|
|
Barry E. Hammel
Missouri Botanical Garden, P.O. Box 299, St. Louis,
Missouri 63166-0299 U.S.A.
Novon 9: 349-359. 1999. Published on 15 September 1999. ©
Missouri Botanical Garden 1999.
Abstract. Two new species of Clusiella, one from Costa Rica and Panama (C. isthmensis) and one from Brazil (C. impressinervis), are described, one old species from Colombia (C. cordifolia Cuatrecasas) is newly placed in synonymy, and all eight species in the genus are brought together in a key. a description of the genus and brief descriptions are given, specimens are cited and distributions are mapped for all species. In view of its stipuliform structures, contorted petals, psilate pollen exine, baccate fruits, exarillate seeds, and embryos with well-developed cotyledons, Clusiella's presumed close relationship to Clusia is challenged.
When Planchon and Triana (1860) published Clusiella (based on C. elegans) the genus was considered monotypic and known only from pistillate flowers. It remained that way for nearly 100 years. However, Planchon and Triana appended to their treatment of Clusiella a full description of staminate material of a species now known as C. axillaris (Engler) Cuatrecasas. Planchon and Triana implied that the collections on which that description was based (Spruce 2159 and 2854) might belong to a second species of Clusiella, but they gave it no species name, and presented the description under the heading "Asthotheca, Miers" with the prophetic injuction to see it "without prejudice as to placement among the other Clusioids" (Planchon & Triana, 1860, ser. 4, vol. 14: 254). Although all genera, except this one, are specifically designated as such in their large publication Planchon and Triana did effectively publish the genus. The spelling "Asthotheca," however, must certainly have been a typographical error for Astrotheca; on p. 240 of the same publication they refer to the "genre manuscrit Astrotheca, Miers." Bentham and Hooker (1862) repeated that error when they reduced "Asthotheca" to a section of the genus Clusia. Meanwhile, Engler (1888) published Clusia axillaris Engler based on the same two Spruce collections. Vesque (1892, 1893), using the correct spelling, and aware that "the discovery of its female flowers and of the male flowers of Clusiella might one day permit the combination of these two genera" (Vesque 1892: 16), assigned two species to Astrotheca, A. cuspidata Vesque and A. sulphurea (Poeppig) Vesque. The former was clearly a superfluous epithet because Vesque cited the earlier Clusia axillaris Engler and both Spruce collections. Astrotheca sulfurea is not a Clusiella (Cuatrecasas, 1950). Cuatrecasas (1949, 1950) finally brought Astrotheca (as to A. cuspidata Vesque) to rest in synonymy under Clusiella and described five new species.
Clusiella Planchon & Triana, Ann. Sci. Nat. Bot. Sér. 4. 14: 253. 1860. TYPE: Clusiella elegans Planchon & Triana.
Astrotheca Miers ex Planchon & Triana, Ann. Sci. Nat. Bot. Sér. 4. 14: 254. 1860 (as "Asthotheca," a typographical error). No species indicated, but based on Spruce 2159 & 2854, from Brazil. LECTOTYPE, here designated: Astrotheca cuspidata Vesque (=Clusiella axillaris (Engler) Cuatrecasas).
Clusia subg. Criuva sect. Astrotheca (Miers ex Planchon & Triana) Bentham & Hooker, Gen. Pl. 1: 170. 1862 (as "Asthotheca").
Dioecious, epiphytic shrubs; resin (latex) clear; internodes lenticellate, bearing several pairs of caducous or rarely persistent bud scales usually clustered just above the node (one pair more distant). Minute, interpetiolar stipuliform structures present, these broadly triangular, scarious, drying dark, caducous. Petioles short, somewhat channeled by virtue of the narrowly decurrent leaf blade. Leaves opposite, the blades elliptic, sometimes cordate, ± coriaceous, occasionally glaucous in dried material; major lateral veins often very indistinct until dried, merging to a submarginal vein, the intersecondaries often almost as prominent as the secondaries; translucid (sometimes only with intense light) resin dots and dashes present, occasionally drying dark and surficially visible but then usually only on the lower surface, resin canals sometimes also visible on thin, new leaves. Inflorescences terminal or evicted by the growth of an axillary shoot and then appearing axillary, at only one side of the node, short, 1-several-flowered, ± dichotomously branching cymes with numerous small, decussate bracts dispersed on the rachis. Flowers with 5 small imbricate sepals and 5 larger contorted petals, the petals white (maroon or wine-red), often with yellow or pink markings at the base within. Staminate flowers with the filaments connate for most of their length into a narrow column but free for a short distance apically and forming a ± globose capitulum, the column surrounded by a collar of clavate, resiniferous staminodia or resin globules; anthers ± globose and only slightly wider than the filaments (Figs. 1, 2); pollen spheroidal-oblate, tricolporate, psilate (Fig. 3). Pistillate flowers with the ovary surrounded by a resiniferous collar of staminodes that grade acropetally from resin glands to more clearly stamenlike structures; stigmas sessile, 5 to ca. 20, circular and distinct (when few) or packed into a disk and indistinctly triangular to rectangular (when many); ovules sometimes with a long funicle. Fruit a ± globose, many-seeded berry with about as many locules as stigmas; seeds elongate-ovoid, small (ca. 1 X 0.5 mm) with a clear, gelatinous outer integument attached only at the hilum, the exotegmen pitted (Figs. 4, 5), with sinuous anticlinal cell walls, lignified; endosperm white, embryo with cotyledons ca. 2/5 the length of the seed.
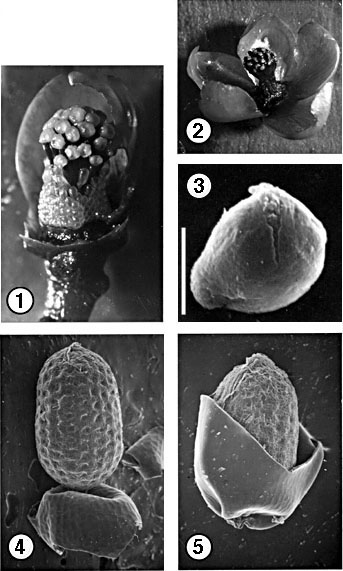
Figures 1--5. Clusiella (1--3, C. elegans Planchon & Triana; Hammel & Kress 11287; 4, 5, C. isthmensis Hammel, Hammel & Trainer 12721). ---1. Staminate bud with petals removed, showing undehisced anthers and staminal column subtended by whorl of resinous staminodia. ---2. Mature staminate flower. ---3. Pollen grain. ---4. Seed with integument laid back to show pitted exotegmen. ---5. Seed with the clear integument partially removed. Scale bar: for Figs. 1, 4 & 5 = 3 mm; for Fig. 2 = 6 mm; for Fig. 3 = 10 µm.
Distribution. Clusiella is restricted to the Neotropics, where species occur in Costa Rica, Panama, Colombia, Venezuela, Ecuador, northern Peru, and northern Brazil (Fig. 6).

Figure 6. Distribution of Clusiella. (Occurrence of C. axilaris with __> pertains to off-map collection in Pará state of Brazil).
Clusiella albiflora Cuatrecasas, Ann. Inst. Biol. Mex. 20: 111. 1949. TYPE: Colombia. Valle del Cauca: Río Yurumanguí, Peña de Candelario, Costa Pacífica, 10 m, Feb. 1944 (stam. fl), Cuatrecasas 16123 (holotype, F; isotype, US).
Bud scales sometimes persistent, to ca. 4 cm long, their scars up to 3.5 cm above the node. Petioles 8--10 mm; leaf blades elliptic, 8--12 x 4--6 cm, the base acute, the apex acuminate to a 1.5--2.5 cm acumen, drying glaucous above, dark tan above and below, lateral veins ca. 1--2(--4) mm apart, merging to a distinct submarginal vein ca. 1--1.5 mm from the margin, the intersecondaries prominent and easily confused with the secondaries. Inflorescences branched several times from above a peduncle ca. 5 mm long, 1-several flowered; flower buds ca. 8 mm diam.; petals ca. 10 x 5 mm. Staminate flowers with the staminal column, including capitulum, very short, ca. 3 mm long. Fruits globose, ca. 10 mm diam., the stigmatic disk with ca. 6 closely packed, ± triangular stigmas, fruit wall rather thick, ca. 0.5 mm.
Distribution. Colombia, from near sea level.
This species is still known only from the type and one paratype. It has longer petioles and closer lateral veins than the other species as well as a very long acuminate leaf tip and a very short staminal column. In leaf shape and color, though not in size, it appears most similar to C. elegans.
Additional specimen examined. COLOMBIA. Cauca: Río Micay, Guayabal, Costa Pacífica, 520 m, Feb. 1943 (fr), Cuatrecasas 14132 (F, US).
Clusiella amplexicaulis Cuatrecasas, Rev. Acad. Colomb. Cien. Exact. 8: 61. 1950. TYPE: Colombia. Valle del Cauca: Río Cajambre, Barco, 5--80 m, Apr. 1944 (stam. fl), Cuatrecasas 17188 (holotype, F).
Bud scales up to 7 mm above the node. Petioles indistinct, ca. 1--2 mm; leaf blades ovate, 13--15 x 7--9 cm, the base cordate and ± amplexicaul, the apex abruptly acujminate, drying slightly glaucous above, light tan on both surfaces; lateral veins (3--)5--10 mm apart, merging to a distinct submarginal vein ca. 2 mm from margin, the intersecondaries faint but occasionally reaching to the submarginal vein; translucid resin dashes faint. Inflorescence with ca. 1--3 flowers (in bud). Fruits unknown.
Distribution. Colombia, from near sea level.
This species is still known only from the type collection, material with immature staminate flowers only. The large, amplexicaul leaves with short petioles and distant primary lateral veins distinguish it from all other species. In leaf color and texture C. amplexicaulis is similar to C. elegans.
Clusiella axillaris (Engler) Cuatrecasas, Rev. Acad. Colomb. Cien. Exact. 8: 61. 1950. Clusia axillaris Engler in Mart., Fl. Brasil. 12: 413. 1888. Astrotheca cuspidata Vesque, Epharm. Genitalia Foliaque Clusiearum et Moronobearum. 3: 16, Tab. 80 & 81. 1892. LECTOTYPE here designated: Brazil. Amazonas: São Gabriel de Cachoeira: Rio Negro, Jan.--Aug. 1852 (stam. fl), Spruce 2159 (syntypes, GH, GOET; photo A, MO).
Bud scales sometimes persistent, 5--20 mm long, the scars up to 20 mm above the node. Petioles 49 mm, deeply channeled; leaf blades elliptic, 10--13(--14) x 4--6(--7) cm, the base acute to rounded, the apex acute to acuminate to a 5--20 mm acumen, drying somewhat shiny, dark olive gray above, tan below, lateral veins 2.5--5 mm apart, merging to an indistinct submarginal vein 1 mm or less from the margin, intersecondaries ca. 1--2 mm apart; translucid resin dots visible only with intense light or on thin, new leaves. Inflorescence usually 1-flowered; flower buds to 1.5 cm diam.; petals to 2 x 1.3 cm. Staminate flowers with the staminal column, including capitulum, ca. 5--8 mm tall. Fruit to ca. 4 cm diam., the stigmatic disk with ca. 20 closely packed rectangular stigmas, fruit wall rather thick, ca. 0.5 mm.
Distribution. Amazonian Brazil, Colombia and Venezuela, from ca. 100 to 750 m.
This species is easily recognized by virtue of its black twigs, dark drying leaves, large flowers and large fruits. It is distinctive also for its Amazonian distribution. It is probably related to C. impressinervis and C. pendula.
Additional specimens examined. BRAZIL. Acre: E slope of Serra da Moa, Apr. 1971 (stam. fl), Prance et al. 12584 (MO); Rio Negro, Rio Cauaburí, 750 m, Nov. 1965 (stam. fl), Maguire et al. 60422 (F, GH). Amazonas: Maraa, Rio Japurá, Dec. 1982 (pist. fl), Plowman et al. 12252 (F); São Paulo de Olivença, Apr. 1944 (fl, fr), Ducke 1626 (A, F, MO); basin of Rio Solimões, Dec. 1936 (stam. fl), Krukoff 8626 (A, F, MO); basin of creek Belem, (stam. fl), Krukoff 8933 (A, F, MO), Rio Solimões, Igarapé Jandiaruba, Jan. 1969 (fl), Fróes 23906 (GH). Pará: Road BR 22, Camparema to Maranhão, km 66, vicinity of Piriroro, Nov. 1965 (pist. fl), Prance & Pennington 1971 (GH). COLOMBIA. Amazonas: Leticia, Tarapacá, Parque Nacional Natual Amacayacu, 200 m, Nov. 1991, Pipoly et al. 16055 (MO), 100 m, July 1992, Rudas et al. 5471 (MO). Guainía: San Felipe Neri, on Río Negro, across from San Carlos de Río Negro, Venezuela, 120 m, May 1979 (stam. fl), Clark & Clark 7161 (MO). Vaupés: Río Apaporis, Sorajama (above mouth of Río Kananarí) and vicinity, 275 m, (pist. fl), Schultes & Cabrera 16089 (GH); Mitrú & vicinity, along Río Vaupes about 6 km below Mirú at Tukanaré, Aug. 1976 (fr), Zarucchi et al. 1883 (GH). PERU. Loreto: Maynas, Mishana, Río Nanay between Iquitos and Santa María de Nanay, 150 m, Mar. 1979 (stam. fl), Gentry & Aronson 25305 (F, MO), 130 m, Nov. 1978 Diaz et al. 1984 (MO); Alipahuayo (Estación IIAP), Nov. 1984 (pist. fl), Vásquez et al. 5887 (F, MO); Requena, Jenaro Herrera, Río Ucayali, May 1982 (fr), Encarnación 26132 (MO); Sapuena, Jenaro Herrera, 170 m, Nov. 1987 (stam. fl), Vásquez et al. 10051 (F, MO). VENEZUELA. Territorio Federal Amazonas: Dept. Atures, Río Autana, 90--110 m, Nov. 1984 (fr), Guanchez 3249 (MO); Dept. Río Negro, middle part of Río Baría, 80 m, July 1984 (fr), Davidse 27549 (MO); 3 km NE of San Carlos de Río Negro, 120 m, Apr. 1979 (stam. fl), Liesner 6777 (MO); Ríos Pacimoni-Yasya, Casiquiare, 100--140 m, Jan. 1954 (stam. fl), Maguire, Wurdack & Bunting 37439 (GH); Río Negro, Picora de Cocuí, Dec. 1947 (stam.? fl), Schultes & López 9436 (GH).
Clusiella elegans Planchon & Triana, Ann. Sci. Nat. Bot. 13 (ser 4): 254. 1860. TYPE: Colombia. Chocó: 2000 m, 1866 (fr), Triana s.n. (holotype, P; isotypes, F-fragment, G [photo F, MO], W [photo MO]).
Bud scales inconspicuous, their scars clustered at or just above the nodes. Petioles 1--2(--3) mm; leaf blades elliptic, 4--4.5(--10) x 1--1.5(--4.5) cm, the base acute (rarely rounded), the apex acuminate to a 1--1.5(--2) cm acumen, drying glaucous and olive green above, shiny and tan below, lateral veins 2.5--5 mm apart, merging to an indistinct submarginal vein ca. 1 mm from the margin, intersecondaries faint, ca. 1--2 mm apart; resin dots and dashes translucid or sometimes dark. Inflorescences unbranched or often with up to 6 branches; flower buds 2--4(--6) mm diam.; petals ca. 4 x 3 mm. Staminate flowers poorly known, the staminal column, including capitulum, ca. 2(6?) mm tall. Fruits ovoid, 4--8(--12) x 3--5(--10) mm, somewhat apiculate; stigmas 5--7, ± distinct, circular.
Distribution. Panama and Colombia from near sea level to 2000 m.
Originally described from pistillate material, this species is still known from very few staminate collections. Only three such collections are known: one from the disjunct population in Panama (Hammel & Kress 11287) and two from Colombia (Garcia-Barriga 11130 and Luteyn et al. 10682). In general, the plants are much more delicate than the other species, but C. elegans is particularly distinguished by its small, ovoid fruits with 5--7, more or less distinct stigmas. As discussed below, the large-leaved forms of C. elegans look much like C. isthmensis and also like the small-leaved forms of C. macropetala. Indeed, the glaucous upper surface of the leaves, as well as their color and venation, suggest that the three species are closely related. Betancur 802 (Cited below) has leaves that are large for the species and somewhat glaucous below as in C. macropetala. The fruits of that collection are also large for the species, but in concordance with placement in C. elegans they have a thin and leathery, rather than thick and bony fruit wall, and the stigmas are few and ± distinct.
Additional specimens examined. PANAMA. Coclé: between Llano Grande and Coclecito, 600 m, July 1979 (fr), Antonio 1396 (MO), Dec. 1979 (fr), Antonio 3050 (MO); Mar. 1982 (stam. fl), Hammel & Kress 11287 (DUKE). COLOMBIA. Antioquia: Amalfi a Rumazón, 1550 m, Sep. 1988 (fr), Betancur et al. 802 (MO). Chocó: between La Oveja and Quibdó, Apr. 1931 (fr), Archer 1753 (US); 15 km E of Quibdó, 75 m, Apr. 1931 (fr), Archer 2214 (US); Quibdó, Carretera Yute--Lloró, 70 m, Sep. 1976 (fr), Forero & Jaramillo 2721 (MO), 80 m, June 1983 (fr), Forero et al. 9700 (MO), 100 m, Aug. 1976 (fr), Gentry & Fallen 17817 (pro parte, mixed collection with C. macropetala, MO), Nov. 1988 (fr), Ramirez 1 (MO); highway Bolivar--Quibdó, km 52--70, 500--600 m, July 1944 Garcia-Barriga 11130 (US); km 175--176, 456 m, Mar. 1984 (fr), Juncosa 2578 (MO); 37--40 km W of El Carmen, 671--1360 m, May 1984 (stam. fl), Luteyn et al. 10682 (MO); road to Cabí SW of Tutuendo, 100 m, 19 Jan. 1979 (fr), Gentry & Renteria 24450 (MO); Río Serrano, 4--6 km arriba de Guayabal, 50 m, Apr. 1975 (fr), Forero et al. 1362 (MO). Valle del Cauca: Buenaventura, Bajo Calima, 50 m, July 1988 (fr), Croat 69479, 69515 (MO), Dec. 1981 (fr), Gentry 35589 (MO), ca. 100 m, July 1984 (fr), Gentry & Monsalve 48413 (MO), Feb. 1989 (fl), Gentry et al. 65553 (MO), June 1987 (fr), Monsalve 1509 (MO), Oct. 1987 (fr), Monsalve 1982 (MO), 50--100 m, Feb 1984 (fr), Juncosa 2125 (MO), Buenaventura to Cali, 100 m, June 1944 (fr), Killip & Cuatrecasas 38926 (F, GH, US); Punta Arenas, Buenaventura Bay, near sea level, June 1944 (fr), Killip & Cuatrecasas 38647 (US); Quebrada de Aguadulce, 0--10 m, Feb 1946 (pist.? fl), Cuatrecasas 19996 (F, US).
Clusiella impressinervis Hammel, sp. nov. TYPE: Brazil. Amazonas: Tonantins, Mar. 1944 (stam. fl), Ducke 1625 (holotype, US; isotypes, A, F). Figure 7.

Figure 7. Clusiella impressinervis Hammel; Ducke 1625. ---A. Habit. ---B. Bud scales. ---C. Base of leaf, abaxial view. ---D. Base of leaf, adaxial view.
- A speciebus aliis Clusiellae in costa supra impressa et punctis resiniferis utrinque foliorum manifestis, differt. Clusiella axillaris fortasse affinis, sed foliis et floribus minoribus.
Bud scales often persistent, several sets borne just above the node and one pair borne further above. Petioles 2--3 mm, deeply channeled; leaf blades elliptic to obovate, widest at or above the middle, (7--)8--9 x (2.5--)3--3.5 cm, the base acute, the apex abruptly acuminate to a 1.5-cm acumen, drying shiny, light green both surfaces; lateral veins mostly 1.5(--3) mm apart, merging to a very indistinct submarginal vein mostly hidden along the recurved margin; resin dots very conspicuous, both surfaces. Inflorescences apparently 1-flowered; flower buds 7--8 mm diam.; petals ca. 10 mm long x 7 mm wide, white. Staminate flowers with the staminal column, including capitulum, ca. 4 mm tall, filaments ± free apically but congested. Pistillate flowers and fruits unknown.
Distribution. Brazil and Venezuela, from ca. 50 to 1220 m
Since Clusiella impressinervis is so poorly known it is difficult to relate to the other species. However, by virtue of its non-glaucous leaves, its deeply channeled petioles, its persistent bud scales, and its Amazonian distribution, the species may be related to C. axillaris. Leaves of that species also occasionally have resin dots visible on the upper surface. The new species differs from C. axillaris by its smaller leaves and flowers and from all species not only by its impressed midrib but also because its leaves dry light green on both surfaces. More material, especially fruiting, is needed to clarify its relationships.
Additional specimens examined. VENEZUELA. Bolívar: Cerro Venamo, 1220 m, Jan. 1964 (fl. bud), Steyermark et al. 92790 (F, US).
Steyermark et al. 92790 will key to C. impressinervis by virtue of its impressed midrib and resin dots on both leaf surfaces. It also has deeply grooved petioles and somewhat persistent bud scales. However, it differs from the type by its ovate leaves, ca. 6--7 x 3 cm, which are widest below the middle, have a rounded base, and dry dull on both surfaces, the upper olive gray and the lower reddish tan, and by its shorter petioles only ca. 1--2 mm long. It is also found at a much higher elevation and is the easternmost collection known of the genus. By virtue of sharing unusual vegetative characters with C. impressinervis this specimen is tentatively included here. The description of the species is based on the type collection alone.
Clusiella isthmensis Hammel, sp. nov. TYPE: Costa Rica. Limón: Cerro Coronel, 5 m, Sep. 1986 (fl, fr), Davidse & Herrera 31421 (holotype, CR; isotypes, F, MO) Figure 8.

Figure 8. Clusiella isthmensis Hammel; Davidse & Herrera 31421. ---A. Habit. ---B. Pist. flower. ---C. Base of leaf, abaxial view. ---D. Base of leaf, adaxial view.
Clusiella macropetala Cuatrecasas affinis sed in exocarpio coriaceo (non osseo) folia membranaciore, et petiolio breviore differt.
Bud scales mostly deciduous, their scars congested at or near the nodes, rarely persistent (Kennedy et al. 2394) with a pair up to 3.5 cm above the node. Petiole 2--3 mm, somewhat channeled; leaf blades elliptic-lanceolate, widest below the middle (6--)8--14 x (2--)3--4.5 cm, the base acute to rounded, the apex gradually acuminate to a 1--2 cm-acumen, in dried material brown to glaucous and gray above, tan and sometimes glaucous below, lateral veins 2--3(--5) mm apart, merging to an indistinct submarginal vein mostly less than 1 mm from and hidden along the recurved margin; translucid resin dots and dashes sometimes also surficially visible on lower surface. Inflorescences mostly 1(--3)-flowered; petals ca. 10 x 6 mm, white. Staminate flowers with the staminal column, including capitulum, ca. 6 mm tall, the filaments free for a short distance apically. Pistillate flowers with apparently 10 stigmas but these forming a disk and difficult to distinguish; ovary 10-celled. Fruits ca. 1 cm diam. at maturity, translucent white, soft.
Distribution. Costa Rica and Panama, from near sea level to 1500 m.
Although this species was previously identified as Clusiella elegans (D'Arcy, 1980; Hammel, 1986) it is more closely related to C. macropetala. As here understood, C. elegans is generally a much more delicate plant with smaller leaves, flowers, and fruits than both C. isthmensis and C. macropetala. Most importantly, the ca. 10 stigmas of the latter two species are ± fused into a stigmatic cap, whereas those of C. elegans are fewer (ca. 5--7) and distinct. In addition to the characters mentioned in the diagnosis, C. isthmensis can be distinguished from C. macropetala by its generally smaller, more gradually acuminate leaf blades and by its shorter, thinner petioles. Nevertheless, in leaf size, shape, and color, both C. macropetala and C. elegans appear to overlap somewhat with C. isthmensis. The only other species that might be confused with C. isthmensis is C. pendula Cuatrecasas However, the leaves of this species dry darker with more prominent lateral veins, are shiny rather than glaucous, and the flowers are red or maroon rather than white. One collection, Foster et al. 14661, from lowland Bocas del Toro province, Panama, is problematic. The leaf size, shape, and coloration, as well as flower color and fruit size, all correspond with C. isthmensis, but the fruits have fewer and nearly separate stigmas, more like those of C. elegans.
Original field notes for the type of this species indicate 11 duplicates. These were distributed as C. elegans, and their whereabouts, except as indicated above, are uncertain.
Additional specimens examined. COSTA RICA. Heredia: La Selva Biological Station just E of the juncture of the Sarapiquí and Puerto Viejo Rivers, ca. 100 m, (all collections from La Selva are pistillate), Apr. 1981, Folsom 9673 (DUKE), Apr. 1981, Folsom 9893 (DUKE, F, MO), Mar. 1980, Hammel 8252 (DUKE, F, MO), May 1980, Hammel 8625 (DUKE), June 1980, Hammel 8934 (DUKE), July 1980, Hammel 9286 (DUKE), June 1982, Hammel & Trainer 12721 (DUKE), Aug. 1981 Damon Smith 109 (DUKE, MO), Oct. 1981, Damon Smith 457 (DUKE, F, MO); Parque Nac. Braulio Carrillo, between Río Peje and Río Sardinal, Atlantic slope of Volcán Barva, 1200--1400 m, Nov. 1985 (fr), Grayum & Herrera 7867 (CR, MO); Estación el Ceibo, 520 m, Dec. 1992 (fr), Boyle 1448 (CR); along W fork of Río Sardinal, Atlantic slope of Volcán Barva, 670 m, Dec. 1987 (fr), Grayum 8501 (F, INB, MO). Limón: ca. 2 km S of Río Colorado along new road and ca. 1 km E, 5 m, Mar. 1987 (fl, fr), Stevens et al. 25074 (MO); Cantón de Talamanca, entre Sukút y Amubri, 700--900 m, July 1989 (stam. fl), Hammel et al. 17625 (CR); Alto Urén, 1190 m, July 1989 (stam. fl), Herrera 3368 (INB, MO); Alto Lari, 1300 m, Mar. 1992 (stam. fl), Herrera 5190 (INB, MO); 1500 m, Mar. 1992 (stam. fl) Herrera 5422 (INB, MO). PANAMA. Coclé: Continental divide above El Copé, 750 m, Feb 1982 (stam. fl), Knapp & Dressler 3410 (MO). Darién: Río Tuquesa headwaters, ca. 2 km from continental divide, ca 400 m, Aug. 1974 (fl, fr), Croat 27213 (MO). Panamá: El Llano--Cartí road, ca. 350 m, Mar. 1973 (stam. fl), Croat 22894 (MO); Feb 1973, Kennedy et al. 2394 (MO); Feb 1973, Kennedy et al. 2397a (MO); Mar. 1973 (fl, fr), Liesner 658 (MO); Mar. 1973 (stam. fl), Liesner 1208 (MO); Nov. 1985 (fl), McPherson 7585 (MO); Feb 1975 Mori et al. 4701 (MO); Feb 1975 (fl, fr), Mori et al. 4705 (MO). San Blas Comarca: El Llano--Cartí road, 13.8--15.8 km N of Interamerican Highway, 325 m, Aug. 1984 (stam. fl), de Nevers et al. 3747 (MO); 19.1 km N of Interamerican Highway, 350 m, Nov. 1984 (fr), de Nevers 4300 (MO). Veraguas: Atlantic slope NW of Santa Fe, 450550 m, Dec. 1974 (stam. fl), Mori et al. 3875 (MO).
Clusiella macropetala Cuatrecasas, Ann. Inst. Biol. Mex. 20: 110. 1949. TYPE: Colombia. Valle del Cauca: Río Cajambre, La Trojita, 5--50 m, 29 Feb 1944 (stam. fl), Cuatrecasas 16578 (holotype, F; isotype, US).
Bud scales deciduous, the scars clustered at or just above the nodes. Petioles 3--10 mm, shallowly channeled; leaf blades elliptic, (10--)13--19 x (4--)6--8 cm, the base acute to rounded, the apex acute to acuminate to a 5--15 mm-acumen, in dried material glaucous on both surfaces, pale gray-green above, tan below, distinctly coriaceous; lateral veins 3--5(--7) mm apart, merging to a submarginal vein ca. 1 mm from margin, the intersecondaries often ± prominent; translucid resin dots and dashes visible only by intense light. Inflorescences unbranched or branched several times, with ca. 1--6 flowers; flower buds ca. 7--10 mm diam.; petals 10--17 x 6--10 mm. Staminate flowers with a staminal column, including capitulum, 5--10 mm tall. Fruits 8--12 mm diam., the stigmatic crest with ca. 10 closely packed stigmas, fruit wall rather thick, ca. 0.7 mm.
Distribution. Colombia, from near sea level to 700 m.
As noted above, the leaves of C. macropetala are glaucous on both surfaces and the fruits have a thick, several-layered fruit wall that is bony when dry. Smaller-leaved and smaller-flowered forms may otherwise be confused with C. elegans and C. isthmensis (q.v.), but C. macropetala is probably more closely related to the latter.
Additional specimens examined. COLOMBIA. Antioquia: Anorí, Valle de Anorí, entre Dos Bocas y Norí, 700 m, June 1971 (stam. fl), Soejarto 2869 (F, MO, GH). Chocó: Quibdó--Istmina road to Lloró, ca. 100 m, June 1983 (fl, fr), Forero et al. 9562 (MO), June 1983 (stam. fl), Forero et al. 9591 (MO), June 1983 (fl, fr), Forero et al. 9596 (MO), June 1982 (stam. fl), Gentry & Brand 36855 (MO), June 1982 (fl, fr), Gentry & Brand 36958 (MO), June 1982 (fl, fr), Gentry & Brand 36963 (MO), Aug. 1976 (stam. fl), Gentry & Fallen 17817 (pro parte, mixed collection with C. elegans, MO); San José del Palmar, hoya del Río San Juan, alrededores de Docordó, 0 m, Mar. 1979 (stam.? fl), Forero et al. 4328a (MO); Tutuendo--Quibdó road to Tubadó, 90 m, Jan. 1979 (fr), Gentry & Renteria 24507 (MO). Valle del Cauca: Bajo Calima, Buenaventura, 100 m, Jan. 1988 (stam. fl), Monsalve 2074 (MO); Río Cajambre, 580 m, May 1944 (stam. fl), Cuatrecasas 17497 (F, US).
Clusiella pendula Cuatrecasas, Ann. Inst. Biol. Mex. 20: 109. 1949. TYPE: Colombia. Valle del Cauca: Bajo Calima, Buenaventura, Bahia de Buenaventura, 0--10 m, 20 Feb 1946 (stam. fl), Cuatrecasas 19877 (holotype, F; isotype, US)
Clusiella cordifolia Cuatrecasas, Rev. Acad. Colombiana Cien. 8: 61. 1950. Syn. nov. TYPE: Colombia. Valle del Cauca: Bajo Calima, Buenaventura, coastal thickets, Killip 11731 (holotype US; isotype GH).
Bud scales deciduous, the scars clustered at or just above the nodes. Petioles ca. 3--5 mm, shallowly channeled; leaf blades elliptic to cordate, (9.5--)11.5--16(--19) x (3.5--)4--6.5(--8) cm, the base acute or rounded to sometimes cordate, the tip acuminate to a ca. 2 cm acumen, in dried material often shiny, (not glaucous) on both surfaces and dark olive above, dark brown below; lateral veins 2.5--4 mm apart, merging to a submarginal vein ca. 1 mm from margin, intersecondaries often also quite prominent; resin dots and dashes translucid. Inflorescence usually much-branched and with several flowers; flower buds ca. 5 mm diam.; petals ca. 10 x 5 mm, purplish-brown (maroon) to red. Staminate flowers with a staminal column, including capitulum ca. 3 mm tall. Fruits globose or slightly ovoid, ca. 15--20 x 10--15 mm, green(?) at maturity; stigmas ca. 8 or 9, closely packed and difficult to distinguish on fruits.
Distribution. Coastal Colombia and Ecuador from near sea level to ca. 800 m.
This species is easy to distinguish by virtue of its dark red instead of white or pinkish flowers and especially by its dark-drying, shiny leaves with prominent lateral veins. For the shape and color of its leaves it is most similar to the amazonian C. axillaris.
The many new collections from Colombia, Valle Dept., Buenaventura in Bajo Calima, near the type locality of both C. pendula and C. cordifolia, allow the conclusion that the latter should be considered a synonym of the former. The leaf color, texture and venation of the type of C. cordifolia coincide with those of C. pendula. Although the type specimens differ in shape of the leaf base, it is now obvious that this can vary from acute through rounded to slightly cordate even on a single specimen.
A label or numbering mix-up may have occurred with the type specimen of C. cordifolia. As noted by Cuatrecasas in the original description of the species, the label reads "Tree, fruit blue." However, not only are trees and blue fruits otherwise unknown in the genus, the specimen is of a flowering staminate individual!
Additional specimens examined. Colombia. Valle del Cauca: Bajo Calima, Buenaventura, Bahia de Buenaventura, Apr. 1939 (stam. fl), Killip 34966 (F, US); coastal thickets, Oct. 1922 (stam. fl), Killip 11731 (GH, US); el Forge, sea level, June 1944 (fl, fr), Killip & Cuatrecasas 38962 (F, US); Carretera Hans, km 22, 50 m, Dec. 1987 (fl), Gentry et al. 59554 (MO); Concesión Cartón de Colombia, 50--100 m, July 1987 (stam.? fl), Faber-Langendoen & Renteria 1209 (MO), Dec. 1981 (sterile), Gentry 35301 (MO), Feb 1983 (stam. fl), Gentry et al. 40394 (MO), Mar. 1986 (stam. fl), Gentry et al. 53618 (MO), Apr. 1987 (stam. fl), Gentry et al. 56701 (A, MO), Nov. 1986 (stam. fl), Monsalve 1206 (MO), Dec. 1986 (stam. fl), Monsalve 1405 (MO), Apr. 1987 (stam. fl), Monsalve 1457 (MO), Sep. 1987 (fr), Monsalve 1823 (MO), Oct. 1987 (stam. fl), Monsalve 1892 (MO), Oct. 1987 (fr), Monsalve 1933 (MO), Oct. 1987 (stam. fl), Monsalve 1980 (MO), Jan. 1988 (stam. fl), Monsalve 2083 (MO), June 1982 (stam. fl), Murphy 558 (MO); Concesión Pulpapel, 100 m, Aug. 1984 (fr), Monsalve 324 (GH); Quebrada Algeria, Bahia Malaga, 50 m, Dec. 1985 (pist. fl), Gentry et al. 53334 (MO); Río Calima, La Esperanza, 5--10 m, Mar. 1944 (stam. fl), Cuatrecasas 16761 (F, US); road to Juanchaco Palmeras, 100 m, July 1984 (pist. fl), Gentry et al. 47819 (MO), July 1984 (fr), Gentry et al. 48291 (MO). ECUADOR. Esmeraldas: San Lorenzo, 22 km from Lita on road to San Lorenzo, 800 m, May 1990 (fl), Gentry et al. 69990 (MO); Alto Tambo, 15 km oeste de Lita, 400 m, Sep. 1990 (fr), Rubio 618 (MO); Carretera Lita--Alto Tambo--La Punta, 400 m, Feb. 1991, Gudiño & Moran 1263 (MO).
Discussing the placement of Clusiella within the family, Planchon and Triana (1860) pointed out that the contorted aestivaton of the petals was characteristic of the tribe Moronobeae (subfamily Moronoboideae of Engler, 1925). However, because of the sessile, largely cupuliform stigmas and nonascendent ovules of Clusiella, they preferred to place it in their tribe Clusieae, although they were not certain of their disposition because of the lack of fruits and staminate flowers. Engler (1925) also felt that these "incompletely known" plants "probably belong" next to Clusia in subfamily Clusioideae. Subsequent published accounts have made no explicit statements about possible relationships of Clusiella, although most imply a close relationship with Clusia (but see Hammel, 1984, unpublished thesis).
Numerous additional characters, including interpetiolar stipliform structures, bud scales, resin gland-dots in the leaves, psilate pollen exine, baccate fruits, small fovelate seeds that lack an aril, and an embryo with unusually large cotyledons, are now known for Clusiella and make this genus very unusual if not misplaced among the clusioid genera. Stipuliform structures are uncommon in Clusiaceae, but ones similar to those found in Clusiella also occur in the Moronoboideae (e.g., Moronobea, Symphonia) and in Garcinia s.l. Bud scales also occur in Moronobea and Symphonia (and in some related genera), while some species of Garcinia have one or two pairs of bud scales, and bud scales occur in some of the Calophylloideae. Contorted petal aestivation does not otherwise occur within the whole subfamily Clusioideae (Engler, 1925), but is most likely the primitive state for the family (P. Stevens, pers. comm., 1987). The psilate pollen exine of Clusiella also appears to be otherwise unknown within the family except for Symphonia and Moronbea (Cf. Seetharam, 1983: It should be noted that Seetharam's study and light micrographs of "Clusiella elegans Klug 1950," describing it as having "perreticulate" exine, pertain to a species of Quapoya (i.e., Clusia), not Clusiella). The small, foveolate seeds and embryo with well-developed cotyledons of Clusiella are most like those in the Hypericoideae and basal Calophylloideae, e.g. Marila. Were it not for its epiphytic habit, dioecy, and resiniferous androecium (all reminiscent of Clusia), but particularly because of its small foveolate seeds and embryo structure, Clusiella would seem to be closely related to Symphonia. More detailed, comparative studies of pollen and the foliar resin system in particular are needed.
Characters such as those noted above in a sense pull Clusiella strongly outward and downward from Clusia. Work in progress towards an understanding of relationships among genera in the whole family, in fact suggests that Clusiella may be sister to a large clade including both the Clusioideae and Moronoboideae (P. Stevens, pers. comm.). The diminutive Clusiella, distinctively derived in many ways, and long masquerading as Clusia's little cousin, seems to be an epiphytic refugium for a number of relatively primitive characters in the Clusioideae-Moronobiodeae, and could be called, more accurately, Clusia's great aunt.
Acknowledgments. This work was supported, in part, by funds from the National Science Foundation through grants (eg., DEB-9300814) to the author and co-PI M. H. Grayum for the Manual to the Plants of Costa Rica. An early draft of the manuscript benefitted greatly through correspondence with Peter Stevens. I thank John Meyers for the line drawings of the two new species.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}