 Garden Home |
Research HomeGarden Home |
Research Home
Garden Home |
Research HomeGarden Home |
Research Home


 |

|
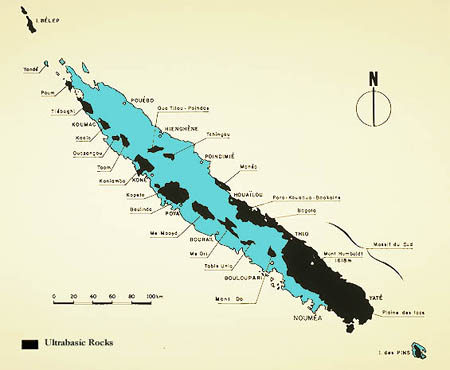
Many authors have suggested that New Caledonia's ultrabasic substrates have played a central role in the evolution and diversification of the island's flora (e.g., Holloway, 1979; Jaffré, 1980; Thorne, 1965; Virot, 1956). Thorne (1965) hypothesized that the ultrabasics allowed many older Australasian groups to survive in a sort of local, ecological refugium. He argued that by adapting to the ultrabasics shortly after their deposition in the Paleocene and Eocene, many primitive groups were able to persist on New Caledonia because these substrates provided a degree of ecological separation and protection from more competitive rain forest plants that continually arrived from other tropical areas.
The flora on New Caledonia's ultrabasic substrates (primarily in the moist evergreen forests and maquis) is rich in species and has a very high degree of endemism (90.6%; see above). At the same time, very few of the island's non-endemics are found on ultrabasics: of the 740 native species shared with other areas, only 173 (23.4 %) occur on ultrabasics, while 587 are found on other substrates, with only 20 of these occurring on both ultrabasics and non-ultrabasics (data derived from Morat, 1993).
Jaffré et al. (1987) presented a series of ideas on the changes that are thought to have accompanied the establishment of New Caledonia's ultrabasics, and the processes by which the modern flora occurring on these substrates was established.
1. Some species present in the original, pre-Eocene flora must have been eliminated because they were not able to adapt to the extreme conditions of the ultrabasics. Other plants were able to survive, however, perhaps in forest islands on non-ultrabasic substrates that are thought to have persisted in areas that were not effected by the processes of peridotite deposition.
2. Components of the original flora were able to occupy and adapt to the highly selective conditions of these substrates in several ways:


Examples of species whose ancestors probably colonized newly formed ultrabasic substrates. From left to right: Grevillea meissneri (Proteaceae); Hibbertia baudouinii (Dilleniaceae).




Examples of sclerophyllous groups that have speciated on New Caledonia. From left to right: Metrosideros tetrastichus (Myrtaceae) on summit of Mt. Humboldt; Cunonia macrophylla (Cunoniaceae) on Mt. Mou; Dracophyllum verticillatum (Epacridaceae) on Mt. Mou; Stenocarpus sp. (Proteaceae) on the Ile des Pins
3. The initial vegetation occupying New Caledonia’s ultrabasic substrates must have been very depauperate, and these areas thus presented ecological conditions favorable for speciation among the components of the original flora that were able to colonize and survive in them. Jaffré et al. (1987) concluded that a new wave of diversification and adaptive radiation gave rise to a large neo-endemic component of the flora, which today accounts for much of the species-level diversity seen on the ultrabasics, especially in maquis vegetation, and also in the moist evergreen forests. Speciation was particularly active among sclerophyllous groups that are pre-adapted to poor soils (Beadle, 1966)(e.g., many Myrtaceae, Cunoniaceae, Epacridaceae, Proteaceae, etc.), and resulted in the evolution of taxa that are able to limit the uptake of Ni or Mg, or to tolerate high concentrations of heavy metals (Brooks et al., 1974, 1981; Jaffré, 1977a, b, 1979; Jaffré and Schmid, 1974; Jaffré et al., 1976, 1979a, b; Kersten et al., 1979; Lee et al., 1977b).
Species and entire groups adapted to ultrabasic substrates have long benefited from some protection against foreign elements reaching New Caledonia (Holloway, 1979; Jaffré et al., 1987; Morat, 1993; Morat et al., 1984, 1986). However, it is not clear that this process provided a special, differential protection to the older floristic elements, in which case one would expect a higher proportion of species belonging to these groups (e.g., gymnosperms and primitive angiosperm families) on ultrabasics than on other substrates, where competition from more recently arrived elements is surely greater. This can be tested by examining the substrate preferences of species in primitive groups.
Each of the three main gymnosperm families (Araucariaceae, Podocarpaceae, and Cupressaceae) has more species occurring on ultrabasics than elsewhere (15:6, 18:8, and 6:1 species, respectively), and the same is true for Lauraceae (36:22)(data from Jaffré et al., 1987). However, approximately equal numbers of Winteraceae (11:9) and Annonaceae (8:5) occur on and off of ultrabasics, and the single species of Amborellaceae is limited to non-ultrabasic substrates; equal numbers of Chloranthaceae occur on both kinds of substrate (2:2), while the figures for Monimiaceae (3:8), Atherospermataceae (0:1), and Trimeniaceae (0:1) show more species on non-ultrabasics (Jérémie, 1982).
Many groups that must have reached New Caledonia after it separated from Australia, before deposition of the ultrabasics, show the same distribution between ultrabasic and non-ultrabasic substrates (Jaffré et al., 1987). Thus, while the ultrabasics may have provided a kind of ecological isolation and protection against elements that arrived in post-Eocene times, they do not appear to have done so indifferently with respect to the primitive and more advanced lineages that were then present on New Caledonia.
The groups present in sclerophyllous forests do not appear to have undergone the same kind of intense speciation that produced large numbers of neo-endemics in the moist evergreen forests and maquis. Only a few genera are well represented in the sclerophyllous forests (e.g., Eugenia, 13 species; Diospyros, 11 species; Austromyrtus, 8 species), and species level diversity is much lower than elsewhere -- although 59 of the endemic species are restricted to the sclerophyllous forests, along with one endemic genus of Rubiaceae (Captaincookia). Several groups that are well represented in the moist evergreen forests are completely absent from the sclerophyllous forests, including gymnosperms, Arecaceae, Epacridaceae, Fagaceae, Pandanaceae, and Winteraceae, (Jaffré et al., 1993; Bouchet et al., 1995). This may in part be an artifact of the relictual nature of the remaining stands, and it also reflects the relatively young age of this formation, whose flora comprises elements largely dating from the Miocene and Quaternary (Jaffré et al., 1993).