-northeast band in the Southeastern U.S.A, barely in Florida.
[Gaultherieae, Andromedeae, Vaccinieae]: ?
Age. The age of this clade has been estimated to be ca 32.4 Ma (Rose et al. 2018: Gau.Vac).
9C. Gaultherieae Niedenzu - Gaultheria L.
Small to largish shrubs; (grayanotoxins [cyclic diterpenes], methyl salicylate +); indumentum (lepidote); stomata paracytic; inflorescence terminal, (bracts foliaceous); anthers (shortly tubular), (disintegration tissue 0), awns four/0; integument 4-11 cells across; K persistent, fruit (a berry)/(K accrescent, fleshy); n = 11 (12, 13).
1/292. Esp. China to Malesia, also northern Hemisphere, northeast South America, southern Andes, southeast Australia, New Zealand. Photo: Gaultheria flowers.
Age. Crown-group Gaultherieae are around 13.9 Ma (Rose et al. 2018) or ca 18 Ma (Lu et al. 2019a).
Pollen from western Europe about 56 Ma has been identified as that of Gaultheria (Hofmann 2018).
9D. Andromedeae Klotzsch —— Synonymy: Andromedaceae Döll
Shrub; multicellular hairs 0 [on lamina surface]; lamina (vernation revolute), (entire); anther with two or four awns, (disintegration tissue 0 - Andromeda); integument ca 10 cells across [Andromeda]; (n = 11).
2/2. North ± cold temperate to boreal, southeast United States.
9E. Vaccinieae Reichenbach —— Synonymy: Oxycoccaceae A. Kerner, Vacciniaceae Perleb, nom. cons.
Shrubs (small trees), epiphytic [ca 1/4 spp]), (stem base swollen); root with (hyphal sheath and Hartig net +, hair roots short); (stem cork cambium subepidermal); stomata paracytic; lamina (plinerved), (with basal marginal glands); anthers with tubules (not), disintegration tissue usu. 0, appendages spurs/2 or 4 awns/0; G inferior (superior), (pseudo 10-/10-locular); ovules 1-many/loculus; fruit baccate (drupaceous - Gaylussacia); (testa mucilaginous); (embryo chlorophyllous).
30-35/1,563 (Schwery et al. 2014): "Vaccinium" (?785 - in Southeast Asia-Malesia ca 400, but to include Agapetes s. str. (110), Rigiolepis (26)), Dimorphanthera (ca 85), in tropical America Cavendishia (155), Psammisia (60), Thibaudia (60), Macleania (55), Gaylussacia (50). Esp. montane Southeast Asia to Malesia, Central and South America, also southeast Brazil, also general Northern Hemisphere, few Australia (Queensland) and Africa. Photos: Psammisia flowers, Vaccinium flowers, © J. Maunder.
Age. The age of crown-group Vaccinieae may be (35.6-)29.5(-21.5) Ma (Becker et al. 2024), ca 21.3 Ma (Rose et al. 2018: Vaccinium arboreum, Cavendishia, Utleya); see also Diaz-Garcia et al. (2021) for some ages within Vaccinium - sect. Cyanococcus-V. macrocarpum, actually, most of the tribe - a mere 10.4 Ma or so.
Pollen from western Europe about 56 Ma has been identified as that of Vaccinium (Hofmann 2018).
Floral formula: * ⚥ K [5]; C [5]; A 10; N; G [5].
Evolution: Divergence & Distribution. Ericaceous pollen has been identified in heathland vegetation dated to 75-65.5 Ma in Central Australia (Carpenter et al. 2015), or perhaps a little earlier from the Otway Basin, but the form species Ericipites scabratus into which much ericaceous pollen has been placed is difficult to identify much more accurately and may come from Ericoideae, Epacridoideae or even Clethraceae (Dettmann & Jarzen 1990; Jordan & Hill 1995). For an evaluation of the fossil record of Epacridoideae, much from Tasmania and southeast Australia, see Jordan and Hill (1995, 1996). Pollen from Western Europe ca 56 Ma has been identified as Ericoideae and placed in three genera, and perhaps somewhat younger pollen as Vaccinioideae and placed in two genera (Hofmann 2018); note that the ages here are often very different - far older - than some other estimates. There is Late Cretaceous ericaceous pollen from southern Africa (Scholz 1985: Namaqualand), Palaeocene and later pollen from India (Kumar 1995), while pollen with viscin threads, quite possibly from Rhododendron, is known from several localities in the northern hemisphere in deposits as old as ca 45 Ma (see Zetter & Hess 1996). Quite well preserved seeds of Rhododendron (R. newburyanum) were described from Palaeocene deposits in southern England (Collinson & Crane 1978), however, Xia et al. (2021) suggest that the genus soon became extinct there. Maiella miocaenica, from 16.3-12.8 Ma deposits in Poland, is similar to Calluna in many respects, but not in seed micromorphology, and although it was thought to be the oldest fossil record for Ericeae (Kowalski & Fagúndez 2017), pollen of other genera appears to be older (Hofmann 2018). There are lots of questions here! Indeed, Comarostaphylis globula (Arbutoideae), of about the same age and also from Poland, is probably not ericaceous at all (Kowalski & Worobiec 2018).
Kron and Luteyn (2005) discuss the historical biogeography of Ericaceae; an Eurasian origin of the family is likely. They give useful distribution maps for the subfamilies, Cassiopoideae being shown as growing throughout Greenland, perhaps in anticipation of the disappearance of the ice. However, the biogeographical connections of some of the newly proposed relationships are not easy to understand, for example, Gillespie and Kron (2010) found that the Guayanan Ledothamnus was sister to the northeast Asian Bryanthus. A number of ages for major generic groupings in the family can be estimated from Fig 1 in Z.-W. Liu et al. (2014); some are given above.
One way of thinking about diversification in the family is to focus on clades in montane habitats, often shrubby and with low SLA (specific leaf area = relatively high mass/surface area, see below). Bouchenak-Khelladi et al. (2015) thought that being shrubby and with a low SLA, features that evolved at/below the base of the family, were essential pre-requisites for the later radiations within it; here there was movement into montane environments where taxa in general tend to have a low SLA, and this might sometimes trigger diversification. Schwery et al. (2014) also noted that the association of Ericaceae with distinctive mycorrhizal types and oligotrophic, acidic conditions (see below) might also be contributing factors to these montane radiations. More specifically, there are six major radiations in the family, including Epacridoideae-Richeeae, Ericoideae-Rhodoreae and -Ericeae, and Vaccinioideae-Vaccinieae (both Gaultheria and the inferior-ovaried taxa on both sides of the Pacific), including in total almost 80% of the family, in which there has been considerable diversification in mostly temperate or montane habitats (Schwery et al. 2014; Bouchenak-Khelladi et al. 2015; Hughes & Atchison 2015). See also Ecology & Physiology: Group 4 (below) for some discussion of the epiphytic habit here.
- L. Lu et al. (2019a) looked at the biogeography and diversification of Gaultheria s.l., and found (i.a.) that species with red fruits were more likely to disperse long distances, while diversification was associated with species that had violet fruits, and there were also correlations of fruit colour with both latitude and altitude.
- The inclusion of the largely Australian Epacridaceae within Ericaceae as Epacridoideae means that the rarity of Ericaceae in Australia no longer presents a evolutionary/biogeographical problem as it did in the 1970s when I was writing a thesis on what were then Ericaceae - Ericaceae as a clade are indeed quite common there. Diverse early Pleistocene fossils of Epacridoideae are known from New Zealand (Jordan et al. 2007), but there are serious conflicts between molecular and fossil estimates of clade ages. Thus leaves and pollen from New Zealand and identified as Richeeae are ca 25-20 and 47-40 Ma old respectively, while in a molecular study Wagstaff et al. (2010) date the stem age of the clade of Dracophyllum that now grows in New Zealand at a mere (7.2-)6.2(-5.2) Ma. Indeed, Jordan et al. (2010) thought that many of the fossil records in New Zealand might belong to extinct lineages, and this idea was seconded by Puente-Lelièvre et al. (2012) who suggested that Styphelieae would have to be some 210-120 Ma if these fossils were assumed to be members of the clades currently growing on the islands (see also Wagstaff et al. 2010). Puente-Lelièvre et al. (2012) found that seven of the eight New Zealand species of Epacridoideae that they examined all had their closest relatives in Australia (that of the other species was in New Guinea) and had arrived in New Zealand within the last 4 Ma or so.
- Within Ericoideae the numerous African species of Erica form a clade that originated within that part of the Erica tree that otherwise includes a few other taxa currently found in Europe (Pirie et al. 2011, 2024; see also McGuire & Kron 2005). Erica, at around 659 species in the Cape Floristic Region alone and as many as 860 species in South Africa (estimates vary) is the most diverse genus in the hyperdiverse Cape Floristic Region, especially in the Fynbos, but not very common in the Renosterveld (Linder 2003, see also Pirie et al. 2017; du Preez et al. 2025b); probably monophyletic there, Cape diversification has been dated to within the last 15 Ma or so, and the species are often closely related - the few European taxa in fact encompass a disproportional amount of phylogenetic diversity (Pirie et al. 2016, 2024). There are some studies on how pollinators might be implicated in the generation/maintenance of this diversity (S. D. Johnson 2010; Bouman et al. 2017; van der Niet et al. 2020). 71-64 Ma pollen identified as Ericaceae has been found in Namaqualand, South Africa (Linder 2003); knowing what plant produced it would be of more than passing intereat. Ericaceous pollen appears ca 9 Ma in ocean sediments off Southwest Africa, and for changing climates then in the context of changing ocean currents, see Dupont et al. (2011).
- Rhododendron, which grows throughout much of the northern hemisphere, is notably diverse in much of the Malesian-South East Asian area. There are around 225 species in the Hengduan region of China alone (Boufford 2014), and two increases of diversification in the genus there are dated to 15-9 Ma, perhaps at the beginning of the uplift of the Hengduan mountains - see subgenera Rhododendron and Hymenanthes (Xing & Ree 2017; Y. Ma et al. 2022: Fig. S16). Mo et al. (2022) suggest that diversification in the genus increased around 14 Ma and especially 5 Ma and since, in both cases times when global temperatures were falling. Lack of seasonality (N. Shrestha et al. 2017) and environmental heterogeneity may play roles in diversification in Rhododendron (see also orchids), which may have started in NE Asia, the genus then moving around the northern hemisphere and latterly into the Southeast Asian tropics; there the species are young and are nearly always to be found in montane habitats (N. Shrestha et al. 2018; but c.f. Khan et al. 2021, if only in part). Similarly, Xia et al. (2021) suggested that Rhododendron was originally from northeast Asia, became circumboreal, but subsequent diversification rates were particularly high in eastern Asia, especially Malesia (sect. Schistanthe), partly because of the habitat heterogeneity (considerable elevational range) and high precipitation there. Species from higher northern latitudes tend to be basal in the groups to which they belong. Two clades were recovered in subgenus Hymenanthes, basal in one were species from southwest China and surrounds and from Japan and southwestern Eurasia, while in the other were species from North America, southwestern Eurasia and northeastern Asia (Ma et al. 2022). Although subgenus Therorhodion (two extant species) diverged from the rest of the genus in the Palaeocene, it was around 20 or 32 Ma (see Xia et al. 2021: Figs S4, S5 for dates; Mo et al. 2022 respectively) before diversification in the genus began in earnest in the late Eocene (Xia et al. 2021) or late Oligocene (Mo et al. 2022). Species are predominantly diploid in subgenus Hymenanthes, but there seems to have been extensive hybridisation that has been involved in its diversification - there are ca 302 species here... (Ma et al. 2022). There are around 290 species of Rhododendron in Malesia (Sleumer 1966), nearly all members of sect. Schistanthe (subgenus Rhododendron, the old Vireya group), a clade 21.8 Ma (Xia et al. 2021) or ca 31 Ma (Soza et al. 2022). These rhododendrons seem to have moved through the archipelago west to east, basal members of the clade being from southeast Asia, while in New Guinea un particular is to be found a speciose clade of subsection Euvireya (Goetsch et al. 2011; see also Xia et al. 2021; G. Khan et al. 2024) - all told, there are some 170 or so species on the island, diversification in the major Euvireya clade restricted to the island beginning ca 6.3 Ma (Soza et al. 2022); there are also a few taxa from places south and east of the island. There is bird pollination, perhaps especially at higher altitudes (Stevens 1976), and Soza et al. (2022) looked at the considerable floral variation of sect. Schistanthe in the context of its phylogeny - they were unclear if its ancestor had polysymmetric or monosymmetric flowers, it seems to depend on the axes of the PCA considered. Khan et al. (2021) linked diversification here to genome downsizing after polyploidy (1 Cx = 0.483-0.618 pg, vs up to 2.777 pg elsewhere in the family) - small genomes ≡ fast diversification, as in other clades. For diversification in Rhododendron, see also J. B. Thompson et al. (2023); Y. Shen et al. (2024) looked at the phylogenetic distribution of a few characters.
- Vaccinioideae-Vaccinieae include taxa from low altitudes in boreal and temperate habitats to high altitudes in South America and Malesia; they are particularly diverse in the mountains of Central and South America and of Southeast Asia-Malesia. As is clear below, many of these species are epiphytic and/or lianes, some have distinctive sticky seeds probably associated with the epiphytic habitat, and bird pollination is likely important in both the Old and New Worlds. However, depressingly little is known about ecology and diversification here. The hyperspeciose clade in Vaccinieae - it includes "Vaccinium", Agapetes s. str., Cavendishia, etc., i.e., most of the tribe - has been dated to (54.4-)45.6(-37.2) Ma (Schwery et al. 2014), although given the relationships of "Vaccinium" to other Vaccinieae, a major effort is needed to put variation and evolution in the tribe as a whole in a phylogenetic context. Kron and Luteyn (2005) suggested that there were perhaps two migrations from North to South America, and they dated diversification in South American Vaccinieae to the Late Miocene. Vaccinieae from the Neotropics and North America may have had similar mycorrhizal Sebacinales-Serendipitaceae in common, and some kind of synchronised island hopping of plant and fungus between the two continents may have been needed (Setaro & Kron 2011; see also below). Although it has been suggested that connections between North and South America were established around the middle of the Miocene ca 24 Ma (see Bacon et al. 2015 and the discussion of that paper; Montes et al. 2015: geology re-evaluated), a recent comprehensive refutation of this idea should be consulted (see O'Dea et al. 2016). Outlines of interesting biogeographical groupings in Neotropical Vaccinieae in particular are developing (Kron et al. 2002a; see also Powell & Kron 2003; Pedraza-Peñalosa 2009), and these may be correlated with variation in wood anatomy (Lens et al. 2004c). Becker et al. (2024) have recently discussed the evolution and dispersal of Vaccinieae, and their paper should be consulted for details - i.a. they look at the evolution of a number of characters such as fruit colour. They suggest that the shift in ovary morphology 5-locular→pseudo 10-locular happened only once, the taxa involved including the North American Vaccinium sect. and the South East Asian-Malesian Vaccinium/Agapetes complex (Becker et al. 2024). It has been clear for some time that little separates the 100-plus species of Agapetes s. str. from the even more numerous species of Vaccinium from the southeast Asian area (e.g. Stevens 1985)- see also Phylogeny below. Becker et al. (2025) noted that Agapetes s. str. and associated species of Vaccinium represent a major movement of the Vaccinieae into warm-temperate/tropical regions, another being represented by the Malesian Paphia and Dimorphanthera and the numerous New World Vaccinieae centred on the mountains of Central and South America. Atkinson et al. (1994) had early suggested possible correlations between polyploidy, geography, and anatomy in tropical Vaccinieae. These authors also suggest that Vaccinieae are of North American origin and that there have been some seven movements into the tropics and four main diversification rate increases. Polyploidy is prevalent in taxa in the East Malesian—Andean part of their tree and is associated with one of these rate shifts (Becker et al. 2024).
Intersectional hybrids in Vaccinium are well known (e.g. Fritsch et al. 2020a: sections Myrtillus X Pyxothamnus, California), but the situation in Hawai'i and surrounds is more complicated, as Becker et al. (2025) found out. The 3-18 species (species concepts!) of Vaccinium on Hawai'i are montane plants and an important source of food for birds, and they are early colonizers of places like new lava flows - rather extreme habitats. Becker et al. (2025) found that these species belonged to section Myrtillus, and they are sister to V. yatabei, from Japan. Temperate East Asia is a rather uncommon point of origin for Hawai'an plants; Vaccinium probably moved from there to Kaua'i 8.4/6.2-5.1/3.7 Ma. A wrinkle or two: V. cereum, from Ua Pou (the Marquesas islands), consistently groups within these Hawaii'an taxa, while three other specimens of V. cereum, but from three other islands in French Polynesia, seem to be hybrids between sections Bracteata and Myrtillus. The morphology of the Oa Poa Vaccinium is poorly known, but it seems to be closer to that of sect Myrtillus than that of the other Marquesan specimens (Becker et al. 2025). Finally, a specimen of V. ovalifolium (section Myrtillus) from west North America was found to be nested in the Hawai'ian clade in chloroplast analyses, and possibly represents a hybridization event between a specimen of the Hawai'ian clade that dispersed to North America with the local V. ovalifolium (Becker et al. 2025). Even if this is not confirmed, there seems to have been extensive and quite recent long-distance dispersal in this part of Vaccinieae. All in all, Vaccinieae are a large, diverse and ecologically important group our understanding of which is hamstrung by our lack of knowledge of relationships from the species level on up and by unclear boundaries between some of the species themselves.
Turning now to Arbutoideae, prominent in Mediterranean vegetation although hardly very diverse, members of this clade may have moved from the New to the Old Worlds around 39.2-21.2 Ma (Hileman et al. 2001; Vargas et al. 2014), although Mediterranean vegetation is thought to be well under half that age (Vargas et al. 2014). Parker and Stickrod (2022) examined changes in phenology and morphology in New Word Arbutoideae associated with changing climate in the Tertiary, although dating is clearly an issue (ibid., see also above). Hardy and Cook (2012) compared diversification in Monotropoideae (it has slowed) with that of Arbutoideae (exponential increase). For the circumboreal species of Pyrola, a genus perhaps Asian in origin, and their places of origin, see Z.-W. Liu et al. (2014). Cassiope, also hardly diverse in terms of numbers of species (but for leaf morphology, see below), seems to have had an origin in the north (Hou et al. 2015), unlike other arctic-alpine genera (Hagen et al. 2019). Empetrum is likely to have arrived in southern South America in the Pleistocene, perhaps by long-distance dispersal from northwestern North America (Popp et al. 2011).
Heads (2003) perhaps rather improbably suggested that the main elements of the distribution patterns in the family were best explained by vicariance. The diverse Malesian Ericaceae in particular were, he thought, largely derived from taxa that lived in the mangroves (for him, the ancestral habitat of angiosperms as a whole), their current prevalance in higher-altitude vegetation being the result of rapid tectonic uplift, the plants hanging on as the mountains rose. However, in this context Ericaceae are hardly noted for being a megatherm family and why they should not be better represented in mangroves (and other tropical lowland habitats) than they are now is unclear.
Hybridization seems to have been important in the evolution of Gaultherieae in particular (Lu et al. 2019b and references), at least some groups of Rhododendron section Schistanthe (Soza et al. 2022), perhaps also in the small but vegetatively very variable Cassiope (Gillespie & Kron 2013: 3/13 species examined involved), and so on. Back in the late 1930s there was some excitement when an intergeneric cross between Gaultheria and Pernettya, Gaulnettya wisleyensis or Gaulnettya x Wisley Pearl, appeared at Wisley Gardens where the putative parents, G. shallon and P. mucronata, were both being grown (Mulligan 1939); alack, the two species are now both in Gaultheria. Furthermore, given that it is proving possible to hybridize taxa like Vaccinium bracteatum (immediately related to Agapetes, etc.) and the Andean V. meridonale with the core species of Vaccinium(Edger et al. 2022), and that limits of many of the genera recognized in Vaccinieae (as of iv.2025) seem to have little to do with relationships, it is going to be of general interest to see what the limits to/patterns of hybridization are in this area.
Lens et al. (2003, 2004a, b, c) place variation in wood anatomy in a phylogenetic context. I have not attempted to put this variation on the tree, but there is extensive homoplasy in most wood characters within a subfamily. For wood and its evolution in the family, see also Dulin and Kirchoff (2010). For a phenetic analysis of some staminal characters, see Vander Kloet and Avery (2007). For apomorphies of the various clades, see Kron et al. (2002b) in part; data in Jordan et al. (2010) have been used to suggest some vegetative/anatomical apomorphies in Styphelioideae. Anderberg (1992, 1993) discussed the evolution of a number of chemical features, and this work needs to be integrated into current ideas of relationships. Sorokin et al. (2023) looked at the anatomy of the capsule of Enkianthus which, they thought, was the plesiomorphic fruit type for the family as a whole (see also Sorokin et al. 2025 - similar fruits are also common in Clethra); Cyrillaceae have indehiscent fruits. Indeed, Yatsenko et al. (2025) suggested that the berry of Arbutus and drupe-type fruit of Arctostaphylos independantly evolved from such Enkianthus-type capsules.
Kriebel et al. (2023: to be consulted for details) also looked at at macroevolutionary shifts in Ericaceae that could be associated with changes in four floral features. Thus they found 17 or 11 (depending on the method of analysis) such shifts in corolla shape, three of which were in common between the two analyses: Shifts to more linear corollas in neotropical Vaccinieae, Styphelioideae and within Rhododendron sect. Schistanthe. Shifts in corolla tube length consistently picked up the evolution of separate petals in Pyroloideae and Bejaria. Interestingly, mean values of floral traits were quite similar when comparing continents, and they suggested phylogenetic conservatism was involved, for instance, both Africa and Europe had many species of Erica (there are in fact only 20 species of the latter genus in Europe), and a similar situation in Rhododendron in North America (again, not that many species) and Asia; overall, Kriebel et al. (2023) thought that there there had been substantial parallel evolution in floral form. Diversification in Rhododendron sect. Schistanthe depended in part on the evolution of relatively long and narrow corollas, although rate shifts were local (context: Ericaceae) and postdated the evolution of honeyeater pollination - c.f. hummingbirds and New World Vaccinieae, where there were similar corolla shifts, but diversification rate shifts were in the context of Ericales as a whole (Kriebel et al. 2023). Y. Yu et al. (2019a) examined the evolution of pollen morphology-disparity which, although not changing at a constant rate over time, appeared not to be linked with species diversity. Puente-Lelièvre (2015) noted that pseudomonads seem to have evolved more than once in the family, and they are probably not an apomorphy for Styphelieae (see also Furness 2009). Freudenstein and Broe (2024) found a fair amount of parallelism/convergence in morphological features within Monotropoideae in part because of the position of the [Allotropa + Hemitomes] clade; being holomycoheterotrophic, morphology and anatomy in the subfamily tend to be bizarre.
Ecology & Physiology. Ericaceae are prominent components of vegetation growing in often rather acidic, coolish and well-lit habitats; they are at best uncommon in lowland tropical rainforest, grasslands and on alkaline soils. The majority of the family is associated with ericoid mycorrhizae (EM), the hyphae growing in and surrounding the ultimate branches of their roots, and this EM association is likely to be an important element in the ecological success of Ericaceae as a whole; other mycorrhizal types are also known from the family, particularly the more basal clades. In terms of general functional distinctiveness, Ericaceae are in a clade with distinctive mycorrhizae that has notably low leaf nitrogen and a notably high leaf mass per area, i.e. a low specific leaf area - SLA (Cornwell et al. 2014).
The map here shows some of the areas with shrubby vegetation in which Ericaceae dominate. Note that it differs considerably from that showing the distribution of EM (see below) in Soudzilovskaia et al. (2017) where EM records are absent from nearly all Africa, India, Asia and Malesia, but that was based on GBIF records, and the authors noted other problems with distributions. Ericoideae-Ericeae and Epacridoideae respectively are common shrubs in the Mediterranean heathlands of southern Africa and southwest Australia, thus there are 600 or more species of Erica growing in the Cape region of South Africa alone, in ericoid thickets at high altitudes (2000 m<) in Madagascar, and so on (Oliver 2000; Hackel et al. 2025). In California Arctoctaphylos and relatives are a conspicuous component of the vegetation (Parker & Stickrod 2022). (Rundel et al. 2016 discuss Mediterranean biomes in general). Most Ericaceae are shrubby plants, and Ericaceae (Ericoideae-Rhodoreae, Vaccinioideae-Vaccinieae) may dominate in montane shrubberies, especially in the northern Andes, parts of the eastern Himalayan-Yunnan region and south into Malesia, and are a prominent feature of alpine and arctic tundra and boreal forests (e.g. Chapin & Körner 1995; Jonasson & Michelsen 1996; Michelsen et al. 1998; Sistla et al. 2013) - and of course, because a plant is shrubby does not necessarily mean that it is short-lived. From the literature review in Centenaro et al. (2023: Appendix S1) the oldest clonal plants - indeed, the oldest seed plants known, period - known were Gaylussacia spp. (= "clonal herb") at ca 13,000 years, and some other Ericaceae were also notably longlived. However, clones of Lomatia tasmanica (Proteaceae) are estimated to be some 43,000 years old (DeWoody et al. 2008) and those of Posidonia oceanica (Posidoniaceae) in the Mediterranean may be up to 15 km across and thousands to 200,000 years old (Arnaud-Haon et al. 2012).
Rhododendron ponticum can be an invasive producing chemicals that may be allelopathic, and it also has other effects on the environment (Erfmeier & Bruelheide 2010; see also Hüftlein et al. 2024).
There are six (partly overlapping) main ecological groupings in Ericaceae, albeit rather different in size: 1, Taxa with fleshy fruits of one sort or another, ca 1,500 species; 2, taxa with xeromorphic leaves living in more or less Mediterranean or dry habitats, 1,300+ species; 3, taxa of the arctic tundra and boreal forests, ca 5 species; 4, taxa that are epiphytic (or epilithic) and/or climbers, ca 650 species; 5, taxa with ericoid mycorrhizae, ERM, most of the family; and 6, taxa that are holomycoheterotrophic, ca 16 species. And partly cross-cutting these groups are taxa that are major nectar providers, and this is touched on briefly after this section.
Group 1. Taxa with Fleshy Fruits are discussed avove under Diversity & Distribution and also Seed Dispersal below.
Group 2. Mediterranean climates. Ericaceae growing in such climates include about 488-675 species of Erica (Ericoideae-Ericeae) in the Cape fynbos vegetation and 120 species of Epacridoideae in West Australian kwongan vegetation (see Cowling et al. 1990). Ericaceae in Mediterranean climates in particular (Ericoideae in the South African Cape Region, Epacridoideae in Australia; Arctostaphylos and relatives in the Californian chaparral), have xeromorphic leaves. Some species form starch-rich lignotubers with buds that allow the plants to resprout after fires, while others, like about three quarters of the species of Arctostaphylos, regenerate by seeding (Wells 1969; Parker & Stickrod 2022); germination is enhanced by heat and/or smoke (Bell & Ojeda 1999; Cairney & Ashford 2002; Rundel et al. 2016 - c.f. Restionaceae).
Group 3. Alpine and Arctic Regions. Ericaceae are often common in alpine and arctic tundra (e.g. Jonasson & Michelsen 1996; Michelsen et al. 1998). Tundra alone, the main component of heathland sensu Specht (1979a, b; map: see end-papers in Specht 1979a; White et al. 2000), occupies ca 8% of the earth's surface (Kranabetter & MacKenzie 2010; Gardes & Dahlberg 1996) and Vaccinium and Empetrum, both with fleshy fruits, are two of the seven major biomass accumulators there (Chapin & Körner 1995). Boreal forests occupy ca 17% of the land surface of the earth (Lindahl et al. 2002), and there the trees (Pinaceae, some Salicaceae and Betulaceae) are all ectomycorrhizal (ECM) while EM Ericaceae often dominate in the understory (e.g. Villareal et al. 2004; Vrålstad et al. 2002; Vrålstad 2004; Kranabetter & MacKenzie 2010) - see also Group 5 below. However, in many of these habitats mosses like Sphagnum are very common, and they may dominate in overall carbon accumulation (e.g. Ragoebarsing et al. 2005; Flanagan 2014).
Empetrum may dominate in mid- and sub-arctic heaths, a dominance apparently facilitated by the allelopthic suppression of potential competitors. The dihydrostilbene batasin-III (3,3'-dihydroxy-5-methoxybibenzyl) is the compound involved, and its suppresses germination and seedling growth of both trees and graminoids (Lorion & Small 2021 and references).
Group 4. The Epiphytic habit. Many species of Vaccinieae are epiphytes, and they are an important component of the woody epiphytic flora in both the Indo-Malesian and the Andean regions, indeed, Ericaceae are one of the more important epiphytic families among broad-leaved angiosperms, along with Gesneriaceae, Piperaceae and Melastomataceae (Benzing 1990; Zotz 2013). For ericaceous epiphytes, see Zotz, Weigelt et al. (2021: list), while Wagner et al. (2021), Hietz et al. (2021) and Zotz et al. (2021) discuss the ecophysiological characteristics of epiphytes - however, little is known about such features in woody epiphytes in general. Woody epiphytes (here I include the few epilithic taxa in taxa mentioned) are commonest in the fleshy-fruited tropical Vaccinieae, especially prominent in the New World, while in Southeast Asia-Malesia these include taxa currently placed in Rigiolepis and Agapetes s. str.. A number of Ericaceae are scrambling lianes, especially in the New World, where there are 300 epiphytic/scandent species (Gentry 1991; see also Sperotto et al. 2023), and lianes are quite common in South East Asian/Malesian Vaccinioideae such as Vaccinium s.l. (including Agapetes), Paphia and Dimorphanthera. Lignotubers are known from some epiphytic taxa; they lack buds and may be involved in water storage (Evans & Vander Kloet 2010).
The leaves of the largely Malesian Rhododendron sect. Schistanthe (the old Vireya rhododendrons), many species of which prefer montane habits and/or are epiphytic, are covered in scales, and these are thought to be involved in water conservation, especially at the middle part of the altitudinal range of the section (Nilsen et al. 2014). Species of sect. Schistanthe alone have idioblasts, large cells particularly prominent in the adaxial but also abaxial hypodermes. These idioblasts are notably prominent in epiphytic species and are probably involved in water relationships there, although they may also contain tannins and be involved in plant defence (Nilsen et al. 2014; Tulyananda & Nilsen 2017). Interestingly, idioblasts are not to be found in the bog-dwelling R. saxifragoides and the leaves there are amphistomatous (Nilsen et al. 2014; Tulyananda & Nilsen 2017) - perhaps connected features. G. Khan et al. (2024) could not find much evidence for the importance of features concerned with water relationships, including those just mentioned, in their survey of the epiphytic species of sect. Schistanthe, although epiphytic species did tend to be smaller than the others. For variation in potentially ecologically important leaf and root traits and the evolution of Rhododendron, see Medeiros et al. (2017), and for rate shifts in the evolution of functional traits, where there was a notable slowdown and subsequent stabilization in Rhododendron sect. Ponticum, see Xia et al. 2021).
Melastomataceae, also woody, Piperaceae and Gesneriaceae are the other big epiphytic eudicot families, see also Bromeliaceae and Orchidaceae).
Group 5. Ericoid Mycorrhizae. Many Ericaceae have a distinctive association with fungi, ericoid mycorrhizae (ERM), and this is connected with their ability to grow in open, often rather nutrient-poor heathland habitats in particular. ERM are ectendomycorrhizae - ascomycetes and some basidiomycetes are involved - in which the fungal hyphae form complex and usually coiled intrusions in the exodermal cells of the distinctive very thin and aptly-named hair roots and sometimes also surround the roots with a sheath; such mycorrhizae are not known from other land plants. Hair roots are as little as 40 µm across, monarch, and as close to as thin as a root hair proper gets (hair roots lack root hairs), and in Vaccinium corymbosum, at least, even low-order roots, i.e. not the ultimate rootlets, are also quite thin (Massicotte et al. 2005; Valenzuela-Estrada et al. 2008). Hair roots consist of little more than endodermis, exodermis, tracheid, sieve tube, and companion cell, yet they are relatively long-lived, indeed, secondary growth started in around the fourth order rootlets in Vaccinium macrocarpum, and as this progressed evidence of any fungal association disappeared, the roots becoming involved in transport rather than absorbtion (Atucha et al. 2021). (Although Medeiros et al. (2017) note that the first order roots in the Rhododendron they studied were 0.1-0.8 mm across, "on the larger side compared with most plants" (ibid.: p. 11), Valverde-Barrantes et al. (2017, c.f. in part 2020) found that the diameter of ericoid roots was substantially below the mean diameter for seed plants as a whole, while Atucha et al. (2021) suggest that they are about 100 µm across.) There are other ERM-type morphologies in Ericaceae, and one can think of them as showing various combinations of the development of a fungal sheath or mantle, a Hartig net (and if this is just between epidermal cells or the hyphae penetrate more deeply into the cortex), and hyphal protrusions into the cell (Imhoff 2009: for a summary of ERM, somewhat sceptical in places, see Vohnik 2020); see also below.
Overall, ERM Ericaceae make up ca 1.4% of all seed plants (Brundrett & Tedersoo 2018). They are most commonly found in open, more or less acidic, nutrient-poor and nitrogen-limiting soils in cold to warm temperate climates, that is, in generally rather stressful habitats, and they can also grow on soils with toxic metals (Read 1991: useful summary, 1993; Vrålstad et al. 2000; Cairney & Meharg 2003; Perotto et al. 2012; Garbaye 2013). They are often common in heathlands world-wide - in the past, characteristic families of such heathlands were thought to include Epacridaceae, Prionotaceae, Empetraceae and Vacciniaceae, but all these are now in Ericaceae (Grubbiaceae and Diapensiaceae were the only other families listed, see Specht 1979b; Read 1996). Indeed, if you see plants of Ericaceae when in the field, it is a pretty reliable sign that the soil is acidic. They can grow on old soils where ERM compete successfully for phosphorus with microbes (Turner et al. 2012). Enzymes, etc., produced by the ERM plant/fungus association contribute to the formation of acidic mor humus that ERM plants like and AM plants do not (Read 1991). Ericaceous leaves are well defended chemically, often long-lived, and the plants are efficient at removing N and P from them when they die, so ericaceous litter is of low quality, and the result is persistent, nutrient-poor humus suitable only for the rather slow-growing Ericaceae, their fungal associates, and a relatively few other species (Read 1991; Cornelissen et al. 2001; see also Peay 2016; Van Geel 2020). The hyphae of ERM fungi also have melanin, which, like lignin, is resistant to decay, and is an important component of the sequestered carbon in at least some older boreal forests (Read et al. 2004; Clemmensen et al. 2014; Fernandez et al. 2014, 2015; Lindahl & Clemmensen 2017).
Soils with acid mor humus may not be suitable for many plants, but many Ericaceae thrive on such soils. Thus the protein-tannin complexes that come from Rhododendron may result in nitrogen being held in the stable soil organic matter; this nitrogen is then more easily accessed by the saprotrophic activities of the ericoid fungi than by AM fungi in particular (Read 1991; Wurzburger & Hendrick 2009); the fine ericoid rootlets are very dense, if shallow, and efficiently permeate the soil (try digging up a rhododendron). In ERM, the fungal intrusions in the exodermis are not broken down by the host (Frank 1887; Read 1996; Perotto et al. 2012), but organic N and P taken up by the fungus move to the ericaceous associate; N in amino acids released by the fungus are also taken up by the plant (Cairney & Ashford 2002; E. A. Hobbie & Hobbie 2008; Perotto et al. 2012). ERM fungi show considerable metabolic diversity, having Plant Cell Wall Degrading Enzymes (PCWDs) and being able to break down cellulose, chitin, accessing the N it contains, and some perhaps even degrading lignin in a manner akin to brown rot fungi (e.g. Perotto et al. 2012 and references; Vohník et al. 2012; Martino et al. 2018; Maillard et al. 2023; see also Selosse et al. 2017c; Lebreton et al. 2021); for some details of the movement of nutrients and photsynthesates between Ericaceae and other plants and their associated fungus, see below. In a broad study of various kinds of mycorrhizal mutualists, the ERM ascomycete Oidiodendron maius (the only ERM included) was found to be saprotrophic and can break down Sphagnum peat; it has both cellulases and some lignin-decomposing enzymes that came from its saprotrophic ancestors, and its plant cell wall degrading enzymes may also be involved in penetrating the cell walls of its host (Kohler et al. 2015). Unlike orchid mycorrhizae, ERM are not involved in carbon uptake by the host (Kohler & Martin 2017), indeed, ECM fungi may use host photosynthesate.
Note that there can be a close association between ERM plants and ECM trees in forests in the temperate eastern U.S.A. leading i.a. to an overall higher soil C:N ratio - the ERM plants increasing C, the gymnosperms decreasing N, and there was an increase in the basal area of gymnosperms and of the cover of the ERM shrubs (Ward et al. 2021). Gerz et al. (2017) comment on the relatively broad niches of ERM plants. See also elsewhere for more general ecological aspects and again for ERM and the ecologically somewhat similar ECM associations.
Interestingly, distinctive cavendishioid mycorrhizae - here the ericoid roots are short, and there is both a fungal sheath and a Hartig net - have been found in (hemi-)epiphytic Vaccinieae from Andean South American forests (Setaro et al. 2006, 2008). Kottke et al. (2008a) also discuss the mycorrhizal fungi associated with epiphytic Ericaceae in the Andes, interestingly, bushy Ericaceae growing in open habitats have ordinary ERM, so differing from the fungal associations in the forest-dwelling species (Kottke et al. 2008b). For more on the general issue of situations where only a few species dominate over very large areas, see elsewhere.
The adoption of the ERM habit in Ericaceae can perhaps be dated to at least 72-66 Ma, Leucothoe eocenica (Vaccinioideae) being reported from deposits of that age in Europe (Knobloch & Mai 1986; Strullu-Derrien et al. 2017), and if the identification is correct - the fruits are tiny, ca 1.8 x 1.8 mm, although apparently ripe - the clade characterized by ERM plants, the [[Cassiopoideae + Ericoideae] [Harrimanelloideae [Epacridoideae + Vaccinioideae]]] clade, must be substantially older. Indeed, Martino et al. (2018) dated the age of the common ancestor of the four ERM fungi (ascomycetes, Leotiomycetes) that they sequenced at ca 118 Ma, agreeing with the age of the family (see also Lebreton et al. 2021), ca 117 Ma in Schwery et al. (2014), but in this case the origin of the ERM association would have to be younger given the relationships/mycorrhizal associations just mentioned - and note that the 117 Ma age for the family itself is a remarkably high estimate. Wagstaff et al. (2010) date the ERM clade to ca 65 Ma and the estimate is around 77 Ma in Z.-Y. Liu et al. (2014).
Note that Enkianthus has arbuscular mycorrhizae (AM) of the Paris type (Abe 2005; Obase et al. 2013, but c.f. in part Vohnik 2020); the mostly ECM [Arbutoideae [Monotropoideae + Pyroloideae]] make the next branch up. AM are also reported in Rhododendron from the Himalayas (Chaurasia et al. 2005). I have seen few accounts of mycorrhizae in Clethraceae or Cyrillaceae, but the distinctive ERM associations may well have become established only after Enkianthus and the [Arbutoideae [Monotropoideae + Pyroloideae] clades diverged from the rest of the family. (Although Diapensiaceae are reported to have ERM fungi (Brundrett & Tedersoo 2018), the origin of such associations there are likely to be independent of those in Ericaceae.)
Group 6. The Holomycoheterotrophic Habit. This is a version of hyperparasitism in which the ericaceous member of the association has more or less lost the ability to photosynthesize, its need for C, at least, being met by its ECM fungal associate, which in turn gets its C from its other plant associate, a normal photosynthesizing plant (see Björkman 1960). The holomycoheterotrphic habit has arisen at least twice in Ericaceae, in Monotropoideae and Pyroloideae (Zimmer et al. 2007; Hynson et al. 2013; Lallemand et al. 2016; Tedersoo & Brundrett 2017; c.f. Cullings 1994). Unlike most other angiosperms, giberellic acid had little effect on germination in Pyroloideae, at least, the exceptions being Orchidaceae with holomycoheterotrophic associations (references in Miura et al. 2023).The relationship between fungi and Ericaceae is closest in the echlorophyllous and holomycoheterotrophic Monotropoideae. Here the underground parts can be massive and the roots thick, some 200-600 µm across, often with a thick fungal mantle (already evident in germinating seedlings), a Hartig net, and fungal pegs (Imhof et al. 2013). Basidiomycetes are often the ECM fungi involved (Bruns et al. 2002; Garbaye 2013). The specifiity of the plant—fungus association varies, and it is sometimes cloase to a 1:1 association (Cullings et al. 1996). In Pyroloideae, Pyrola aphylla (a form of P. picta) is a holomycoheterotroph at times as are albino variants of P. japonica, other members of the subfamily are mixotrophic or fully heterotrophic - for example, ca 50% of the C needs of Orthilia secunda are met by its fungal associate (Tedersoo et al. 2007a; Whitfield 2007; see also Hynson et al. 2009b; Selosse & Weiß 2009; Bowler et al. 2017; Matsuda et al. 2020). Pyrola aphylla may have small green leaves and the fungi associated with this species show no particular specificity, as in other more photosynthetically conventional species of Pyrola (Hynson & Bruns 2009; Johansson et al. 2017). Both C and N move from the ECM fungal associates to chorophyllous Pyroloideae and echlorophyllous Monotropoideae alike (Zimmer et al. 2007; Tedersoo et al. 2007a; Matsuda et al. 2012; Johansson et al. 2015; Lallemand et al. 2016); Hynson et al. (2016) discuss N metabolism, while Kranabetter and MacKenzie (2010) noted the distinctiveness of the N metabolism in Pyroloideae when compared with that of other Ericaceae with ERM, emphasizing their probably mixotrophic nutrition. Lallemand et al. (2016) discussed the evolution of the holomycoheterotrophic habit and of mixotrophy in this whole clade (which includes Arbutoideae) in terms of predispositions, although it was unclear what these might be. Pyrola japonica s.l. included more or less aphyllous and mycoheterotrophic and leafy, "normal" haplotypes (Shutoh et al. 2016); fully echlorophyllous forms of this species have been discovered (Shutoh et al. 2020). Matsuda et al. (2020) suggested that three different forms of this species had different mycorrhizal preferences, especially for different species of Russula. Moneses uniflora showed geographical variation in the extent of partial mycoheterotrophy it showed (Hynson et al. 2015). See also Johansson et al. (2017) for discussion about Swedish taxa. Of course, there are also mixotrophic and mycoheterotrophic species in Orchidaceae and echlorophyllous seedlings are widespread there (see elsewhere for more on mycoheterotrophy in general; L. Cai (2023: Table 1) also discusses the evolution of holomycoheterotrophy.
Hashimoto et al. (2012) found that in Pyrola asarifolia from Hokkaido, Japan, non-ECM fungi (Sebacinales-Serendipitaceae: Weiß et al. 2016) were associated with the plant as it germinated, at that stage the plant-fungus relationship being rather like that between germinating orchids and fungi; different fungi were associated with the roots of the adult plant, and these were ECM fungi also associated with Betulaceae growing in the same area. However, the duration of the subterranean, non-photosynthetic, holomycoheterotrophic stage and other details of this early relationship are poorly known (Hynson et al. 2013; see Johansson et al. 2017 and references for some details). In some Pyroloideae shoots do not appear above ground every year, as also in some Orchidaceae, the phenomenon of vegetative dormancy in which mycorrhizae can play a role (Shefferson et al. 2018; Hurskainen et al. 2018).
Other matters. Lens et al. (2003, 2004a, b, c) looked at ecological aspects of the woody anatomy of various members of the family. They found correlations between aspects of anatomy with latitude, and also with life form and precipitation. Erica is a notable component of the Afrioalpine flora (Brochmann et alk. 2021).
Pollination Biology & Seed Dispersal. Overall, surprisingly little is known about details of pollination in the family (c.f. in part de Luca & Vallejo-Marín 2013). The anthers of most Ericaceae have functionally apical pores and may be grouped in the centre of the flower, common in buzz-pollinated plants, but nectaries are also very frequent, which makes for a rather unusual combination (but see also Boraginaceae); the nectar may on occasion appear to be coloured (Niu et al. 2025). There are some reports of buzz pollination: In Pyroloideae, for instance, the lack of an endothecium and absence of a nectary there are associated with buzz pollination (Liu et al. 2011), and as in other such clades pollen is the reward (but not always: see Knudsen & Olesen 1993), while buzz pollination is also reported from Conostephium (Epacridoideae), and here the pollen comes out of a narrow opening at the apex of the corolla tube, and again, there is no nectary (in Epacridoideae the anthers open by slits). Buzz pollination also occurs in genera like Sprengelia, although here the petals are spreading (Houston & Ladd 2002). Vibrations of the anther that result from bumblebees working the nectariferous flowers of Rhododendron may facilitate pollination, but the bees do not buzz the flowers, at least in the species studied (King & Buchmann 1995). On the other hand, in Vaccinium and Erica growing in temperate heaths in Europe, both genera nectar-producing, buzz pollination and nectar foraging may occur on the same plant (Andrena and Bombus are the bees), and both may result in effective pollination - and both activities were also noted on Zenobia (Dorr 1981). The oligolectic Melitta americana simultaneously sonicates the flowers of Vaccinium stamineum (it is the major pollinator) and takes nectar (Cane et al. 1985). A diversity of bees - 112 species, solitary Halictidae (polylectic) and Andrenidae (poly- and oligolectic) being most common, one third of all species trapped - were active during the period of highbush blueberry flowering on farms in Michigan alone (Tuell et al. 2009). Such bees may buzz the flowers (e.g. Bombus, also the oligolectic solitary bees Andrena carolina and Habropoda laboriosa) or visit the flowers to collect nectar (e.g. Apis, also the solitary bee Megachile rotundata); nectaring bees were least efficient in pollination (Fowler 2016; Danforth et al. 2019 for a summary). Buzz-pollinating bees are generally important in the pollination of blueberries, cranberries, etc. (Cooley & Vallejo-Marín 2021). Ericaceae are a major food resource - both pollen and nectar - for bumblebees like Bombus jonellus in the early spring and late summer (Moquet et al. 2016, 2017a, b). For more on buzz pollination, see elsewhere. Pollination by bees, especially bumblebees, is common in temperate and arctic-alpine members of the family, almost regardless of their floral morphology. Thus bumblebees pollinate alpine species of Rhododendron, Vaccinium (these genera both have species with polypetalous and tubular urceolate-campanulate flowers in this habitat), Elliottia (polypetalous), Phyllodoce, Pieris, Chamaedaphne, Cassiope, Gaultheria (all urceolate to campanulate), etc. (Heinrich 1979; Ranta & Lundberg 1981; Tomono & Sota 1997; Kudo et al. 2011 and references), as well as being the main pollinators of the mycoheterotrophic Monotropoideae (Waterman et al. 2013). Other pollinators are also effective, and selfing also occurs. Bees, mostly Bombus, were the main visitors to the flowers of Kalmia latifolia; apparently they were nectaring (although the nectar is poisonous), in the course of which they triggered the stamens, the anthers being held in pockets of the corolla, and the pollen was flung out - maximum velocity 3.5 m s-1, maximum acceleration 4,100 m s-2 (Switzer et al. 2018 - note that species like K. procumbens are polypetalous, and so are pollinated in a different way).
Bird pollination is particularly common in the Andean Vaccinieae, where it has been dated to ca 14.5 Ma (Stiles 1981 and references; Kriebel et al. 2023). There are ca 600 species of Vaccinieae in the tropical Andes, over 500 species being found above 1,000 m, and the centre of their diversity is the Colombia-Ecuador region, where hummingbirds, which pollinate most of these species, are also maximally diverse (Luteyn 2002); Gentry (1982) discussed the diversity of bird-pollinated taxa of putative Gondwanan origin in tropical and premontane parts of the northern Andes. see also Gesneriaceae-Gesnerioideae, Lamiaceae-Nepetoideae-Salviinae, Bromeliaceae and Zingiberales-Heliconiaceae; for further details of hummingbird pollination, see elsewhere). Several other factors have been implicated in this Andean diversification, including the adoption of the epiphytic habitat by some of these plants, the microtopographic diversity associated with the uplift of the Andes, etc. (Luteyn 2002) - but remember, little is known about their phylogeny. It has been suggested that vibrations of the flowers caused by the hummingbirds wings help dislodge pollen from the anthers of these plants (Murcia 2000) - whirr pollination, hummingbirds' answer to bees... Pseudanthia are reported from Cavendishia (Baczynski & Claßen-Bockhoff 2023). For an interesting mutualism involving the hummingbird Basilinna xantusii and Arbutus peninsularis in Baja California, see Abrahamczyk et al. (2017). There is probably bird pollination in many of the large, red, often tubular-flowered Indo-Malesian Vaccinieae like Dimorphanthera (ca 75 spp., some were recently in Vaccinium), and Paphia (20 spp., ex Agapetes s.l.), both from Malesia, and Agapetes s. str. (110 spp.), from mainland Southeast Asia. Interestingly, stem-group hummingbirds were also nectar-eaters, and they are known only from the Old World, where fossils have been dated to ca 45 Ma (Mayr 2005, 2009), indeed, it has been suggested that Agapetes s. str. may initially have been pollinated by hummingbirds, and McGuire et al. (2014) has suggested that hummingbirds moved to the New World via Beringia, consistent with such an idea. However, although species in Agapetes have some of the features of the hummingbird pollination syndrome like pendant flowers, they are now apparently pollinated by honeyeaters and their like - sunbirds, Nectariniidae, seem not to be involved (Kriebel et al. 2023), although the story is complicated - as Wikipedia noted (as of x.2024) "For the Southern Asian birds formerly known as honeyeaters, see Sunbird". There is further discussion elsewhere.
A variety of pollinators visit South East Asian-Malesian species of Rhododendron, passerine birds being quite prominent pollinators in the Himalayas s.l. (Z.-H. Huang et al. 2017; Y.-P. Song et al. 2018). There is also bird pollination in Malesian vireya rhododendrons (= Rhododendron sect. Schistanthe), and this group seems to have moved through the archipelago west to east, New Guinea and places immediately to the south and east being inhabited by a separate and very speciose clade of subsection Euvireya (Goetsch et al. 2011; Soza et al. 2022); here, too, there is bird pollination - and nectar-eating mites may use the birds to travel from flower to flower (Stevens 1976). Basnett et al. (2019) looked at some rhododendrons along an altitudinal gradient in the Sikkim Himalayas and found that features like corolla length and nectar volume decreased with elevation, nectar concentrations were increased; birds visited the species with larger flowers with more and more dilute nectar at lower elevations, and bumblebees and flies the smaller-flowered species with more concentrated nectar at higher elevations, indeed, bees are another common pollinator of Rhododendron (see references above, also Berry et al. 2017). The centre part of the adaxial corolla lobe is often patterned, and this area absorbs UV light, the rest of the corolla reflecting it; both bees (and butterflies) have UV vision and use the patterning as a honey guide. Interestingly, white-flowered species pollinated by hawkmoths lack such guides - no UV in the evening (Kawakita et al. 2024).
Viscin threads are known from about 900 species, especially in Rhodoreae and a number of other Ericoideae, and they have also been reported from Gaultheria (Lu et al. 2009). Rather unusually (but like the threads of Onagraceae) these threads are made up of sporopollenin (Halbritter et al. 1997). It would seem that such threads should increase the efficiency of placement of pollen grains on the pollinator, but little is known about their role in pollination (e.g. Zetter & Hesse 1996). Both ericaceous taxa with viscin threads and those that apparently have buzz-pollinated flowers often produce nectar, unlike other taxa with viscin threads and flowers with the classical buzz-pollination syndrome (e.g. Moquet et al. 2017b). Moquet et al. (2017b) noted that bumblebees were effective pollinators of two species of Vaccinium and Erica tetralix, and although the flowers might indeed be buzzed, the bees then collecting pollen, effective pollination was predominantly by nectar-collecting bees in V. myrtillus and E. tetralix. Ericaceae were the main resource for bumblebees in the heathlands of Belgium, especially in the early spring and late summer (Moquet et al. 2017a: focus on pollen); Moquet et al. (2017b) noted that hoverflies were important visitors to Calluna vulgaris there.
In both bird- and bee-pollinated flowers the pollen may initially come from the flower in little puffs, almost explosively. Thus Geerts and Pauw (2010) noted that in the Cape species of bird-pollinated Erica that they were examining the apices of the anthers were initially tightly pressed to each other and to the style so forming a complete ring around the style, the corolla tube being almost blocked; pollen could not get out. However, when Nectarinia (= Anthobaphes) violacea, the orange-breasted sunbird, visited the flower for nectar, as its bill entered the tube it disturbed the anthers and they separated, only then releasing the pollen as little puffs; the bees that also visited the flower could not break this ring of anthers (Geerts & Pauw 2010). Similar puffs of pollen were produced by both hummingbirds and bees as they visited species of three unrelated genera of Vaccinioideae in Brazil (Freitas et al. 2006); details of the mechanisms involved differed, and the birds and bees mostly visited different species.
In southern Africa around 100 species of Erica in the Fynbos, perhaps 66 species from the southwestern Cape alone, are pollinated by a few species of nectariniids, but particularly by Anthobaphes violacea These bird-pollinated species often had colour polymorphisms or flowers with more than one colour (Rebelo et al. 1984, 1985; Rebelo 1987; S. D. Johnson 2010; Geerts & Pauw 2010). There was clearly no co-evolution here! Overall, Rebelo et al. (1985) found that 339 (ca 80%) species of the 426 species of Erica from the southwestern Cape were pollinated by insects, 66 species (15%) by birds, and 21 (ca 5%) by wind. Members of the latter group had notably small flowers with large cyathiform/peltate stigmas - note that wind-pollinated species in the minor "genera" like Philippia and Salaxis, which also have small flowers, etc., and a number of which have single pollen grains, were not included, although they are now placed in Erica. Rebelo et al. (1985) also noted that of the insect-pollinated taxa, 31 species were likely to be pollinated by the long-tongued Nemestrinidae flies (= rhinomyophily), distinctive pollinators in that part of the world, although perhaps rather less is known about these flies than one would like, only about half the nemestrinids in South Africa having been described (Theron et al. 2023). All told, perhaps 73 species of Erica are pollinated by such flies (Lombardi et al. 2021); for pollination in Erica, see also McCarren et al. (2024) who noted that the pollen transfer efficiency of nemestrinids and nectariniids was higher than bees, i.e. less pollen was lost in transport (bees groom themselves, and travel clean) - nevertheless, here, as elsewhere, bee-pollinated flowers seem to do quite well... Wronska-Pilarek et al. (2018) looked at the pollen morphology of some of the Cape Erica. Nicolson (2022) noticed the combination of tubular flowers, sucrose-rich pollen and specialist pollinators in Erica.
Secondary pollen presentation - the pollen sticking to the hairs of the recurved corolla tips by its copious pollenkitt - occurs in the Australian Acrotriche, which is probably pollinated by the marsupial mouse, Antechinus stuartii (McConchie et al. 1986). Keighery (1996) outlined pollination in western Australian Epacridoideae, where bee pollination is common, but there is also pollination by other insects and, in a few species at least, birds. Pollination by Prosoeca, a long-proboscid fly, has recently been demonstrated in the South African Erica aristata, and such pollination is estimated to occur in around 9% of the species in the genus, i.e. in around 73 species (Lombardi et al. 2021). Interestingly, stickiness of the outside of the corolla is common in Cape species of Erica in particular, and it is thought to prevent nectar robbing, for example from flowers pollinated by birds and long-proboscid flies (McCarren et al. 2021). Wind pollination in Ericaceae has evolved at least twice, in Empetrum and its relatives and in taxa that were placed in genera like Philippia, but now included in Erica. Both these groups have expanded stigmas.
Given that Ericaceae quite often are dominant plants in the places where they grow, little has been written about the importance of Ericaceae as nectar sources for their pollinators, although there is often mention of the amount of nectar produced by the flowers. However, Baude et al. (2015) found that Calluna vulgaris and Erica cinerea were two of the four species that together provided over 50% of nectar for the bee population in Great Britain in 2007, Calluna being number 2.
Fritsch et al. (2020b) discuss gynodioecy in New Zealand species of Gaultheria; the species involved are probably descended from South American species that were also gynodioecious.
Fleshy-fruited taxa, whether the calyx, the pericarp or the walls of the inferior ovary are fleshy, predominate in Vaccinieae. In both the Old and New Worlds some Vaccinieae have seeds with a mucilaginous testa and a chlorophyllous embryo, perhaps mistletoe-type seeds; plants with such seeds are generally epiphytic or epilithic (pers. obs.). Seeds of other Vaccinieae, and of other Ericaceae in general, have a hard testa and white embryos. In the New World, Vaccinieae with fleshy fruits are much eaten by tanagers (see also Myrtaceae), which tend to remove seeds greater than 2 mm long from the fruits before ingesting them (Stiles & Rosselli 1993); the seeds of Vaccinieae are often a bit bigger than this. There is scatter-hoarding by rodents of the stones of the dry semi-drupaceous fruits found in some Arbutoideae in the New World (Parker & Stickrod 2022). Seeds in fleshy fruits of the mycoheterotroph Monotropastrum humile in Japan are dispersed by camel crickets (Rhaphidophoridae: Tachycines elegantissima), as are the seeds of unrelated mycoheterotrophs (Suetsugu 2017), indeed, the woodlouse Porcellio scaber, to 11 mm long, also disperses the seeds of M. humile, and is the smallest animal that disperses seeds internally - P. scaber is introduced in Japan (Suetsugu et al. 2024b).
Plant-Animal Interactions. Bejaria racemosa, appropriately named "tar flower", traps numbers of insects on its flower buds and fruits with its glandular hairs. Eisner and Aneshansley (1983) found that the stickiness of the secretion of the hairs and that of Tanglefoot was similar, and suggested that the plants might get nitrogen from the decay of the trapped insects as it entered the nutrient-poor sandy soils on which the plants grew. Mirid bugs are associated with a sticky-leaved (again, glandular hairs) species of Rhododendron from Japan; the benefits to the insect from eating the carcasses of other insects that had become stuck to the leaves were examined (Sugiura & Yamazaki 2006; for such mirids, see Wheeler & Krimmel 2015), also LoPresti et al. (2015) and elsewhere.
The berry-like fruits of Empetrum, along with those of five more shrubs of which four are other Ericaceae (two species of Arctous and two of Vaccinium), also Rubus chamaemorus, are important sources of food for birds (Lorion & Small 2021).
Vaccinium is an important host plant for lepidopteran larvae in the contiguous United States of America, being included in the top 10 such plants (Narango et al. 2020).
Nectar-eating mites in the flowers of Schistanthe rhododendrons growing at higher altitudes in Papua New Guinea may use birds probably pollinating those plants to travel from flower to flower (Stevens 1976).
Grayanotoxins, polyhydroxylated cyclic diterpenes, are quite common, if scattered, in Ericaceae; they are neurotoxins, fatal to some insects. However, there is both inter- and intraspecific variation in whether or not, where, when (changes during the growth of the plants) and the amount of grayanotoxin produced (Fattorini et al. 2023: Rhododendron the focus); grayanotoxins are toxic to honeybees, but not the pollinating Bombus terrestris, in R. ponticum (Nicolson 2022), but little seems to be known about how grayanotoxins function as plant protectants. "Mad honey" (see below) is a honey with high levels of grayanotoxin. For the distribution of grayanotoxins (andromedotoxin is a particular form of this), see C. Zhou et al. (2012), S. A. Jansen et al. (2012), Y. Li et al. (2013) and C.-H. Li et al. (2019) and references.
Plant-Bacterial/Fungal Associations. For Ericaceae-mycorrhizal associations in general, see also Cullings (1996) and Smith and Read (1997) and the discussion in Ecology & Physiology above. Enkianthus, sister to the rest of the family, has arbuscular mycorrhizae (AM) of the Paris type (Abe 2005; Obase et al. 2013), and it lacks hair roots, however, Vohnik (2020) suggested that it might indeed form ERM. AM, along with ERM, are also reported from some Hawai'ian Ericaceae, for example, species of Vaccinium formed both AM (on coarser roots) and ERM (on fine roots) associations (Koske et al. 1990).
Arbutoideae, Pyroloideae and Monotropoideae, all in the same immediate clade above, are commonly associated with the largely ECM basidiomycete Agaricomycete-Sebacinales-Sebacinaceae (Selosse et al. 2007; Toju et al. 2016; Weiß et al. 2016; see also Brundrett 2017a; Tedersoo 2017b; Tedersoo & Brundrett 2017 for literature, ages, etc.), although other basidiomycetes may be involved (Cullings et al. 1996). Although there are hyphal coils in the root cortical cells, the sheath and Hartig net make the association quite ECM-like (Molina et al. 1992; Hynson et al. 2013). ECM Agaricales were found associated with Arctostaphylos alpina (Toju et al. 2016), while suilloid basidiomycetes such as Rhizopogon may be associated with Arbutoideae and Monotropoideae - and as ECM on co-occuring Pinaceae (Bruns et al. 2002 and literature), indeed, Pterospora andromedea seems to form associations with the Rhizopogon subcaerulescens species group alone (Cullings et al. 1996). Similarly, Di Rita et al. (2020) found that Amanita muscaria and a number of other fungi in central Italy that now have extra-zonal distributions seem to have moved on to Arbutus unedo from conifers that have since died out in the area (and the Amanita is sister to a clade of American populations...). Fungi associated with individual species of the echlorophyllous mycoheterotrophic Monotropoideae are mostly ECM basidiomycetes (Bidartondo & Bruns 2001, 2002; Bidartondo 2005: questions as to the identity of the plants compromise earlier literature; Hynson & Bruns 2010). Several species of the basidiomycete Russula can be associated with Monotropa uniflora from the one area (S. Yang & Pfister 2006), although there is often considerable fungus-monotropoid specificity, the specificity being more on the side of the monotropoid partner (Bidartondo & Bruns 2001, 2002; Bruns et al. 2002). Indeed, suilloid Rhizopogon were found to be associated with Pterospora and Sarcodes alone, these latter being sister to other Monotropoideae, and these other Monotropoideae were associated with Gomphales, Thelephorales and Tricholomatales, which were in a clade sister to Suillloids... (Bidartondo & Bruns 2001). The specificity of the fungal associations here increases during plant growth (Bidartondo & Bruns 2001) - and this may also happen in some partly mycoheterotrophic species of Pyrola (Johansson et al. 2017). Thus Hashimoto et al. (2012) found that in P. asarifolia from Hokkaido, Japan, non-ECM Sebacinales-Serendipitaceae were associated with the seedling - which looks almost like a thalloid liverwort - and they formed hyphal coils in the epidermal cells, but not a sheath. However, the fungi in roots of the adult plant, although including some Sebacinales, also included other basidiomycete ECM fungi associated with Betulaceae growing in the same area, a switch also found in Orchidaceae (see also Jacquemyn & Merckx 2019). Note that both Pyroloideae and Arbutoideae show less specificity in their fungal associates than do Monotropoideae (e.g. Kennedy et al. 2012; Jacquemyn & Merckx 2019). Interestingly, 13C and 15N were notably high in P. subaphylla and P. aphylla.
Turning now to ERM associations, the morphologies of these associations varies within the family, but the distinctness of the types that have been described needs to be be confirmed. Fungal sheaths have been reported from the roots of a number of Vaccinieae (e.g. Setaro et al. 2006; Vohník et al. 2012; Yagame et al. 2016). These include the so-called "cavendishioid" (see above) and "sheathed ericoid" mycorrhizal types, and as the sampling of tropical montane and southern hemisphere Ericaceae improves, we may expect to find yet more variation in Ericaceae-fungal relationships there. Fungi in both ericoid and cavendishioid mycorrhizae are Sebacinales-Serendipitaceae, often saprophytic or endophytic (Setaro et al. 2006; Selosse et al. 2007; Weiß et al. 2016: the two families of Sebacinales are sister taxa, but the particular groups of fungi under discussion are not very close; Weiß et al. 2016: Fig 3), indeed, the route to these ERM associations may be via an endophytic stage (Strullu-Derrien at al. 2018). Many Leotiomycetes (ascomycetes) are ERM fungi and they increase in diversity towards the poles where Ericaceae-dominated heath vegetation is conspicuous (Tedersoo et al. 2014b; Wardle & Lindahl 2014). Indeed, things get complex. Ascomycetous dark septate endophytes (the dark colour = melanin) are found in the roots of alpine Ericaceae from the Rocky Mountains (Stoyke & Currah 1991), and there are intermediates between ERM (and ECM) and dark septate endophytes in Rhododendron (Vohník & Albrechtova 2011; Lukesová et al. 2015). For various kinds of ascomycete associations, see Tedersoo et al. (2009a), Kernaghan and Patriquin (2011: Cenococcum geophilum on conifers, various ascomycete endophyte communities). Vohník et al. (2013) found a complex series of interactions between the ECM Picea abies and its endophytic communities - both ERM and dark septate endophytes were involved, and fungi from both groups can form ECM and ERM associations (for the latter, see also Lukesová et al. 2015). Meliniomyces variabile (part of the Rhizoscyphua ericae aggregate), is one of these fungi, but R. ericae itself was not found as an endophyte on spruce; Vohník et al. (2013) thought that the spruce-endophyte habit might be an alternative for ERM fungi in habitats where there weree no suitable ericaceous hosts. Lukesová et al. (2015) found 380 ericaceous mycobiont species in the Bohemian Forest (Czech Republic).
Note that some fungi may be ERM or ECM, depending on the host, and the distinction between the two mycorrhizal types, although at first sight quite substantial, is within the capacity of a single fungal isolate to express (e.g. Grelet et al. 2009: ECM and Pinus silvestris/ERM and Vaccinium vitis idaea; Imhoff 2009; Martino et al. 2018; see elsewhere). Similarly, ERM fungi in Epacridoideae may be dark septate endophytes when growing on other plants (Chambers et al. 2008). Toju et al. (2016) described the numerous fungi associated with Ericaceae growing in extreme Alpine environment in Japan. The ascomycete Helotiales dominated, although Sebacinales-Serendipitaceae were also common, and they are also associated with members of Ericoideae, Epacridoideae and Vaccinioideae (see also Setaro et al. 2006, 2008; Selosse & Weiß 2009; Selosse et al. 2009; Weiß et al. 2009, esp. 2011; Toju et al. 2016); the fungi associated with Cassiopoideae are unknown. Bougoure et al. (2007) detailed the variety of ERM fungi associated with Vaccinium and Calluna, suggesting that the ERM fungi on the two differed, but c.f. J. F. Walker et al. (2011). Basidiomycete associates may be proportionally particularly common in Vaccinioideae, including the tropical members (Bougoure et al. 2007; Setaro et al. 2006, 2008; Yagame et al. 2016). Interestingly, the basidiomycete isolated from the sheathed ERM roots of Vaccinium myrtillus was quite distinct from other fungi associated with Ericaceae (Vohník et al. 2012). Kohout et al. (2017) found some effect of host plant on ECM (and endophyte) community structure, but not fungus richness at high altitudes in Papua New Guinea, interestingly, fungi that are ERM on Ericaceae might be endophytic on non-Ericaceae. For the mycorrhizae of Epacridoideae in particular, see Cairney and Ashford (2002) and Chambers et al. (2008) and references; the latter emphasize that dark septate endophytes on a variety of plants in southeast Australian sclerophyll vegetation may also form ERM associations with epacrids.
At any one locality, fungi particularly common on individual species of Ericaceae may differ, that is, there is some host specificity, and ericaceous species shared fewer fungal associates than might be expected by chance (Toju et al. 2016: "anti-nestedness"). There was geographical variation in the mycorrhizal associates of Moneses uniflora, although they were in the same family, the basidiomycete Atheliaceae (Hynson et al. 2015; see also Massicotte et al. 2008 for Pyroleae). Furthermore, plant-fungal relationships in Argentinian species of Gaultheria differ from those in the northern hemisphere Gaultheria (Bruzone et al. 2013). However, Setaro and Kron (2011) found some clades of Sebacinales-Serendipitaceae on both North and South American Vaccinieae, while Andean Orchidaceae and Vaccinieae growing together were associated with different but closely related clades of Serendipitaceae (Setaro et al. 2013). The specificity of Arctic ERM fungi, at least, is low (J. F. Walker et al. 2011; Timling & Taylor 2012), different ericaceous genera in the one place being linked by an ERM network - even if plots only 2-3 m apart had rather different fungi (Kjøller et al. 2010). Similarly, species of tropical American Serendipitaceae can form associations with more than one species of Ericaceae (Kottke et al. 2008; see also Weiß et al. 2011). However, Van Geel et al. (2020) found that in Europe, at least, effects of both environment/habitat and host plant led to strong host plant preferences for particular fungi.
Rinaldi et al. (2008) suggested that the diversity of fungi associated with Ericaceae might not be very high, but their figure of 15 species is a gross underestimate, more species than this being found associated with Arbutus menziesii at a single site in Oregon (Kennedy et al. 2012), while 224 fungal OTUs were detected from three species of Ericaceae from Alaska, and these OTUs belonged to 14 distinct clades in the ascomycetan Helotiales alone (J. F. Walker et al. 2011; Timling & Taylor 2012). At least 150 species of fungi is the estimate in van der Heijden et al. (2014), but clearly this number is still far too low. Recent studies that focus on Sebacinales (e.g. Weiß et al. 2011, see also 2016) suggest that quite a number of species in that group also grow with Ericaceae. All told, rather little seems to be known about the number of fungi associated with Ericaceae and their distributions (see also Vohnik 2020), a situation not helped by uncertainty over fungal species limits (Kohout 2017: see e.g. Oidiodendron).
As with other mycorrhizal associations, details of how the fungus affects the plant depend in part on the genotype of the latter (Brody et al. 2019). The relationships between the partners and the pathways of nutrient flow can be complex, and mycorrhizal networks can involve other than echlorophyllous Pyroloideae and Monotropoideae. For example, a member of Sebacinales-Sebacinaceae found growing on Diphasiastrum alpinum (Lycopodiaceae) and also on Calluna vulgaris from the same habitat may allow the movement of nutrients from the latter to the former (Horn et al. 2013). In western North America the arbutoid madrone, Arbutus menziesii, is a common subordinate tree in forests with ECM Fagaceae and Pinales that occupies over 3.9 x 106 acres in California alone (Waddell & Barrett 2005). Its diverse fungal associates are also found on other angiosperms and in particular Pinaceae (Pseudotsuga and Pinus) in Oregon (Kennedy et al. 2012). Arbutus menziesii resprouts after fire and it may be a source of fungal inoculum for newly-germinated seedlings of associated Pinaceae, so facilitating their regeneration (Kennedy et al. 2012 and references), although many temperate, but not Arctic, ECM associations develop from propagules in the soil (Hewitt et al. 2017). As Kühdorf et al. (2015: p. 110) noted, the arbutoid "C[omarostaphylis] arbutoides is a refuge plant for ECM fungi as it shares these fungi with tropical ECM trees such as Quercus costaricensis". The ascomycete Rhizoscyphus ericae (= Hymenoscyphus ericae, Helotiales), is also found associated with Podocarpus oleifolius (Podocarpaceae), Weinmannia pinnata (Cunoniaceae) and Graffenriedia paniculata (Melastomataceae) in the New World (Dalling & Willey 2025); Pezizella ericae, Pezoloma ericae, etc. (these = Hyaloscyphus hepaticicola - Vohnik 2020) is a very common associate of the hair roots of ERM North Temperate Ericaceae; this fungus can also be an ECM associate of Pinus and Abies growing with Ericaceae (Read 1996; Grelet et al. 2009, 2010; see also Villarreal-Ruiz et al. 2004; Martino et al. 2018). Rhizoscyphus also forms mycorrhizal associations with Jungermanniales-Schistochilaceae and other leafy liverworts, colonizing their rhizoids (Duckett & Read 1995; Upson et al. 2007; Pressel et al. 2006c, 2008); other ascomycetes are also involved, including Cenococcum and Geomyces. (See Lofgren et al. 2018 for cautionary comments about host specificity.) Note that the result of ECM/EM associations, and including associations with Rhizoscyphus, is a soil with a very deep organic horizon, the material there being low density, unstable and dessication-prone, and forests with ECM/ERM have around 70% more C per unit nitrogen than soils with AM (Averill et al. 2023; Dalling & Willey 2025). Furthermore, some ERM fungi form endophytic associations (ERM-Epacridoideae/other families-dark septate endophytes: Chambers et al. 2008; ERM-Vaccinium myrtillus/ECMPicea abies: Vohnik et al. 2013), and some endophytic fungi are physiologically quite similar to ERM fungi (Martino et al. 2018). For additional information, see papers in Martin (2017).
Ericaceae like Vaccinium may also have ascomycetous dark septate endophytes which form microsclerotia in the roots, but some of these fungi, like Phialocephala glacialis, formed structures approaching ERM, while Acephala species produced both similar intermediate structures and full-blown ERM (Lukesová et al. 2015). Ngugi and Scherm (2006 and references) discuss fungal associates of Vaccinium including Monilinia vaccinii-corymbosi (an ascomycete, Helotiales; other species also involved). Conidia are produced on the leaves and the conidial patches may seem like floral calyxes to the pollinator, which moves spores to flowers where they behave just like pollen grains but parasitize the fruits (mummy berries); they can cause serious losses of the berries.
Endophytes are common (Petrini 1988; Sieber & Grünig 2013: general summary); Koudelková et al. (2017) discuss the ability of endophytes to tolerate the essential oils of Rhododendron tomentosum.
Host preferences of the basidiomycete rust fungi Chrysomyxa and Exobasidium link the old Empetraceae with Ericaceae and Ledum with Rhododendron; Chrysomyxa is also known from Arctostaphylos, etc., apparently not from Enkianthus, but also from Ilex, while Exobasidium is also found on Theaceae, Symplocaceae and Lauraceae (Savile 1959, 1979b; see Jackson 2004 for possible codivergence).Exobasidium includes around 50 species or more; it is a basidiomycete that belongs in a class by itself, and its members form leaf spots and distinctive galls of various kinds – including converting leaves of Vaccinium into remarkable linear structures. Little is known about the life cycle of Exobasidium, but Newell et al. (2023) suggested that insects were involved in the transmission of spores of E. maculosum in commercial V. virgatum (see also Monilinia below). Interestingly, the reflectance of Exobasidium galls is different from that of leaves and more like that of flowers, and insect-attracting volatile organic compounds are also produced by the gall.
On a very different subject, Urbaneja-Bernat et al. (2020) found that the concentrations of sugars and proteins in the guttation droplets on the leaves of Vaccinium corymbosum were remarkably high (that of the former was higher than in floral nectar). The droplets attracted everything from spiders and ants to gall midges, very largely carnivorous, but, interestingly, not aphids, mosquitoes, or plant herbivores. The amount of guttation fluid was high when the plant was metabolically particularly active, i.e. when leafing out and reproducing, but it continued for pretty much the life of the leaf (Urbaneja-Bernat et al. 2020). Fruits of Ericaceae are a food source for the ascomycete Monilinia (polyphyletic - ascomycete-Sclerotiniaceae), also found on Rosaceae (Holst-Jensen et al. 1997). Thus M. vaccinii-corymbosi causes "mummy berry disease" in V. corymbosum. Conidia are produced in a sugary matrix on the infected leaves which also reflect UV light like some flowers, also cinnamyl alcohol and cinnamic aldehyde are produced, and these are attractive to bees in particular; the bees move the conidiospores to the flowers, the fungus overwintering in the mummified berries (McArt et al. 2016).
And on yet another subject, callunene, 4-(3-oxobut-1-enylidene)-3,5,5-trimethylcyclohex-2-en1-one, is found in the nectar of the important nectar plant Calluna vulgaris and can prevent the establishment of the trypanosome pest Crithridia bombi in its pollinator Bombus terrestris, but it cannot cure an infestation; it causes the flagellum by which the parasite attaches to the bee gut, to drop off (H. Koch et al 2019). For Helianthus, Bombus and Crithidia, see Ericaceae..
Vegetative Variation. Variation in leaf morphology in Ericaceae is extensive. For instance, linear leaves are found in Diplycosia (= Gaultheria), Rhododendron, Empetrum and relatives, Killipiella (= Sphyrospermum), Agarista, etc.. "Ericoid leaves" - typically scleromorphic, narrow (less than ca 5 mm wide), more or less linear, and often with apparently recurved margins - are quite common, as in Erica itself. Within the small genus Cassiope leaf morphologies vary from flat and more or less linear, or again with apparently strongly recurved margins, or like the finger of a glove (hypoascidiate), to peltate (Stevens 1970), but the different morphologies do not map simply on to the recent phylogeny of the genus, where divergence is dated to ca 17 Ma (Gillespie & Kron 2013; Hou et al. 2015). Indeed, the ericoid leaves of Erica, Empetrum, etc., are not revolute, rather, the revolute appearance is the result of outgrowths from the abaxial side of the lamina (e.g. Hagerup 1946, 1953; Leinfellner 1959). Leaves of Epacridoideae do not have recurved margins (and only rarely leaf teeth), but are otherwise scleromorphic and often more or less ericoid.
Genes & Genomes. Y. Wang et al. (2020) detected genome duplications dated at ca 9.0, 43.4 and 116.9 Ma in cultivated blueberries; the last is probably the γ hexaploidy event. A genome duplication may include part of Rhododendron to all of Ericoideae (Larson et al. 2019/2020); see also Diaz-Garcia et al. (2021) for variously-named duplications.
Atkinson et al. (1994) looked at cytological variation in tropical Vaccinieae, and Schlautman et al. (2017) found extensive colinearity between the genomes of Vaccinium macrocarpum and diploid species of section Cyanococcus which diverged ca 10.4 ma. Stace et al. (1997: c.f. tree) discussed cytological evolution in Epacridoideae; they thought x = 13 might be quite basal in Ericaceae. Khan et al. (2021) looked at variation in the size of the nuclear genome in Rhododendron.
Fajardo et al. (2013) found a number of inversions in the long single copy area of the plastome of Vaccinium macrocarpon, and the genome is very large; more sampling in the Ericaceae-Ericales area in particular is needed to evaluate the significance of this. There were also extensive changes in the plastome of Arbutus unedo, including gene losses, tandem repeats, etc., and some of the changes there were the same as in V. macrocarpon, thus even in these autotrophic Ericaceae the small single copy region, at a little over 3,000 bp, is very short (Martínez-Alberola et al. 2013). Collinearity/synteny of the plastome of perhaps all Ericaceae when compared to that of other Ericales, etc., is low (Martínez-Alberola et al. 2013; Braukmann et al. 2017).
For the loss of chloroplast genes in mycoheterotrophic Ericaceae, see Braukmann and Stefanovic (2012), Logacheva et al. (2016) and Braukmann et al. (2017). The genome in such plants is small and the inverted repeat is lost in six genera, including Pterospora, or it is very small, as in Monotropa, although it may have been secondarily regained there (Braukmann et al. 2017). For plastid evolution in parasitic flowering plants in general, see also Wicke and Naumann (2018).
Economic Importance. In addition to taxa grown for their flowers, most notably Rhododendron, a number of species of Vaccinium are grown for their "berries" (Edger et al. 2022). These include species of section Cyanococcus such as highbush and lowbush blueberies (V. corymbosum and V. angustifolium respectively), also cranberry (V. macrocarpon), bilberry (V. myrtillus) and lingonberry (V. vitis-idaea), although the last two are usually not cultivated. Bradshaw et al. (2025) noted that the powdery mildew Erysiphe vaccinii (it originated on E. North American Vaccinium species) had quite recently become globally distributed, largely as the asexual stage, and posed a serious threat to the blueberry industry.
Chemistry, Morphology, etc.. For a survey of flavonoids and simple phenols, see Harborne and Williams (1973); note that ellagic acid has been found in the pollen of some European Ericoideae (Ferreres et al. 1996) even though the subfamily is generally considered to lack the tannin. For hydroxycinnamates, diverse and showing some correlation with taxonomy in Rhododendron, see N. Shrestha et al. (2017).
Rays in some epacrids may be very low and narrow (Carlquist 2015b). The best developed pit membrane remnants in Ericaceae occur in Enkianthus. They are more poorly developed in other genera, but are well developed in other families in this part of Ericales (Carlquist & Schneider 2005) - a plesiomorphy? Sylleptic branching is at best uncommon (Keller 1994). The leaf midrib of Cassiopoideae may not have associated ("pericyclic") fibres (Kron et al. 2002b), but details of the distribution of this feature are not clear. A group of genera around Lyonia can be characterized by having a lignified epidermis, bands of fibres in the secondary phloem (i.e. stratified phloem), anomocytic stomata, etc.. Variation in indumentum in the family is considerable (e.g. Seithe 1960: Rhododendron; Hubrecht & Bourguignon 2016: U.V. fluorescence Rhododendron scales).
It can be difficult to interpret the floral morphology of Monotropoideae, as with other myco-heterotrophic and parasitic groups, especially of Monotropeae. For instance, members of the outer perianth whorl of the flowers of Monotropoideae such as Monotropa itself have small buds in their axils and are interpreted as being modified bracts by Freudenstein and Brown (2015). Seeds and embryos are usually very small, and Monotropa uniflora has a two-celled embryo (Olson 1991), and much smaller than this you cannot get (except in Anemone); There are about nine endosperm cells in M. hypopitys, the shoot apex aborts during germination and inflorescence shoots develop from buds that come from the roots (Francke 1935: embryo huge - 3 cells; Kaplan 2022).
There are a number of reports of sepals with but a single trace in Ericaceae, although the trace may soon divide into three (Chou 1953). Monosymmetric flowers in Rhodoreae have inverted symmetry, the median sepal being abaxial. Any speckling or colour marking of the corolla occurs on the adaxial petal(s) - as in Lupinus, also with inverted flowers. Tripetaleia (= Elliottia, Ericoideae-Phyllodoceae) apparently has three petals, but very early in develpopment there are five primordia, four of which fuse in pairs, evident in the vascular supply; note that the androecium and gynoecium, but not the calyx, are also trimerous at maturity, but apparently without there being such an obvious reduction in primordium number (Nishino et al. 1983, 1988). Obdiplostemony is reported from some Ericaceae (Ronse De Craene & Bull-Hereñu 2016). Anther pores form in two ways. In one, resorbtion tissue, crystals of uncertain nature develop in the cells and the walls break down; this process is similar to what goes on in the granular pouches on the backs of the anthers of some species. In the other, collapse tissue, the cells vacuolate, flatten, and break down, but no crystals form (Hermann & Palser 2017). For the development of the distinctive pollen of many Epacridoideae in which only a single cell of the tetrad persists, see Furness (2009) and Lemson (2011). A common surface morphology of pollen grains in Ericaceae is faintly cerebellar, although there are some notable exceptions, as in Vaccinium japonicum - indeed, pollen is somewhat more variable than one perhaps might have thought (Sarwar et al. 2006: Vaccinium, 2008: Arbutoideae; Sarwar & Takahashi 2006a: Vaccinioideae excl. Vaccinieae, 2006b: Enkianthus, 2007: Vaccinieae, 2009: Cassiopoideae and Harrimanelloideae, 2014; Erica; Lu et al. 2009: Gaultheria and relatives). Tetrads are found almost throughout the family, but in Epacridoideae-Styphelieae they have secondarily become monads (= pseudomonads) by the abortion - programmed cell death - of three of the four products of meiosis; other kinds of modified tetrads also occur in Styphelieae (Smith-White 1959; Furness 2009: see also Cyperaceae). The pollen morphologies of these pseudomonads are quite unlike single pollen grains elsewhere in the family (see Puente-Lelièvre 2015), whether plesiomorphic (Enkianthus) or derived (Erica). Carpels are opposite the calyx in Vaccinium, Dracophyllum and Monotropa (Schnizlein 1843-1870: fams 160, 161). For more on the distinctive embryology of Ericaceae, also floral anatomy here, see Stushnoff and Palser (1970 and references).
For general information on the family, see Kron et al. (2002b) and Stevens et al. (2004a), for Monotropoideae, see Copeland (1941), Wallace (1975) and papers in Merckx (2013), especially Merckx et al. (2013a), Rhododendroideae, see Copeland (1943), Pyroloideae, see Knudsen and Olesen (1993), Oligarrheneae, see Albrecht et al. (2010), Dracophyllum, see Venter (2021), Erica, see Oliver (1991: Ericoideae, 2000: important) and Oliver and Oliver (2005) and references, Rhododendron, see Milne (2017), Pterospora, see Bakshi (1959), and for New World taxa, see Luteyn (2000, 2002). For oligosaccharide storage, Fouquieriaceae, Diapensiaceae, and Cyrillaceae (and Lennoaceae) also sampled, see Pollard and Amuti (1981); for protein crystals in the nucleus, see Speta (1977, 1979), for wood anatomy of Cassiope and Harrimanella, see Wallace (1986), of Epacridoideae, see Lens et al. (2003) and of superior-ovaried Vaccinoideae, see Lens et al. (2004a); for nodal anatomy of Rhododendron, see Philipson and Philipson (1968); for epidermal variation in Gaultherieae, see Y.-H. Wang et al. (2015), for epicuticular waxes in Epacridoideae, see Weiller et al. (1994), and for pseudotori, see Rabaey et al. (2006); for vegetative features of Epacridoideae, see Jordan et al. (2010); for venation patterns in some Neotropical Vaccinieae, see Pedraza-Peñalosa et al. (2013); for floral morphology of Empetrum, see Vislobokov et al. (2012), of Erica and many other Ericaceae, Palser and Murty (1967, also references to earlier papers in this series), of Gaultheria, Chou (1953), for pollen, see Skvarla et al. (1978) and Waha (1984), both viscin threads, Takahashi (1987: Monotropoideae), X.-P. Zhang et al. (2002), Sawara (2007: comprehensive), Sawara and Shameem (2008 - Arbutoideae), Wronska-Pilarek et al. (2018: Erica) and Mitra et al. (2019: Diplarche), for nectaries, see Erbar (2014), for integument thickness, see Samuelsson (1913), for embryology of Vaccinieae, see Venkateswarlu and Maheshwari Devi (1973), for seed anatomy, see Peltrisot (1904), for external seed morphology of Gaultherieae, see Lu et al. (2010a) and that of Erica - quite variable - see Szkudlarz (2010), and for seeds and seedlings of Rhododendron, see Hedegaard (1980).
Phylogeny. Early studies are summarized by Kron et al. (2002b). The structure of the tree immediately above Enkianthoideae was initially labile with Pyroloideae, Monotropoideae and Arbutoideae variously arranged and often forming a basal grade, although support values were low (e.g. Cullings 1994; Cullings & Hileman 1997: [Monotropoideae + Arbutoideae]). Freudenstein et al. (2010) in a comprehensive phylogenetic study of the family suggested the relationships [Enkianthoideae [[Pyroloideae [Arbutoideae + Monotropoideae]] [The Rest]]] (also Z.-Y. Liu et al. 2011, 2014; Hardy & Cook 2012; Braukmann & Stefanovic 2012: PHYA, see below; esp. Freudenstein et al. 2016a; Lallemand et al. 2016; Lam et al. 2018: some caveats); these relationships are followed here. However, in early versions of this site (pre-August 2010), Monotropoideae (including Pyroloideae) and Arbutoideae were successively sister to the remainder of the family (other than Enkianthus), while Brundrett (1994) found a very different set of relationships, mycoheterotrophy apparently having evolved several times, Feldenkreis et al. (2011) suggested the relationships [Enkianthoideae [Pyroloideae [[Monotropoideae + Arbutoideae] [The Rest]]]], Schwery et al. (2014) found Arbutoideae to be embedded in a paraphyletic Monotropoideae, while in Rose et al. (2018) [Enkianthoideae [[Monotropoideae + Arbutoideae] [Pyroloideae [The Rest]]]] were the relationships, although the position of Pyroloideae had little support; this topology was recovered, and with very strong support, in the full chondrome analyses of Q. Lin et al. (2022) - note that Freudenstein et al. (2016a) had recovered this topology in their plastid and mitochondrial analyses, so it seems to be an organellar phylogeny. In Braukmann and Stefanovic (2012) Pterospora, on a very long branch, was sister to all other Ericaceae except Enkianthus, and subfamilial relationships in Z.-D. Chen et al. (2016) are also rather different than those above. As might be expected, there was little support for most of the branches in a morphological analysis using 82 morphological characters although most of the major groupings appeared in the strict concensus tree, but Vaccinioideae were strongly paraphyletic and Cassiope and Harrimanella were embedded in Rhododendroideae (Kron et al. 2002b). Anderberg (1993) had recovered a paraphyletic Monotropoideae with Pyroloideae embedded it in an earlier morphological analysis.
Monotropoideae. Cullings (1994) examined relationships within Monotropoideae, and although the subfamily as a whole had poor support, a clade [Sarcodes [Monotropa hypopithys<]/i> + Pterospora]] was sister to a second clade containing the rest of the subfamily (see also Cullings & Hileman 1997), the two being well supported; Monotropsis ordata (sic) was well separate from the rest of the Pyroloideae/Monotropoideae. However, Cullings (2000) found that additional samples of Monotropa hypopithys grouped with this second clade. All rather confusing, but in the analyses of Bidartondo and Bruns (2001) [Sarcodes + Pterospora] alone were consistently sister to all other Monotropoideae, even if relationships within the latter differed somewhat depending on whether nuclear or chloroplast markers were analyzed - e.g. M. hypopithys was strongly supported as being monophyletic in the former analyses but was polyphyletic in the latter. Bidartondo and Bruns (2001) suggested that problems with earlier studies may have resulted from misidentifications, both of fungi and their hosts. Relationships between the three genera included in the chondrome analysis of Q. Lin et al. (2022b) were [Pterospora [Allotropa + Hypopitys]]. Freudenstein and Broe (2024: 102 nucleear genes from 9/11 genera, also plastid coding loci, etc.) recovered groupings like [[Sarcodes + Pterospora] {Pleuricospora ...]], [Allotropa + Hemitomes] (but not in all analyses) and [Monotropsis [Eremotropa + Monotropa + Monotropastrum]], Hypopitys and Monotropa are distinct, and previous ambiguities in rps2 relationships were explained by the existence of plastid and mitochondrial paralogs, and so on.
Pyroloideae. For the phylogeny of Pyrola and its relatives, see Freudenstein (1999) and Z.-W. Liu et al. (2011, 2014). Z.-W. Liu et al. (2011) noted that the position of Orthilia was unstable, and there was a suggestion that allopolyploidy might be involved in its origin. Matsuda et al. (2012) found the well supported relationships of [[Orthilia + Pyrola] [Moneses + Chimaphila]], while relationships in Z.-W. Liu et al. (2014) are [Orthilia [Pyrola [Moneses + Chimaphila]]]; the former set of relationships is preferred here (see also Lallemand et al. 2016; Q. Lin et al. 2022: Orthilia not included).
Arbutoideae. Arbutus sometimes appears to be paraphyletic with respect to the other genera of Arbutoideae (Hileman et al. 2001; see also Kron et al. 2002b), but broader sampling with the ITS gene yields a topology compatible with conventional delimitations of genera, in particular, Arctous is not sister to Arctostaphylos (Greg Wahlert, pers. comm.). For relationships within Arctostaphylos s. str., see Wahlert et al. (2009); the genus may be monophyletic, but no taxa outside the subfamily were included.
Cassiopoideae. Relationships within Cassiope are discussed by Gillespie and Kron (2013: ?hybridization) and Hou et al. (2015).
Ericoideae. Gillespie and Kron (2010: four chloroplast, two nuclear markers) studied relationships across Ericoideae and found i.a. that the distinctive Himalayan Diplarche, previously of uncertain relationships, was sister to Empetreae (see also Z.-D. Chen et al. 2016), and they thought that it should be included there, and the Guyanan Ledothamnus was sister to the northeast Asian Bryanthus. However, separate analyses of the two nuclear markers placed Diplarche as sister to Corema alone (Gillespie & Kron 2010), and relationships along the spine of Ericoideae were poorly supported even in the combined analysis. An earlier study using two chloroplast markers had linked Diplarche with Rhododendron, etc. (Kron et al. 2002b), and Craven (2011) even made combinations of Diplarche in Rhododendron. Kron et al. (2002b) found that the position of Bejaria was uncertain, although they elected to include it in Bryantheae, although later it was found to be sister to all other Phyllodoceae, Elliottia plus the rest, if with low maximum parsimony support (Gillespie & Kron 2010). All a little confusing. For the phylogeny of Ericoideae-Ericeae and the circumscription of Erica itself, see e.g. Oliver (1994), McGuire and Kron (2005) and Pirie et al. (2011); Daboecia is sister to the rest of the tribe and the African species of Erica are probably monophyletic. The circumscription of Rhododendron and relationships within it have been the subjects of much recent work (Kurashige et al. 2001; Gao et al. 2002; Kron 2003: limits of genus, sections; G. K. Brown 2003; Milne 2004: subsection Pontica paraphyletic, includes subgenus Hymenanthes; Brown et al. 2006a, c, d: section Vireya = sect. Schistanthe; de Riek et al. 2008: Therorhodion sister to Rhododendron; Craven et al. 2008; especially Goetsch et al. 2005, 2011). Genera like Azalea, Ledum and Menziesia are now all included in Rhododendron, and the erstwhile Therorhodion is consistently found to be sister to the rest of the genus (e.g. N. Shrestha et al. 2018; Xia et al. 2021; Khan et al. 2021; Mo et al. 2022). Khan et al. (2021: two chloroplast and two nuclear genes, separate and concatenated analyses, 260 species) found that the larger subgenera held, but taxa in the smaller subgenera were not to be placed so easily. Thus the positions of taxa like R. vaseyi (section Rhodora), R. semibarbatum, R. albiflorum (in some classifications separate subgenera, Mumeazalea and Candidastrum respectively), R. tomentosum (= Ledum that was) were uncertain. Xia et al. (2021: 3437 orthologous nuclear genes, 200 species of R., ca 1/5 the genus) suggested that Menziesia (no surprise) and subg. Choniastrum (somewhat surprising) should be merged into subg. Tsutsusi. Soza et al. (2022) focused on relationships within section Schistanthe, all members of which are diploid (n = 13), and found hybridization/introgression within and between all subsections of any size, and even between subsection Pseudovireya, perhaps sister to the rest of the section, and a Rhododendron outside the section; Mo et al. (2022: esp. Fig. 4, 161 spp., plastomes) also commented on the possibility of hybridization events here. General relationships here may be subsections [Pseudovireya [Discovireya [Malayovireya + Euvireya]]], four clades with a generally west to east distribution (Soza et al. 2022); sections earlier recognised by Sleumer (within Vireya) were found not to be monophyletic by G. Khan et al. (2024). Y. Ma et al. (2022) suggested that there had been widespread hybridization in subgenus Hymenanthes, and they found extensive discordance between nuclear and plastome SNP trees there. Y. Shen et al. (2024) found the relationships in the tree above; subgenera Tsutsusi and Azaleastrum were in the same clade.
Epacridoideae, For the phylogeny of Epacridoideae, see Powell et al. (1996), Crayn and Quinn (2000) and K. A. Johnson et al. (2012). Prionoteae and Archerieae are successively sister to remaining Epacridoideae, and this is consistent with morphology (see above). Wagstaff et al. (2010) looked at relationships within the distinctive Richeeae, however, relationships between this tribe and Cosmelieae remain uncertain (Johnson et al. 2012). Relationships in Styphelieae are being disentangled (e.g. Powell et al. 1997: morphology), Cherry et al. 2001; Quinn et al. 2003, 2005; Puente-Lelièvre et al. 2012). In a five-gene (two compartments)-207 taxon study with a focus on Styphelia, it and Leucopogon popped up all over the place - with obvious taxonomic implications (Puente-Lelièvre et al. 2015). Epacris has turned out to be paraphyletic (Quinn et al. 2015).
Vaccinioideae. Tribal relationships are somewhat unclear here. Oxydendrum was found to be sister to the rest of the subfamily by Kron et al. (2002b), while Lyonieae had that position in Schwery et al. (2014). Z.-D. Chen et al. (2016) recovered the relationships [Lyonieae [[Gaultherieae + Andromedeae] Vaccinieae]] among the Chinese taxa (see also Kron et al. 2002b; Rose et al. 2018).
Gaultherieae. Within Gaultheria s.l. the epiphytic Diplycosia with some 100 species and Tepuia (Powell & Kron 2001, 2002; Bush & Kron 2008; Bush et al. 2006, esp. 2009: fruit and inflorescence characters; Bush & Kron 2008; Fritsch et al. 2011; esp. Lu et al. 2019a: see fruit colour, esp. 2019b) may form a clade (clade 1) along with a few species of Gaultheria (including section Gaultheria), however, Tepuia in particular may be separate and make up part of a basic tritomy for the whole clade in some analyses. The majority of Gaultheria forms a clade, clade 2, sister to Tepuia, however, the position of G. procumbens itself was unclear, and the sections and series previously recognized in Gaultheria did not hold up (e.g. Fritsch et al. 2011; see also Fritsch et al. 2020b). Interestingly, the Leucothoides group of Gaultheria, from southeast Asia, has two copies of the waxy gene, and one copy puts it with clade 1 and the other with clade 2 (Lu et al. 2019b). Y.-H. Wang et al. (2015) carried out a morphological analysis of numerous leaf epidermal characters of Gaultherieae and compared the groupings obtained with those from other studies. There may be a number of cryptic species in the high-altitude representatives of the genus (Lu et al 2010a); Cheng et al. (2024) looked at relationships in species of Gaultheria ser. Trichophyllae that grow on sky islands in the Himalaya-Hengduan Mountain region.
Vaccinieae. Outlines of relationships in the tropical inferior-ovaried Vaccinieae are slowly developing (Kron et al. 2002a) and for the most part they cut across the limits of most genera of any size; these are based on floral characters, often variants of a bird pollination syndrome. However, a Vaccinium-type flower, that is, small, ± urceolate, appears to be plesiomorphic in the whole clade and Vaccinium itself is very paraphyletic (see also Powell & Kron 2002, 2003; Pedraza-Peñalosa 2009). In particular, the "Tethyan" Vaccinium section Hemimyrtillus, from the Mediterranean area, etc., may be sister to other Vaccinieae, although there is currently only weakish support for this position (Powell & Kron 2002). Crowl et al. (2022) looked at the relationships of 36 diploid individuals of section Cyanococcus using 323 nuclear loci (from the Angiosperms353 data set) and found a fair amount of resolution; there was also hybridization (and relationships between the three outgroups varied). In Southeast Asia the Agapetes s. str. clade, with 90 or more species centred in the S.W. China-the Himalayan region, will probably need to be extended to include some 250+ species of Vaccinium, all having superficial stem phellogen and a falsely 10-locular ovary, both probably derived features, and forming a single clade (Powell & Kron 2002; Tsutumi 2011; Z.-D. Chen et al. 2016: Agapetes, Vaccinium sections Bracteatum and Rigiolepis in the same immediate clade; Ghandforoush & Kron 2016). J. Wang et al. (2025) looked at the relationships suggested by plastome variation of three species of Agapetes and six species of Vaccinium; outgroups included Gaultheria, and found that although the species of Agapetes formed a clade, it was embedded in Vaccinium, while in an ITS analysis (8 spp. Agapetes, 7 spp. Vaccinium), the polyphyly of Agapetes was extreme, and even the three species of Agapetes in the first analysis did not form a clade. Note that the plastomes of the species in the two "genera" - simple sequence repeats, etc. - were quite similar to one another, and different from those of the other species in the study (Wang et al. 2025). New Guinean Dimorphanthera is sister to Paphia, and both have deep-seated pericyclic stem phellogen and are primarily New Guinean; Paphia used to be included in Agapetes, but the two are not immediately related. Pedraza-Peñalosa (2010) explored the limits of Disterigma and Pedraza-Peñalosa et al. (2015) those of Colombian Vaccinieae in general, and in the latter, extensive polyphyly in the larger genera was found.
Recently Becker et al. (2024: 256 low copy nuclear genes, 210 terminals) looked at relationships in the tribe, sampling 30/35 genera and within Vaccinium, 25/29 sections. Results suggest extensive para/polyphyly, both of Vaccinium (see "sect." below - only some included) and of several other genera. An outline of the overall topology is [[V. sect. Neurodesia, etc.] [[Gaylussacia + sect. Hemimyrtillus] [[Costera [[sect. Vaccinium + sect. Oxycoccus etc.] [Old World clade/genera [Mesoamerican clade/genera + Andean clade/genera]]]] [[sect. Pyxothamnus [sect. Vitis-idaea [sect. Hemimyrtillus + sect. Vaccinium]]] [sect. Cyanococcus [sect. Batodendron [lots of sections, inc. Agapetes serpens (only the one sp.!)]]]]]]. Note that this topology (ibid., Fig. 2) is based on a ML concatenated analysis, and the ASTRAL-III species tree showed substantial differences in places, e.g., in the position of Costera. Becker et al. (2025: plastid and Angiosperms353 data) looked at relationships in Vaccinium section Myrtillus with a focus on the Hawaiian species. Relationships obtained varied according to whether the data were plastid or nuclear, etc.. J. Wang et al. (2025) looked at the plastomes of three species of Agapetes and some species of Vaccinium - Agapetes were sister to three species of Vaccinium - from North America, however, in an ITS analysis involving 8 species of Agapetes and 7 species of Vaccinium (the latter were all from southeast Asia) species from the two groups were intermixed, as perhaps might be expected. Interestingly, Wang et al. (2025) found that Agapetes and non-southeast Asian Vaccinium (Vaccinium from southeast Asia were not included) both had numerous simple sequence repeats (SSRs), although mononucleotide SSRs were relatively uncommon. The bottom line? A global study of Vaccinieae with good sampling - no easy task - using nuclear genes and ideally compared with plastid data is needed - as Becker et al. (2024) note, only then will the needed nomenclatural changes become possible.
For additional information on relationships, see Anderberg (1993), Cullings (2000), Judd and Kron (1993), Kron and Chase (1993), Kron et al. (1999a, b) and Crayn et al. (1998).
Classification. The infrafamilial classification outlined by Kron et al. (2002b) is largely followed here; Gillespie and Kron (2010) adjusted tribal limits in Ericoideae.
Genera in Monotropoideae are becoming ever more finely split (see e.g. Q.-R. Zhao et al. 2018). Argent (2006) provided an account of species of Rhododendron subgenus Vireya (= section Schistanthe); Craven et al. (2008, esp. 2011) listed the subsections that it includes. Azalea, Ledum, Menziesia, Tsusiophyllum, and even Diplarche (Craven 2011, however, see above: to be placed in Empetreae?) are all to be included, indeed, Menziesia hybridizes with related species of Rhododendron (de Riek et al. 2008). However, although Khan et al. (2021) found that a number of taxa recognized by Goetsch et al. (2008; see also M. Shrestha et al. 2018) were monophyletic, the exact relationships of quite a number of taxa varied depending on data analyzed, etc., suggesting that changes may have to be made in more complete analyses. Erica has been expanded to include the wind-pollinated Philippia and several small segregate genera (e.g. Oliver 2000), q.v. for a sectional classification of Erica. Spethmann (1987) provided a comprehensive classification of Rhododenron which integrated i.a. the phytochemical work that he and others had carried out on the genus, as well as seedling leaf venation, nodal anatomy, etc.; Goetsch et al. (2005) provide a molecular framework for the reclassification of the genus. Generic limits in Styphelieae and some other Epacridoideae are difficult, but Quinn et al. (2005) and Albrecht et al. (2010) suggest some realignments - see e.g. Puente-Lelièvre et al. (2015) for a useful discussion about the merits and demerits of having broad or narrow generic limits. As Puente-Lelièvre et al. (2015) note, either the limits of Leucopogon will have to be restricted or those of Styphelia expanded to include much of the tribe, and they prefer the latter option, so whereas most of the diversity in Styphelieae used to be in Leucopogon, it is now in Styphelia... (see also Taaffe et al. 2001; K. A. Johnson et al. 2012; Hislop & Puente-Lelièvre 2017; Crayn et al. 2020). For subgenera in Dracophyllum, see Venter (2021).
There are yet other substantial changes needed, particularly to generic limits in Vaccinioideae. Gaultheria is to include Pernettya, Diplycosia as well as Tepuia (or most of Gaultheria would have to be called Pernettya...); Kron et al. (2020) provide the needed combinations and some sectional names. Generic limits in Vaccinieae in particular are in a major mess (Powell & Kron 2003; Pedraza-Peñalosa 2009, Pedraza-Peñalosa et al. 2015). In Southeast Asia it is likely that the Agapetes clade, with a superficial cork cambium, also includes some 250+ species of Vaccinium (including Rigiolepis - Argent 2019) plus 110 or more species of Agapetes s. str.; this clade includes a number of epiphytic taxa, some with swollen stem bases, taxa with a mucilaginous testa, green embryos, etc.. Dimorphanthera is sister to Paphia, however, if the Paphia clade really does include taxa of the old Vaccinium sect. Pachyantha, merging of Paphia with Dimorphanthera might be best... (c.f. Stevens 2004). Indeed, Vaccinium itself is pretty wildly paraphyletic. However, Vander Kloet and Dickinson (2009) provided a infrageneric classification for "Vaccinium" recognising thirty sections, on the other hand, Argent (2019) recently recognised Rigiolepis, suggesting that it was separate from other Malesian Vaccinium, while Danet (2023) described V. superum, from New Guinea, with a superior ovary and that he tentatively placed in section Nesococcus (included in section Bracteata by Sleumer 1967a)... Given the relationships of "Vaccinium" to the rest of the Vaccinieae, it is probable that there will be numerous nomenclatural changes in Vaccinium and other genera in the tribe as a whole, but these must await a comprehensive phylogenetic analysis; it is likely that the 00s of Indo-Malesian Vaccinieae with pseudo 10-locular ovaries will all end up together (so including species like V. paradoxum, which has a largely superior ovary (Tamayo & Fritsch 2022).
Previous Relationships. Ericaceae here are basically the Ericales of Cronquist and Takhtajan. However, even then there were suggestions that relationships might be more entwined than the separate families they recognized suggested. Thus characters like rust preferences (Savile 1979b) had linked the wind-pollinated and florally very distinctive Empetraceae with Ericaceae-Ericoideae in particular, furthermore, both Rhodoreae and Empetreae have the flavonoid gossypetin - all the taxa in this sentence are in Ericaceae-Ericoideae above.
Botanical Trivia. "Mad honey" disease is the result of eating honey containing the neurotoxin grayanotoxin, and such honey (the grayanotoxin-including nectar came from Rhododendron) temporarily incapacitated some of Xenophon's troops in Asia Minor; the effects of grayanotoxin are only transitory (S. A. Jansen et al. 2012).
){kind=link}
![Fruits.]](javascript:showImage('http://florawww.eeb.uconn.edu/images/byspecies/Marcgravia_rectiflora02.JPG',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![Flowers.]](javascript:showImage('http://www.csdl.tamu.edu/FLORA/BigBend/BB0315.jpg',600,500)){kind=link}
![[Photo - Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-03958_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![[Photo - Flower]](javascript:showImage('http://www.botany.hawaii.edu/faculty/carr/images/bar_asi_cu.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-05130_001.jpg',600,500)){kind=link}
![Fruit].](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-03920_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![[Photo - Habit © S. Hsiao.]](javascript:showImage('http://www.parasiticplants.siu.edu//Mitrastemonaceae/images/Mitrastemma.habit.JPEG',600,500)){kind=link}
![[Photo - Collection.]](javascript:showImage('http://www.botany.hawaii.edu/faculty/carr/images/cam_sp_fl.jpg',600,500)){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04062_001.jpg',600,500)){kind=link}
![[Photo - Symplocos chinensis Flowers.]](javascript:showImage('http://www.pharmakobotanik.de/systematik/7_bilder/yamasaki/yamas208.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-03938_001.jpg',600,500)){kind=link}
){kind=link}
![[Photo - Roridula Flower © M. Schmidt.]](javascript:showImage('http://www2.labs.agilent.com/botany/cp/pictures/roridula/pics/wanze3.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
![[Photo - Cliftonia Habit]](javascript:showImage('http://www.csdl.tamu.edu/FLORA/fa04/fa04031.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
![Pyrola Flower.]](javascript:showImage('http://www.botany.hawaii.edu/faculty/carr/images/pyr_sp_cu.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![Inflorescence.]](javascript:showImage('http://www.botany.hawaii.edu/faculty/carr/images/arc_sp_202.jpg',600,500)){kind=link}
![[Photo - Habit.]](javascript:showImage('http://www.botany.hawaii.edu/faculty/carr/images/cas_tet_hab.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}