EMBRYOPSIDA Pirani & Prado
Gametophyte dominant, independent, multicellular, initially ±globular, not motile, branched; showing gravitropism; glycolate oxidase +, glycolate metabolism in leaf peroxisomes [glyoxysomes], acquisition of phenylalanine lysase* [PAL], flavonoid synthesis*, microbial terpene synthase-like genes +, triterpenoids produced by CYP716 enzymes, CYP73 and phenylpropanoid metabolism [development of phenolic network], xyloglucans in primary cell wall, side chains charged; plant poikilohydrous [protoplasm dessication tolerant], ectohydrous [free water outside plant physiologically important]; thalloid, leafy, with single-celled apical meristem, tissues little differentiated, rhizoids +, unicellular; chloroplasts several per cell, pyrenoids 0; centrioles/centrosomes in vegetative cells 0, microtubules with γ-tubulin along their lengths [?here], interphase microtubules form hoop-like system; metaphase spindle anastral, predictive preprophase band + [with microtubules and F-actin; where new cell wall will form], phragmoplast + [cell wall deposition centrifugal, from around the anaphase spindle], plasmodesmata +; antheridia and archegonia +, jacketed*, surficial; blepharoplast +, centrioles develop de novo, bicentriole pair coaxial, separate at midpoint, centrioles rotate, associated with basal bodies of cilia, multilayered structure + [4 layers: L1, L4, tubules; L2, L3, short vertical lamellae] (0), spline + [tubules from L1 encircling spermatid], basal body 200-250 nm long, associated with amorphous electron-dense material, microtubules in basal end lacking symmetry, stellate array of filaments in transition zone extended, axonemal cap 0 [microtubules disorganized at apex of cilium]; male gametes [spermatozoids] with a left-handed coil, cilia 2, lateral, asymmetrical; oogamy; sporophyte +*, multicellular, growth 3-dimensional*, cuticle +*, plane of first cell division transverse [with respect to long axis of archegonium/embryo sac], sporangium and upper part of seta developing from epibasal cell [towards the archegonial neck, exoscopic], with at least transient apical cell [?level], initially surrounded by and dependent on gametophyte, placental transfer cells +, in both sporophyte and gametophyte, wall ingrowths develop early; suspensor/foot +, cells at foot tip somewhat haustorial; sporangium +, single, terminal, dehiscence longitudinal; meiosis sporic, monoplastidic, MTOC [= MicroTubule Organizing Centre] associated with plastid, sporocytes 4-lobed, cytokinesis simultaneous, preceding nuclear division, quadripolar microtubule system +; wall development both centripetal and centrifugal, 1000 spores/sporangium, sporopollenin in the spore wall* laid down in association with trilamellar layers [white-line centred lamellae; tripartite lamellae]; plastid transmission maternal; nuclear genome [1C] <1.4 pg, main telomere sequence motif TTTAGGG, KNOX1 and KNOX2 [duplication] and LEAFY genes present, ethylene involved in cell elongation; chloroplast genome with close association between trnLUAA and trnFGAA genes [precursors for starch synthesis], tufA, minD, minE genes moved to nucleus; mitochondrial trnS(gcu) and trnN(guu) genes +.
Many of the bolded characters in the characterization above are apomorphies of more or less inclusive clades of streptophytes along the lineage leading to the embryophytes, not apomorphies of crown-group embryophytes per se.
All groups below are crown groups, nearly all are extant. Characters mentioned are those of the immediate common ancestor of the group, [] contains explanatory material, () features common in clade, exact status unclear.
POLYSPORANGIOPHYTA†
Sporophyte well developed, branched, branching dichotomous, potentially indeterminate; hydroids +; stomata on stem; sporangia several, terminal; spore walls not multilamellate [?here].
II. TRACHEOPHYTA / VASCULAR PLANTS
Sporophyte long lived, cells polyplastidic, photosynthetic red light response, stomata open in response to blue light; plant homoiohydrous [water content of protoplasm relatively stable]; control of leaf hydration passive; plant endohydrous [physiologically important free water inside plant]; PIN[auxin efflux facilitators]-mediated polar auxin transport; (condensed or nonhydrolyzable tannins/proanthocyanidins +); borate cross-linked rhamnogalactan II, xyloglucans with side chains uncharged [?level], in secondary walls of vascular and mechanical tissue; lignins +; roots +, often ≤1 mm across, root hairs and root cap +; stem apex multicellular [several apical initials, no tunica], with cytohistochemical zonation, plasmodesmata formation based on cell lineage; vascular development acropetal, tracheids +, in both protoxylem and metaxylem, G- and S-types; sieve cells + [nucleus degenerating]; endodermis +; stomata numerous, involved in gas exchange; leaves +, vascularized, spirally arranged, blades with mean venation density ca 1.8 mm/mm2 [to 5 mm/mm2], all epidermal cells with chloroplasts; sporangia in strobili, sporangia adaxial, columella 0; tapetum glandular; sporophyte-gametophyte junction lacking dead gametophytic cells, mucilage, ?position of transfer cells; MTOCs not associated with plastids, basal body 350-550 nm long, stellate array in transition region initially joining microtubule triplets; archegonia embedded/sunken [only neck protruding]; embryo suspensor +, shoot apex developing away from micropyle/archegonial neck [from hypobasal cell, endoscopic], root lateral with respect to the longitudinal axis of the embryo [plant homorhizic].
[MONILOPHYTA + LIGNOPHYTA]Sporophyte growth ± monopodial, branching spiral; roots endomycorrhizal [with Glomeromycota], lateral roots +, endogenous; G-type tracheids +, with scalariform-bordered pits; leaves with apical/marginal growth, venation development basipetal, growth determinate; sporangium dehiscence by a single longitudinal slit; cells polyplastidic, MTOCs diffuse, perinuclear, migratory; blepharoplasts +, paired, with electron-dense material, centrioles on periphery, male gametes multiciliate; nuclear genome [1C] 7.6-10 pg [mode]; chloroplast long single copy ca 30kb inversion [from psbM to ycf2]; mitochondrion with loss of 4 genes, absence of numerous group II introns; LITTLE ZIPPER proteins.
LIGNOPHYTA†
Sporophyte woody; stem branching axillary, buds exogenous; lateral root origin from the pericycle; cork cambium + [producing cork abaxially], vascular cambium bifacial [producing phloem abaxially and xylem adaxially].
SEED PLANTS† / SPERMATOPHYTA†
Growth of plant bipolar [plumule/stem and radicle/root independent, roots positively geotropic]; plants heterosporous; megasporangium surrounded by cupule [i.e. = unitegmic ovule, cupule = integument]; pollen lands on ovule; megaspore germination endosporic, female gametophyte initially retained on the plant, free-nuclear/syncytial to start with, walls then coming to surround the individual nuclei, process proceeding centripetally.
EXTANT SEED PLANTS
Plant evergreen; nicotinic acid metabolised to trigonelline, (cyanogenesis via tyrosine pathway); microbial terpene synthase-like genes 0; primary cell walls rich in xyloglucans and/or glucomannans, 25-30% pectin [Type I walls]; lignin chains started by monolignol dimerization [resinols common], particularly with guaiacyl and p-hydroxyphenyl [G + H] units [sinapyl units uncommon, no Maüle reaction]; roots often ≥1 mm across, stele diarch to pentarch, xylem and phloem originating on alternating radii, cork cambium deep seated, gravitropism response fast; stem apical meristem complex [with quiescent centre, etc.], plasmodesma density in SAM 1.6-6.2[mean]/μm2 [interface-specific plasmodesmatal network]; eustele +, protoxylem endarch, endodermis 0; wood homoxylous, tracheids and rays alone, tracheid/tracheid pits circular, bordered; mature sieve tube/cell lacking functioning nucleus, sieve tube plastids with starch grains; phloem fibres +; cork cambium superficial; leaf nodes 1:1, a single trace leaving the vascular sympodium; leaf vascular bundles amphicribral; guard cells the only epidermal cells with chloroplasts, stomatal pore with active opening in response to leaf hydration, control by abscisic acid, metabolic regulation of water use efficiency, etc.; branching by axillary buds, exogenous; prophylls two, lateral; leaves with petiole and lamina, development basipetal, lamina simple; sporangia borne on sporophylls; spores not dormant; microsporophylls aggregated in indeterminate cones/strobili; grains monosulcate, aperture in ana- position [distal], primexine + [involved in exine pattern formation with deposition of sporopollenin from tapetum there], exine and intine homogeneous, exine alveolar/honeycomb; ovules with parietal tissue [= crassinucellate], megaspore tetrad linear, functional megaspore single, chalazal, sporopollenin 0; gametophyte ± wholly dependent on sporophyte, development initially endosporic [apical cell 0, rhizoids 0, etc.]; male gametophyte with tube developing from distal end of grain, male gametes two, developing after pollination, with cell walls; embryo cellular ab initio, suspensor short-minute, embryonic axis straight [shoot and root at opposite ends], primary root/radicle produces taproot [= allorhizic], cotyledons 2; embryo ± dormant; chloroplast ycf2 gene in inverted repeat, trans splicing of five mitochondrial group II introns, rpl6 gene absent; ??whole nuclear genome duplication [ζ/zeta duplication event], 2C genome size (0.71-)1.99(-5.49) pg, two copies of LEAFY gene, PHY gene duplications [three - [BP [A/N + C/O]] - copies], 5.8S and 5S rDNA in separate clusters.
IID. ANGIOSPERMAE / MAGNOLIOPHYTA
Lignans, O-methyl flavonols, dihydroflavonols, triterpenoid oleanane, apigenin and/or luteolin scattered, [cyanogenesis in ANA grade?], lignin also with syringyl units common [G + S lignin, positive Maüle reaction - syringyl:guaiacyl ratio more than 2-2.5:1], hemicelluloses as xyloglucans; root cap meristem closed (open); pith relatively inconspicuous, lateral roots initiated immediately to the side of [when diarch] or opposite xylem poles; epidermis probably originating from inner layer of root cap, trichoblasts [differentiated root hair-forming cells] 0, hypodermis suberised and with Casparian strip [= exodermis]; shoot apex with tunica-corpus construction, tunica 2-layered; starch grains simple; primary cell wall mostly with pectic polysaccharides, poor in mannans; tracheid:tracheid [end wall] plates with scalariform pitting, multiseriate rays +, wood parenchyma +; sieve tubes enucleate, sieve plates with pores (0.1-)0.5-10< µm across, cytoplasm with P-proteins, not occluding pores of plate, companion cell and sieve tube from same mother cell; ?phloem loading/sugar transport; nodes 1:?; dark reversal Pfr → Pr; protoplasm dessication tolerant [plant poikilohydric]; stomata randomly oriented, brachyparacytic [ends of subsidiary cells ± level with ends of guard cells], outer stomatal ledges producing vestibule, reduction in stomatal conductance with increasing CO2 concentration; lamina formed from the primordial leaf apex, margins toothed, development of venation acropetal, overall growth ± diffuse, secondary veins pinnate, fine venation hierarchical-reticulate, (1.7-)4.1(-5.7) mm/mm2, vein endings free; flowers perfect, pedicellate, ± haplomorphic, protogynous; parts free, numbers variable, development centripetal; P = T, petal-like, each with a single trace, outer members not sharply differentiated from the others, not enclosing the floral bud; A many, filament not sharply distinguished from anther, stout, broad, with a single trace, anther introrse, tetrasporangiate, sporangia in two groups of two [dithecal], each theca dehiscing longitudinally by a common slit, ± embedded in the filament, walls with at least outer secondary parietal cells dividing, endothecium +, cells elongated at right angles to long axis of anther; tapetal cells binucleate; microspore mother cells in a block, microsporogenesis successive, walls developing by centripetal furrowing; pollen subspherical, tectum continuous or microperforate, ektexine columellate, endexine restricted to the apertural regions, thin, compact, intine in apertural areas thick, orbicules +, pollenkitt +; nectary 0; carpels present, superior, free, several, spiral, ascidiate [postgenital occlusion by secretion], stylulus at most short [shorter than ovary], hollow, cavity not lined by distinct epidermal layer, stigma ± decurrent, carinal, dry; suprastylar extragynoecial compitum +; ovules few [?1]/carpel, marginal, anatropous, bitegmic, micropyle endostomal, outer integument 2-3 cells across, often largely subdermal in origin, inner integument 2-3 cells across, often dermal in origin, parietal tissue 1-3 cells across, nucellar cap?; megasporocyte single, hypodermal, functional megaspore lacking cuticle; female gametophyte lacking chlorophyll, four-celled [one module, egg and polar nuclei sisters]; ovule not increasing in size between pollination and fertilization; pollen grains bicellular at dispersal, germinating in less than 3 hours, siphonogamy, pollen tube unbranched, growing towards the ovule, between cells, growth rate (ca 10-)80-20,000 µm h-1, tube apex of pectins, wall with callose, lumen with callose plugs, penetration of ovules via micropyle [porogamous], whole process takes ca 18 hours, distance to first ovule 1.1-2.1 mm; male gametophytes tricellular, gametes 2, lacking cell walls, ciliae 0, double fertilization +, ovules aborting unless fertilized; fruit indehiscent, P deciduous; mature seed much larger than fertilized ovule, small [<5 mm long], dry [no sarcotesta], exotestal; endosperm +, ?diploid [one polar nucleus + male gamete], cellular, development heteropolar [first division oblique, micropylar end initially with a single large cell, divisions uniseriate, chalazal cell smaller, divisions in several planes], copious, oily and/or proteinaceous, embryo short [<¼ length of seed]; plastid and mitochondrial transmission maternal; Arabidopsis-type telomeres [(TTTAGGG)n]; nuclear genome [2C] (0.57-)1.45(-3.71) [1 pg = 109 base pairs], ??whole nuclear genome duplication [ε/epsilon event]; ndhB gene 21 codons enlarged at the 5' end, single copy of LEAFY and RPB2 gene, knox genes extensively duplicated [A1-A4], AP1/FUL gene, palaeo AP3 and PI genes [paralogous B-class genes] +, with "DEAER" motif, SEP3/LOFSEP and three copies of the PHY gene, [PHYB [PHYA + PHYC]]; chloroplast IR expansions, chlB, -L, -N, trnP-GGG genes 0.
[NYMPHAEALES [AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]]: wood fibres +; axial parenchyma diffuse or diffuse-in-aggregates; pollen monosulcate [anasulcate], tectum reticulate-perforate [here?]; ?genome duplication; "DEAER" motif in AP3 and PI genes lost, gaps in these genes.
[AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]: phloem loading passive, via symplast, plasmodesmata numerous; vessel elements with scalariform perforation plates in primary xylem; essential oils in specialized cells [lamina and P ± pellucid-punctate]; tension wood + [reaction wood: with gelatinous fibres, G-fibres, on adaxial side of branch/stem junction]; anther wall with outer secondary parietal cell layer dividing; tectum reticulate; nucellar cap + [character lost where in eudicots?]; 12BP [4 amino acids] deletion in P1 gene.
[MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]] / MESANGIOSPERMAE: benzylisoquinoline alkaloids +; sesquiterpene synthase subfamily a [TPS-a] [?level], polyacetate derived anthraquinones + [?level]; outer epidermal walls of root elongation zone with cellulose fibrils oriented transverse to root axis; P more or less whorled, 3-merous [?here]; pollen tube growth intra-gynoecial; extragynoecial compitum 0; carpels plicate [?here]; embryo sac monosporic [spore chalazal], 8-celled, bipolar [Polygonum type], antipodal cells persisting; endosperm triploid.
MAGNOLIIDS + CHLORANTHALES + EUDICOTS: Benzylisoquinoline alkaloids +.
EUDICOTS: (Myricetin +), asarone 0 [unknown in some groups, + in some asterids]; root epidermis derived from root cap [?Buxaceae, etc.]; (vessel elements with simple perforation plates in primary xylem); nodes 3:3; stomata anomocytic; flowers (dimerous), cyclic; protandry common; K/outer P members with three traces, ("C" +, with a single trace); A ?, filaments fairly slender, anthers basifixed; microsporogenesis simultaneous, pollen tricolpate, apertures in pairs at six points of the young tetrad [Fischer's rule], cleavage centripetal, wall with endexine; G with complete postgenital fusion, stylulus/style solid [?here], short [<2 x length of ovary]; seed coat?; palaeotetraploidy event.
THIS LOT (but getting fewer and fewer) UNPLACED - Main Tree, Synapomorphies.
Note: In all node characterizations, boldface denotes a possible apomorphy, (....) denotes a feature the exact status of which in the clade is uncertain, [....] includes explanatory material; other text lists features found pretty much throughout the clade. However, the precise node to which many characters, particularly the more cryptic ones, should be assigned is unclear. This is partly because homoplasy is very common, in addition, basic information for all too many characters is very incomplete, frequently coming from taxa well embedded in the clade of interest and so making the position of any putative apomorphy uncertain. Then there are the not-so-trivial issues of how character states are delimited and ancestral states are reconstructed (see above).
Maesaceae [Theophrastaceae [Myrsinaceae + Primulaceae]]: (schizogenous secretory canals [material yellow, red, brown: tannins, etc.]); nodes ?3:3; (stomata anisocytic); small ± immersed often peltate glandular hairs +; inflorescence racemose; C and A from common primordia, C connate, stamens = and opposite C, antesepalous whorl represented by at least a vascular trace [?Maesa], nectary +; G [5], opposite C, placentation free-central, ovules at least partly immersed in swollen placenta, apotropous, bitegmic, micropyle bistomal, endothelium +, tanniniferous, style short, hollow, stigma ± capitate; seeds angled; endotesta crystalliferous; endosperm nuclear, copious, cell walls thick, with amyloid or hemicellulosic.
Evolution: Wikström et al. (2001) suggest a stem group age of 75-72Ma before present, with crown group divergence beginning 49-46Ma before present.
Members of this group are not often eaten by butterfly larvae, but Lycaenidae-Riodininae-Hamearini and a few Riodinini (see also Abisara) are found on them, especially on Maesaceae but not so far on Samolus and Theophrastaceae (Ehrlich & Raven 1964).
Chemistry, Morphology, etc.. Leaves of Theophrastaceae and Myrsinaceae are often described as being involute (?supervolute, c.f. Cullen 1978) or conduplicate. There are common stamen/corolla primordia born on a ring primordium in this clade. but there is variation in the position/relative development of these primordia. In some cases such as Cyclamen the stamens are initiated as adaxial outgrowths of a common primordium, i.e. the petal primordia are early larger than the stamen primordia, as also in Myrsine and Aegiceras (see especially Ma & Saunders 2003), whereas the stamen primordia may initially be larger, as in Samolus (e.g. Sattler 1962). However, this is a tricky character, since there are really two variables, the relative positions of these priordia and how fast they initially develop, and, as with evicted terminal inflorescences, initial topolgical relationships between parts can speedily become disrupted by post-initiation growth. The number of carpels can be difficult to ascertain, but five seems to be a common number, however, the orientation is unclear. The diagrams presented by Dickson (1936) mostly suggest that the carpels are opposite the petals, but in Primula, at least, they suggest that the carpels are opposite the sepals.
For the hollow style, see Guéguen (1901: is Maesaceae known?), for staminodes, see Saunders (1936) and Caris and Smets (2004: those of Samolus and Theoprastaceae are developmentally rather different), for nectar secretion, see Vogel (1986, 1997) and Caris and Smets (2004), for embryology especially of the herbaceous taxa, i.e. Primulaceae in the old sense, see Dahlgren (1916), for wood anatomy, see Lens et al. (2005a), and for floral morphology and ontogeny, Dickson (1936: esp. gynoecial arrangement), Sattler (1962), Sundberg (1982), Ronse Decraene (1992), Ronse Decraene et al. (1995) and especially Ma and Saunders (2003). For general morphology, see Anderberg et al. (2000) and especially Ståhl and Anderberg (2004).
Phylogeny. The monophyly of the group is not in doubt (see Anderberg & Ståhl 1994; Anderberg et al. 1998; and especially Källersjö et al. 2000: note that support values for Samolus as sister to Theophrastaceae s. str. are reduced when morphological data are added to molecular).
Phylogeny. This whole group was often recognised as Primulales in the past. Perhaps the only question, particularly in light of the break-up of Primulaceae, the removal of Maesa from Myrsinaceae, the placement/addition of Samolus as sister to the old Theophrastaceae, the many herbaceous ex-Primulaceae that are sister to the old-style, woody Myrsinaceae rather than being in a clade with other Primulaceae, and the numerous features shared by the group as a whole, is whether it is worth recognising families at all... Subfamilial names are already available.
Previous relationships. Plumbaginaceae (see Caryophyllales here) were often associated with Primulaceae and related families because of similar placentation and stamen arrangement (see Cronquist 19181 for discussion).
MAESACEAE Anderberg, B. Ståhl & Kallersjö - Back to Ericales
Evergreen lianes or trees; vessel element type?; petiole bundles all annular; secretory canals well developed; leaves spiral or two-ranked, induplicate, margin toothed to entire; inflorescence often branched; flowers small, C induplicate-valvate, stamen primordium smaller than petal primordium, A basally connate, attached at the middle of the C tube, nectary on G; G [3-4], half inferior, ovules apotropous, endothelium +, stigma truncate or capitate and lobed; fruit a many-seeded drupe, K persistent; testa 2-layered, inner layer with rhombic crystals; n = 10.
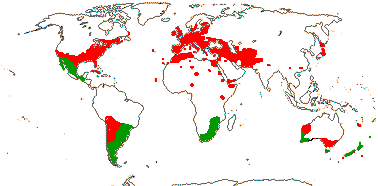
1/150. Old World tropics to Japan, the Pacific, and Australia (map: from Palgrave 2002).
Chemistry, Morphology, etc.. Vessels are in radial multiples (as quite commonly in woody Theophrastaceae and Myrsinaceae); there may be groups of druses in the abaxial epidermis; the fibres are septate; and the lateral bundles arise about half an internode below the leaf they supply. Information on floral development is taken from Caris et al. (2000); the ovules are separated by and partly sunken in placental tissue (see also Utteridge & Saunders 2001).
Theophrastaceae [Myrsinaceae + Primulaceae]: herbs[?]; rays ³5-seriate, uniseriate rays 0 [not herbaceous taxa]; bracteoles 0; C imbricate, arising abaxially on common primordium [i.e. stamen primordium > petal primordium], subrotate, tube rather short.
Phylogeny and Evolution: For the suggestion that rosette herbs may be the plesiomorphic condition for this part of the clade, see Anderberg et al. (2001); however, Lens et al. (2005a) find no evidence from wood anatomy that this is likely (apart from in a few Myrsinaceae). Note that in Myrsinaceae, herbaceous taxa such as Stimpsonia, Ardisiandra and Coris are basal to woody taxa, and variation in habit is very extensive in this clade as a whole. Smith and Donoghue (2008) found that the rate of mollecular evolution in the herbaceous taxa they examined was much greater than in the woody taxa.
THEOPHRASTACEAE Link, nom. cons. - Back to Ericales
Bracts displaced up the pedicels; staminodes +, petal-like, endothelium?
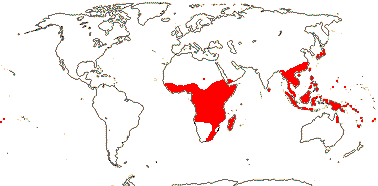
6-9[list]/105 - two groups below. Mostly New World and tropical, some also more temperate and Old World (map: from Hultén 1971).
1. Samolus
Nodes ?1:1; leaves entire; K connate, nectary on ovary; G [5], semi-inferior, style impressed; fruit a 5-valved capsule, seeds many; coat undistinguished, exotesta and endotegmen tanniniferous, the latter crystalliferous; endosperm cell walls thin; n = (12) 13.
1/15. America, the Antipodes, Europe, tropical to temperate (map: from Hultén 1971; Meusel et al. 1978; FloraBase 2005). [Photo - Flowers.]
Chemistry, Morphology, etc.. Ståhl (2004) suggests that a secretory system is present, if not always conspicuous. The stomata are anomocytic. There are several petiole bundles forming an arc, and these seem to diverge very soon after the leaf trace departs from the central stele. The ovules completely cover the placenta, but fingers of placental tissue may poke up between them (but not seen in the material examined by Caris & Smets 2004); Ma and Saunders (2003) suggest that in this whole clade (i.e. Theophrastaceae s.l.) the ovules are not embedded (which would then be a synapomorphy for it). The valves of the capsule are opposite the calyx (Caris & Smets 2004).
For general information, see Ståhl (2004: as Samolaceae).
Synonymy: Samolaceae Rafinesque
2. The Rest (Theophrastaceae s. str.)
Woody, tending to be pachycaul; rays broad; nodes also 1:1 [Jacquinia, dividing into three], 5:5 [Clavija]; secretory system?; petiole bundle deeply arcuate or annular, with small adaxial inverted bundles; scale leaves +; leaves conduplicate, margins spiny-toothed to entire, subepidermal fibres +; plant dioecious or flowers bisexual; anthers extrorse, with calcium oxalate, (nectariferous hairs +), style long, stigma dry or wet; fruit a (rather dry) berry, placentae ± pulpy, (drupe); seeds 1-few, rounded, exotestal cells flattened, thick-walled, hypodermal cells (with thickened anticlinal walls), often crystalliferous; endosperm cell walls pitted, cotyledons usu. foliaceous; n = 18, 20, 24.
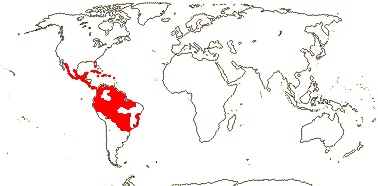
4/90: Clavija (50), Jacquinia (35 - perhaps to be divided). New World tropics (map: from Ståhl 1989, 1991, 1995). [Photos - Collection]
Chemistry, Morphology, etc.. The subepidermal fibres may lack lignification. For reports of glandular dots on calyx and corolla, see Mabberley (1997). Floral primordia may initially be quite strongly monosymmetric, as in Deherania (Sattler 1962), even if the flower at anthesis is polysymmetric.
For morphology, etc., see Ståhl (2004) and in particular Caris and Smets (2004).
Phylogeny. Phylogenetic relationships suggested by Källersjö and Ståhl (2003) imply that some generic realignments are needed.
Primulaceae + Myrsinaceae: two ndhF deletions.
PRIMULACEAE Borkhausen, nom. cons. - Back to Ericales
Cucurbitacins +; ?cork; glands 0, trichomes articulated; leaves involute or revolute, margins entire to dentate or serrate; inflorescence scapose; K often connate, C hypocrateriform; A attached at or above middle of C tube, pollen syn- or polycolpate, nectary on ovary, ovules not immersed in placenta (immersed - Dionysia), (inner integument ca 4 cells across), style usu. long, (heterostyly +); fruit a capsule; seeds many, angled, exotesta ± persistent, walls thickened or not, (endotesta with inner walls thickened [Primula]), endotegmen often crystalliferous; (endosperm cell walls thin); n = 8-12.
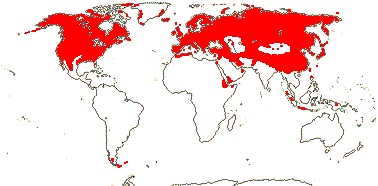
9[list]/900: Primula (490-600: inc. Cortusa, Dionysia [some chasmophytes, for which see Trift et al. 2004, relationships, biogeography; Lidén 2007, revision), Dodecatheon), Androsace (160: inc. Douglasia, Vitaliana, see Schneeweiss et al. 2004b). Northern hemisphere, scattered elsewhere (map: from Hultén 1971; Meusel et al. 1978). [Photo - Dodecatheon flower © R. Kowal] [Photo - Primula flower]
Evolution: Heterostyly is common, although it is unlikely to be an apomorphy for the family; it is sometimes lost, as in those Primula with buzz pollination, the erstwhile Dodecatheon (Mast et al. 2001, 2006).
Chemistry, Morphology, etc.. The involute leaves can be sharply bent rather than incurved (for vernation, see Conti et al. 2000; Mast et al. 2001). Solereder (1908) reports that secretory tissues occur in Androsace lactea. The corolla epidermal cells are isodiametric. Saunders (1936) suggested that some of the lobing of the corolla of Soldanella might be staminodial.
For pollen variation, see Mast et al. (2001), and for general information, see Anderberg (2004).
Phylogeny. For ITS-based relationships within the family, see Martins et al. (2003), and for relationships within Primula, see also Trift et al. (2002) and Mast et al. (2004, 2006), for relationships within Androsace, see Wang et al. (2004) and Schneeweiss et al. (2004b).
Classification. Richards (2003) provides a good general description of the species of Primula s. str.
MYRSINACEAE R. Brown, nom. cons. - Back to Ericales
Also trees to shrubs or lianes; benzoquinones +; (vessel elements with scalariform perforations); (nodes 3:3 - unnamed taxon from Atlantic Forest; Ardisia densiflora); glands/canals throughout the plant (0); leaves (opposite), also supervolute (curved), margins entire (crenate to serrate, teeth cartilaginous); (plant dioecious), inflorescence often fasciculate/corymbose; flowers (3-)4-5(-7)-merous, C often contorted, (nectariferous hairs +); A dorsifixed or basifixed, sagittate, (dehiscing by pores), (micropyle endostomal - Coris; endothelium 0; style 0; long), stigma (punctate), dry or wet; fruit a berry, drupe or capsule [latter in herbaceous taxa], placentae ± pulpy; seeds 1-few, rounded (ruminate; hilum depressed) [woody taxa] or many, small, angular, seed coat undistinguished, (endotesta crystalliferous - Cyclamen), tegmen thickened before becoming crushed, (endotegmen crystalliferous); endosperm walls pitted, (embryo slightly curved; medium); n = 10-13, 15, 17, 23.
41[list]/1435: Ardisia (450), Myrsine (155: inc. Rapanea, Suttonia, many species in the Pacific), Lysimachia (150: sometimes staminodes?, some woody; white flowers with nectariferous hairs, some yellow oil flowers [Macropis the pollinator], or selfers - see Vogel 1986), Discocalyx (115: inc. Tapeinosperma), Embelia (100), Parathesis (85), Stylogyne (60). Pantropical and N. Temperate (map: from Hultén 1958, 1971; FloraBase 2008: S. Hemisphere a bit notional). [Photos - collection woody members, Cyclamen flower © H. Schneider, fruit © H. Schneider], collection of ex Primulaceae.]
Evolution: Vogel (1986) discusses pollination, which in a group of yellow-flowered Lysimachia in particular is by oil-collecting Macropis (Mellitidae) bees (see also Simpson et al. 1983). The oil is secreted by trichomes. Anderberg et al. (2007) suggested that Lysimachia with buzz-pollinated flowers and those with nectar-producing hairs formed separate clades and were both derived from oil-producing ancestors.
Some species of Ardisia have pustules along the edge of the leaf blade; although inhabited by bacteria, it is unclear what role the bacteria might be playing (Miller 1990).
For the evolution of the mangrove habitat, to which Aegiceras is restricted, see Rhizophoraceae and Tomlinson (1986). Aegiceras has a number of anomalous anatomical and morphological features, the seed characters in particular are those that might be expected from a mangrove plant, since seeds lack endosperm and contain a large, viviparous embryo (c.f. Rhizophoraceae-Rhizophoreae, Acanthaceae-Acantheae-Acanthus ilicifolius, etc.).
Chemistry, Morphology, etc.. The presence of coloured glands may well not be a synapomorphy of the family (Hao et al. 2004). There are breakdown areas in the rays of woody members, and these may be filled with dark contents (Lens et al. 2005). Discocalyx has three traces in the petiole base, and some other taxa may be trilacunar; nodal anatomy needs study. The epidermal cells of the corolla are often elongated (as in Glaux - to be included in Lysimachia - lacks a corolla); this is a derived feature within the family. Trientalis has anisomerous flowers (Swenson et al. 2008c). Cyclamen has one cotyledon and one integument. See Oh et al. (2008) for the seed morphology of herbaceous taxa around Lysimachia.
Coris is a particularly distinctive genus morphologically. It is a small ericoid sub-shrub with monosymmetric flowers that have a spine-tipped epicalyx. There is nectary at the base of the ovary and there are only 5-6 ovules. Monosymmetry is expressed early in development by the calyx, but monosymmetry of the corolla becomes evident only later (Ronse Decraene et al. 1995: they suggest that the median sepal is abaxial, i.e. that the orientation of the flower is inverted or oblique). The rays of Aegiceras are relatively narrow, the ovules are unitegmic, etc. (Staåhl & Anderberg 2004; Lens et al. 2005).
Some information is taken from Otegui and Cocucci (1999) and from Ståhl and Anderberg (2004); Lens et al. (2005a) provide much information about wood anatomy.
Phylogeny. Myrsinaceae, previously circumscribed to include only woody taxa, now include Anagallis, Ardisiandra, Asterolinon (?= Lysimachia), Coris, Cyclamen, Glaux, Lysimachia, Pelletiera, Stimpsonia and Trientalis (Anderberg et al. 2000, 2001; the limits of Myrsinaceae are not so clear in Martins et al. 2003, but ITS data alone in the latter study. Anderberg et al. (2007) was particularly interested in the relationships of the herbaceous taxa; the family as a whole had moderate support as being monophyletic (72% jacknife), and Cyclamen, the herbaceous taxa, and the woody taxa then formed a trichotomy. Hao et al. 2004 also provide a phylogeny of the family, although focussing on Lysimachia.
Classification.Generic limits in the woody members in particular are unsatisfactory, but the limits of genera like Lysimachia are also unclear (Anderberg et al. 2007).
Synonymy: Aegicerataceae Blume, Anagallidaceae Adanson, Ardisiaceae Jussieu, Coridaceae J. Agardh, Embeliaceae J. Agardh, Lysimachiaceae Durande
Conniptions![Photo - Primula flower]](javascript:showImage('http://www.csdl.tamu.edu/FLORA/schoepke/pri_da_2.jpg',600,500)){kind=link}
){kind=link}
![fruit © H. Schneider]](javascript:showImage('http://131.152.161.2/bilder/43/y/4382.jpg',600,500)){kind=link}