EMBRYOPSIDA Pirani & Prado
Gametophyte dominant, independent, multicellular, initially ±globular, not motile, branched; showing gravitropism; glycolate oxidase +, glycolate metabolism in leaf peroxisomes [glyoxysomes], acquisition of phenylalanine lysase* [PAL], flavonoid synthesis*, microbial terpene synthase-like genes +, triterpenoids produced by CYP716 enzymes, CYP73 and phenylpropanoid metabolism [development of phenolic network], xyloglucans in primary cell wall, side chains charged; plant poikilohydrous [protoplasm dessication tolerant], ectohydrous [free water outside plant physiologically important]; thalloid, leafy, with single-celled apical meristem, tissues little differentiated, rhizoids +, unicellular; chloroplasts several per cell, pyrenoids 0; centrioles/centrosomes in vegetative cells 0, microtubules with γ-tubulin along their lengths [?here], interphase microtubules form hoop-like system; metaphase spindle anastral, predictive preprophase band + [with microtubules and F-actin; where new cell wall will form], phragmoplast + [cell wall deposition centrifugal, from around the anaphase spindle], plasmodesmata +; antheridia and archegonia +, jacketed*, surficial; blepharoplast +, centrioles develop de novo, bicentriole pair coaxial, separate at midpoint, centrioles rotate, associated with basal bodies of cilia, multilayered structure + [4 layers: L1, L4, tubules; L2, L3, short vertical lamellae] (0), spline + [tubules from L1 encircling spermatid], basal body 200-250 nm long, associated with amorphous electron-dense material, microtubules in basal end lacking symmetry, stellate array of filaments in transition zone extended, axonemal cap 0 [microtubules disorganized at apex of cilium]; male gametes [spermatozoids] with a left-handed coil, cilia 2, lateral, asymmetrical; oogamy; sporophyte +*, multicellular, growth 3-dimensional*, cuticle +*, plane of first cell division transverse [with respect to long axis of archegonium/embryo sac], sporangium and upper part of seta developing from epibasal cell [towards the archegonial neck, exoscopic], with at least transient apical cell [?level], initially surrounded by and dependent on gametophyte, placental transfer cells +, in both sporophyte and gametophyte, wall ingrowths develop early; suspensor/foot +, cells at foot tip somewhat haustorial; sporangium +, single, terminal, dehiscence longitudinal; meiosis sporic, monoplastidic, MTOC [= MicroTubule Organizing Centre] associated with plastid, sporocytes 4-lobed, cytokinesis simultaneous, preceding nuclear division, quadripolar microtubule system +; wall development both centripetal and centrifugal, 1000 spores/sporangium, sporopollenin in the spore wall* laid down in association with trilamellar layers [white-line centred lamellae; tripartite lamellae]; plastid transmission maternal; nuclear genome [1C] <1.4 pg, main telomere sequence motif TTTAGGG, KNOX1 and KNOX2 [duplication] and LEAFY genes present, ethylene involved in cell elongation; chloroplast genome with close association between trnLUAA and trnFGAA genes [precursors for starch synthesis], tufA, minD, minE genes moved to nucleus; mitochondrial trnS(gcu) and trnN(guu) genes +.
Many of the bolded characters in the characterization above are apomorphies of more or less inclusive clades of streptophytes along the lineage leading to the embryophytes, not apomorphies of crown-group embryophytes per se.
All groups below are crown groups, nearly all are extant. Characters mentioned are those of the immediate common ancestor of the group, [] contains explanatory material, () features common in clade, exact status unclear.
POLYSPORANGIOPHYTA†
Sporophyte well developed, branched, branching dichotomous, potentially indeterminate; hydroids +; stomata on stem; sporangia several, terminal; spore walls not multilamellate [?here].
II. TRACHEOPHYTA / VASCULAR PLANTS
Sporophyte long lived, cells polyplastidic, photosynthetic red light response, stomata open in response to blue light; plant homoiohydrous [water content of protoplasm relatively stable]; control of leaf hydration passive; plant endohydrous [physiologically important free water inside plant]; PIN[auxin efflux facilitators]-mediated polar auxin transport; (condensed or nonhydrolyzable tannins/proanthocyanidins +); borate cross-linked rhamnogalactan II, xyloglucans with side chains uncharged [?level], in secondary walls of vascular and mechanical tissue; lignins +; roots +, often ≤1 mm across, root hairs and root cap +; stem apex multicellular [several apical initials, no tunica], with cytohistochemical zonation, plasmodesmata formation based on cell lineage; vascular development acropetal, tracheids +, in both protoxylem and metaxylem, G- and S-types; sieve cells + [nucleus degenerating]; endodermis +; stomata numerous, involved in gas exchange; leaves +, vascularized, spirally arranged, blades with mean venation density ca 1.8 mm/mm2 [to 5 mm/mm2], all epidermal cells with chloroplasts; sporangia in strobili, sporangia adaxial, columella 0; tapetum glandular; sporophyte-gametophyte junction lacking dead gametophytic cells, mucilage, ?position of transfer cells; MTOCs not associated with plastids, basal body 350-550 nm long, stellate array in transition region initially joining microtubule triplets; archegonia embedded/sunken [only neck protruding]; embryo suspensor +, shoot apex developing away from micropyle/archegonial neck [from hypobasal cell, endoscopic], root lateral with respect to the longitudinal axis of the embryo [plant homorhizic].
[MONILOPHYTA + LIGNOPHYTA]Sporophyte growth ± monopodial, branching spiral; roots endomycorrhizal [with Glomeromycota], lateral roots +, endogenous; G-type tracheids +, with scalariform-bordered pits; leaves with apical/marginal growth, venation development basipetal, growth determinate; sporangium dehiscence by a single longitudinal slit; cells polyplastidic, MTOCs diffuse, perinuclear, migratory; blepharoplasts +, paired, with electron-dense material, centrioles on periphery, male gametes multiciliate; nuclear genome [1C] 7.6-10 pg [mode]; chloroplast long single copy ca 30kb inversion [from psbM to ycf2]; mitochondrion with loss of 4 genes, absence of numerous group II introns; LITTLE ZIPPER proteins.
LIGNOPHYTA†
Sporophyte woody; stem branching axillary, buds exogenous; lateral root origin from the pericycle; cork cambium + [producing cork abaxially], vascular cambium bifacial [producing phloem abaxially and xylem adaxially].
SEED PLANTS† / SPERMATOPHYTA†
Growth of plant bipolar [plumule/stem and radicle/root independent, roots positively geotropic]; plants heterosporous; megasporangium surrounded by cupule [i.e. = unitegmic ovule, cupule = integument]; pollen lands on ovule; megaspore germination endosporic, female gametophyte initially retained on the plant, free-nuclear/syncytial to start with, walls then coming to surround the individual nuclei, process proceeding centripetally.
EXTANT SEED PLANTS
Plant evergreen; nicotinic acid metabolised to trigonelline, (cyanogenesis via tyrosine pathway); microbial terpene synthase-like genes 0; primary cell walls rich in xyloglucans and/or glucomannans, 25-30% pectin [Type I walls]; lignin chains started by monolignol dimerization [resinols common], particularly with guaiacyl and p-hydroxyphenyl [G + H] units [sinapyl units uncommon, no Maüle reaction]; roots often ≥1 mm across, stele diarch to pentarch, xylem and phloem originating on alternating radii, cork cambium deep seated, gravitropism response fast; stem apical meristem complex [with quiescent centre, etc.], plasmodesma density in SAM 1.6-6.2[mean]/μm2 [interface-specific plasmodesmatal network]; eustele +, protoxylem endarch, endodermis 0; wood homoxylous, tracheids and rays alone, tracheid/tracheid pits circular, bordered; mature sieve tube/cell lacking functioning nucleus, sieve tube plastids with starch grains; phloem fibres +; cork cambium superficial; leaf nodes 1:1, a single trace leaving the vascular sympodium; leaf vascular bundles amphicribral; guard cells the only epidermal cells with chloroplasts, stomatal pore with active opening in response to leaf hydration, control by abscisic acid, metabolic regulation of water use efficiency, etc.; branching by axillary buds, exogenous; prophylls two, lateral; leaves with petiole and lamina, development basipetal, lamina simple; sporangia borne on sporophylls; spores not dormant; microsporophylls aggregated in indeterminate cones/strobili; grains monosulcate, aperture in ana- position [distal], primexine + [involved in exine pattern formation with deposition of sporopollenin from tapetum there], exine and intine homogeneous, exine alveolar/honeycomb; ovules with parietal tissue [= crassinucellate], megaspore tetrad linear, functional megaspore single, chalazal, sporopollenin 0; gametophyte ± wholly dependent on sporophyte, development initially endosporic [apical cell 0, rhizoids 0, etc.]; male gametophyte with tube developing from distal end of grain, male gametes two, developing after pollination, with cell walls; embryo cellular ab initio, suspensor short-minute, embryonic axis straight [shoot and root at opposite ends], primary root/radicle produces taproot [= allorhizic], cotyledons 2; embryo ± dormant; chloroplast ycf2 gene in inverted repeat, trans splicing of five mitochondrial group II introns, rpl6 gene absent; ??whole nuclear genome duplication [ζ/zeta duplication event], 2C genome size (0.71-)1.99(-5.49) pg, two copies of LEAFY gene, PHY gene duplications [three - [BP [A/N + C/O]] - copies], 5.8S and 5S rDNA in separate clusters.
IID. ANGIOSPERMAE / MAGNOLIOPHYTA
Lignans, O-methyl flavonols, dihydroflavonols, triterpenoid oleanane, apigenin and/or luteolin scattered, [cyanogenesis in ANA grade?], lignin also with syringyl units common [G + S lignin, positive Maüle reaction - syringyl:guaiacyl ratio more than 2-2.5:1], hemicelluloses as xyloglucans; root cap meristem closed (open); pith relatively inconspicuous, lateral roots initiated immediately to the side of [when diarch] or opposite xylem poles; epidermis probably originating from inner layer of root cap, trichoblasts [differentiated root hair-forming cells] 0, hypodermis suberised and with Casparian strip [= exodermis]; shoot apex with tunica-corpus construction, tunica 2-layered; starch grains simple; primary cell wall mostly with pectic polysaccharides, poor in mannans; tracheid:tracheid [end wall] plates with scalariform pitting, multiseriate rays +, wood parenchyma +; sieve tubes enucleate, sieve plates with pores (0.1-)0.5-10< µm across, cytoplasm with P-proteins, not occluding pores of plate, companion cell and sieve tube from same mother cell; ?phloem loading/sugar transport; nodes 1:?; dark reversal Pfr → Pr; protoplasm dessication tolerant [plant poikilohydric]; stomata randomly oriented, brachyparacytic [ends of subsidiary cells ± level with ends of guard cells], outer stomatal ledges producing vestibule, reduction in stomatal conductance with increasing CO2 concentration; lamina formed from the primordial leaf apex, margins toothed, development of venation acropetal, overall growth ± diffuse, secondary veins pinnate, fine venation hierarchical-reticulate, (1.7-)4.1(-5.7) mm/mm2, vein endings free; flowers perfect, pedicellate, ± haplomorphic, protogynous; parts free, numbers variable, development centripetal; P = T, petal-like, each with a single trace, outer members not sharply differentiated from the others, not enclosing the floral bud; A many, filament not sharply distinguished from anther, stout, broad, with a single trace, anther introrse, tetrasporangiate, sporangia in two groups of two [dithecal], each theca dehiscing longitudinally by a common slit, ± embedded in the filament, walls with at least outer secondary parietal cells dividing, endothecium +, cells elongated at right angles to long axis of anther; tapetal cells binucleate; microspore mother cells in a block, microsporogenesis successive, walls developing by centripetal furrowing; pollen subspherical, tectum continuous or microperforate, ektexine columellate, endexine restricted to the apertural regions, thin, compact, intine in apertural areas thick, orbicules +, pollenkitt +; nectary 0; carpels present, superior, free, several, spiral, ascidiate [postgenital occlusion by secretion], stylulus at most short [shorter than ovary], hollow, cavity not lined by distinct epidermal layer, stigma ± decurrent, carinal, dry; suprastylar extragynoecial compitum +; ovules few [?1]/carpel, marginal, anatropous, bitegmic, micropyle endostomal, outer integument 2-3 cells across, often largely subdermal in origin, inner integument 2-3 cells across, often dermal in origin, parietal tissue 1-3 cells across, nucellar cap?; megasporocyte single, hypodermal, functional megaspore lacking cuticle; female gametophyte lacking chlorophyll, four-celled [one module, egg and polar nuclei sisters]; ovule not increasing in size between pollination and fertilization; pollen grains bicellular at dispersal, germinating in less than 3 hours, siphonogamy, pollen tube unbranched, growing towards the ovule, between cells, growth rate (ca 10-)80-20,000 µm h-1, tube apex of pectins, wall with callose, lumen with callose plugs, penetration of ovules via micropyle [porogamous], whole process takes ca 18 hours, distance to first ovule 1.1-2.1 mm; male gametophytes tricellular, gametes 2, lacking cell walls, ciliae 0, double fertilization +, ovules aborting unless fertilized; fruit indehiscent, P deciduous; mature seed much larger than fertilized ovule, small [<5 mm long], dry [no sarcotesta], exotestal; endosperm +, ?diploid [one polar nucleus + male gamete], cellular, development heteropolar [first division oblique, micropylar end initially with a single large cell, divisions uniseriate, chalazal cell smaller, divisions in several planes], copious, oily and/or proteinaceous, embryo short [<¼ length of seed]; plastid and mitochondrial transmission maternal; Arabidopsis-type telomeres [(TTTAGGG)n]; nuclear genome [2C] (0.57-)1.45(-3.71) [1 pg = 109 base pairs], ??whole nuclear genome duplication [ε/epsilon event]; ndhB gene 21 codons enlarged at the 5' end, single copy of LEAFY and RPB2 gene, knox genes extensively duplicated [A1-A4], AP1/FUL gene, palaeo AP3 and PI genes [paralogous B-class genes] +, with "DEAER" motif, SEP3/LOFSEP and three copies of the PHY gene, [PHYB [PHYA + PHYC]]; chloroplast IR expansions, chlB, -L, -N, trnP-GGG genes 0.
[NYMPHAEALES [AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]]: wood fibres +; axial parenchyma diffuse or diffuse-in-aggregates; pollen monosulcate [anasulcate], tectum reticulate-perforate [here?]; ?genome duplication; "DEAER" motif in AP3 and PI genes lost, gaps in these genes.
[AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]: phloem loading passive, via symplast, plasmodesmata numerous; vessel elements with scalariform perforation plates in primary xylem; essential oils in specialized cells [lamina and P ± pellucid-punctate]; tension wood + [reaction wood: with gelatinous fibres, G-fibres, on adaxial side of branch/stem junction]; anther wall with outer secondary parietal cell layer dividing; tectum reticulate; nucellar cap + [character lost where in eudicots?]; 12BP [4 amino acids] deletion in P1 gene.
[MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]] / MESANGIOSPERMAE: benzylisoquinoline alkaloids +; sesquiterpene synthase subfamily a [TPS-a] [?level], polyacetate derived anthraquinones + [?level]; outer epidermal walls of root elongation zone with cellulose fibrils oriented transverse to root axis; P more or less whorled, 3-merous [?here]; pollen tube growth intra-gynoecial; extragynoecial compitum 0; carpels plicate [?here]; embryo sac monosporic [spore chalazal], 8-celled, bipolar [Polygonum type], antipodal cells persisting; endosperm triploid.
[CERATOPHYLLALES + EUDICOTS]: ethereal oils 0 [or next node up]; fruit dry [very labile].
EUDICOTS: (Myricetin +), asarone 0 [unknown in some groups, + in some asterids]; root epidermis derived from root cap [?Buxaceae, etc.]; (vessel elements with simple perforation plates in primary xylem); nodes 3:3; stomata anomocytic; flowers (dimerous), cyclic; protandry common; K/outer P members with three traces, ("C" +, with a single trace); A ?, filaments fairly slender, anthers basifixed; microsporogenesis simultaneous, pollen tricolpate, apertures in pairs at six points of the young tetrad [Fischer's rule], cleavage centripetal, wall with endexine; G with complete postgenital fusion, stylulus/style solid [?here], short [<2 x length of ovary]; seed coat?; palaeotetraploidy event.
[PROTEALES [[TROCHODENDRALES + BUXALES] [CORE EUDICOTS]]]: (axial/receptacular nectary +).
[TROCHODENDRALES + BUXALES] [CORE EUDICOTS]]: benzylisoquinoline alkaloids 0; euAP3 + TM6 genes [duplication of paleoAP3 gene: B class], mitochondrial rps2 gene lost.
[[TROCHODENDRALES + BUXALES] CORE EUDICOTS]: mitochondrial rps11 gene lost.
Age. This node dates to 121-117 Ma (Anderson et al. 2005) or ca 123.9 Ma (Tank et al. 2015: Table S2); Magallón and Castillo (2009) suggest an age of around ca 121.5 Ma, Foster et al. (2016a) an age of ca 135 Ma, Zeng et al. (2017) an age of 141-136 Ma, and ca 182 Ma is the age in Z. Wu et al. (2014). Ages in Vekemans et al. (2012) were about (123-)122.4(-121.4) Ma, in Magallón et al. (2015) around 128.6 Ma, in Wikström et al. (2003) about (142-)137, 124(-119) Ma - but c.f. sister groups there: [B [T + CE]], and Xue et al. (2012) ca 115.4 Ma.
Evolution: Divergence & Distribution. See M.-Y. Zhang et al. (2017) for pollen evolution.
Genes & Genomes. Adams et al. (2002b; see also Lei et al. 2013; Sanchez-Puerta et al. 2016) note that the mitochondrial rps11 gene is absent from all Pentapetalae sampled, including Vitis, Dillenia was not sampled, Buxaceae were (they lacked the gene) as were Trochodendraceae (they had the gene).
Just the one family, 7 genera, 120 species.
Note: In all node characterizations, boldface denotes a possible apomorphy, (....) denotes a feature the exact status of which in the clade is uncertain, [....] includes explanatory material; other text lists features found pretty much throughout the clade. However, the precise node to which many characters, particularly the more cryptic ones, should be assigned is unclear. This is partly because homoplasy is very common, in addition, basic information for all too many characters is very incomplete, frequently coming from taxa well embedded in the clade of interest and so making the position of any putative apomorphy uncertain. Then there are the not-so-trivial issues of how character states are delimited and ancestral states are reconstructed (see above).
Age. Wikström et al. (2001) thought that crown-group Buxaceae were (130-)124, 113(-109) Ma, Magallón and Castillo (2009) suggested an age of ca 111.5 Ma, Magallón et al. (2013) an age of about 118.4 Ma, and Bell et al. (2010) an age of (103-)99, 98(-97) Ma. Anderson et al. (2005) suggested that this group was some 99 Ma old, while the age in Magallón et al. (2015) was only about 85.1 Ma.
Drinnan et al. (1991) described Spanomera mauldinensis from east North American deposits laid down 113-98 Ma (Albian); it is perhaps a member of stem group Buxaceae (see also Crepet et al. 2004; Doyle & Endress 2010; Doyle & Upchurch 2014). The carpellate flowers are dimerous, but there may be five stamens opposite five tepals, the pollen is tricolpate, and the fruit is follicular (Drinnan et al. 1991); relationships with Trochodendraceae were also suggested (Doyle 1999). Recent work suggests that it is in the Buxales-Gunnerales area, whether in their stem or crown groups or in an independant lineage (Schönenberger et al. 2020), and Buxales were possible associates in all the analyses carried out by López-Martínez et al. (2023: Table 3), although various other orders were also suggested. Lusistemon, also with striate, tricolpate pollen (but with different anther insertion and arrangement), from deposits in Portugal of about the same age, seems to be related (Pedersen et al. 2007). See also Friis et al. (2011) for fossils.
Phylogeny. For the phylogenetic position of Buxales, see the Ranunculales page.
Includes Buxaceae.
Synonymy: Didymelales Takhtajan - Buxineae J. Presl - Buxanae Reveal & Doweld
BUXACEAE Dumortier, nom. cons. - Back to Buxales
Evergreen, woody; steroidal pregnan pseudoalkaloids +; true tracheids +; cortex with sclereids/fibers; nodes?; petiole with three arcuate traces; flowers imperfect, stomata cyclocytic; lamina margins entire; flowers small [7> mm across], imperfect; P members with a single trace; anthers ± dorsifixed; styluli stigmatic ± their entire length, strongly canaliculate, unicellular-papillate; ovules 1-2/carpel; micropyle bistomal; seed testal, with several cell layers; endosperm development?, embryo size?; x = 14 (?13, ?7), nuclear genome [1 C] (0.017-)1.268(-95.262) pg.
7 [list]/120: four groups below. ± Temperate to ± tropical, very scattered, not in the Antipodes.
Age. Cathiaria japonica has been found in rocks ca 89 Ma from Japan and it has been placed in Buxaceae s.l. (Takahashi et al. 2017); there is a single carpel subtended by a single bract and tricolpate pollen.
1. Didymeles Leandri —— Synonymy: Didymelaceae Leandri
Cork ?; pericyclic fibres 0; pith chambered; petiole with the two lateral traces inverted; hairs small, peltate; leaves spiral; plant dioecious; staminate flowers: P 0; A 2, subsessile, connate basally, anthers extrorse; pollen colpi 2-orate, surface reticulate, with small spines; pistillode 0; carpelate flowers: ?P 1, scale-like; staminode 0; G 1, compitum necessarily 0; ovules median, epitropous, integuments ± elongated, protruding into the stylar canal, outer integument fringed apically; fruit a drupelet furrowed on one side; rudimentary aril +?; testa cell walls thickened, tegmen obliterated; endosperm 0, development?, embryo large; n = ?
1/2. E. Madagascar, the Comoros. [Photos - Inflorescence.]
[Stylocerateae [Haptanthus + Buxeae]]: raffinose, stachyose + [phloem exudate], tannins 0 [even in flowers]; cork subepidermal or pericyclic; sieve tube plastids with protein crystalloids; nodes 3:3; secretory cells single or in branching rows; cuticle waxes as curled rodlets or irregular platelets; (stomata laterocytic); leaves opposite, lamina ptyxis flat to curved; plant monoecious, inflorescence with a terminal carpelate flower, botyroid; staminate flowers: P (0-)2-4(5), not clearly separated from bracteoles, decussate; tapetal cells binucleate; pistillode +, nectariferous; carpelate flowers: P 6-20, at least some spiral; G [3], completely fused, with five bundles, a roof over the ovary, so styluli ± marginal, spreading, stigma dry or weakly secretory; compitum 0; ovules apotropous, outer integument 5-8 cells across, inner integument 2(-3) cells across, parietal tissue 4-15+ cell layers across, (weak nucellar cap), ovular obturator +, hypostase +; fruit with persistent styles; (seed carunculate), exotesta lignified, palisade, hypodermis often lignified; endosperm copious, (perisperm +); x = ?7.
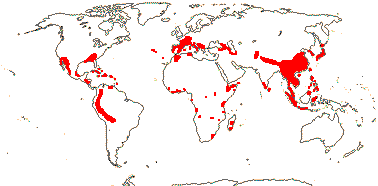
6/115. ± Temperate to ± tropical, very scattered, not the Antipodes. Map: see Meusel et al. (1978) and Trop. Afr. Fl. Pl. Ecol. Distr. vol. 5 (2010). Photo: Inflorescence, Fruits.
Age. Anderson et al. (2005) suggested that this node was some 111-63 Ma old; ages in N. Zhang et al. (2012) were ca 45 Ma and in Wikström et al. (2001) (107-)100, 93(-86) Ma.
2. Stylocerateae Pax —— Synonymy: Pachysandraceae J. Agardh, Stylocerataceae Reveal & Hoogland
(Perennial rhizomatous herbs); (nodes 1:3 - Sarcococca); leaves spiral, lamina margins toothed, (venation triplinerved); (plant dioecious), (carpelate flowers basal in the inflorescence); staminate flowers: A (many, sessile - Styloceras), filaments conspicuous, (endothecium >2-layers); pollen pantoporate, (exine crotonoid, reticulate, small spines - Sarcococca); (pistillode 0); carpelate flowers: bracts many, all spiral; G also [2], not nectarifeous [?all], (loculi divided); ovules epitropous, micropyle bistomal, outer integument 2-3 cells across; fruit a drupe (dry; ?dehiscent, pulp surrounding blue seeds - Styloceras); testa multiplicative, endotegmen lignified [Sarcococca]; endosperm nuclear [Sarcococca], embryo with short radicle; n = 12-14.
3/22: Sarcococca (11). E. Asia to W. Malesia, E. North America, N. South America.
[Haptanthus + Buxeae] (if this clade exists): G [3]; outer integument ca 5 cells across.
3. Haptanthus hazlettii Goldberg & C. Nelson —— Synonymy: Haptanthaceae C. Nelson
Cork ?; fibres storied; vessel elements with scalariform perforation plates; nodes 1:1 [from bracts]; stomata ?; plant glabrous; lamina weakly pli-nerved at the base; inflorescences with staminate flowers borne decussately along the branches, (terminal staminate flower +), bracteoles 0; staminate flowers: P 1; A 2, completely adnate to P, dehiscence ?valvate, endothecium >2-layered; pollen tricolpate; pistillode 0; carpelate flowers: P 0; staminodes 0; G stipitate, nectary 0, symplicate, placentation parietal, no roof over the ovary, styles radiating; ovules 8-15/carpel, details?; fruit septicidal; seeds rugose; n = ?
1/1. Central America, Honduras; recently recollected.
4. Buxeae Dumortier
Reaction wood on lower side of stem [Buxus], (cortical vascular bundles in angles of stems); (giant/D-type stomata on veins - Buxus); (leaves spiral), lamina (venation dendritic [cladodromous] - ?level); staminate flowers: A 4-10, sessile; pollen 3-12 colporate, colpus 2-4 orate/12+ pantoporate, surface reticulate/substriate, pilate-verrucate; (pistillode 0); carpelate flowers: inflorescence with basal decussate "bracts"; G ([2]), median carpel adaxial, nectary +, between styles (0); ovules apotropous, micropyle endostomal, outer integument 5-10 cells across, ?minute third integument, suprachalazal nucellus massive [Buxus]; fruit an explosively-dehiscent capsule, loculicidal; seed shiny; embryo ± curved, radicle long; n = 10, 14.
2/95: Buxus (90). Scattered, inc. Madagasacar, nearly all Central America and West Indies in New World, not Australia nor much of Malesia.
Evolution: Divergence & Distribution. There are a number of distinctive tricolpate-striate pollen types perhaps assignable to plants of this relationship (?Buxeae?) that are widely distributed in rocks back to the early Cretaceous (e.g. Doyle 1999; Pedersen et al. 2007 and references). Krutzsch (1989) discussed the later pollen record of the family, which is widespread in the northern hemisphere in the early Caenozoic. Fossil Didymeles is reported from New Zealand (Lee at al. 2001) and elsewhere in the southern hemisphere (Friis et al. 2011; c.f. Carpenter et al. 2010).
"Pistillode +, nectariferous" is provisionally placed as an apomorphy of [Stylocerateae, Haptanthus, Buxeae]; Köhler (2006) noted that in two of the three genera of the Sarcococceae there was a pistillode, and also that the nectary was central.
Ecology & Physiology. 17/34 of the Cuban species of Buxus are reported to accumulate nickel (Reeves et al. 1996; Brooks 1998) - species limits? Gutiérrez et al. (2023) discuss the evolution of these plants; they seem to have originated in Mexico and became nickel hyperaccumulators on Cuba, where they diversified and whence non-nickel accumulators moved to other Caribbean islands and to northern South America.
Reaction wood with rather odd features for that of angiosperms is reported from Buxus (Nawawi et al. 2016 and references), and this is discussed elsewhere.
Pollination Biology & Seed Dispersal. Members of Buxeae are myrmecochorous (Lengyel et al. 2010).
Chemistry, Morphology, etc.. Iridoids are reported from Didymeles by Sutton (1989); this should be confirmed.
For rather odd There is a broad pith and at best late-developing cork (Leandri 1937); the axial parenchyma is replaced by lignified cells (Takhtajan 1997). Harms (1893) described sclereids in the pericyclic position in Didymeles, and also cortical sclereids. Buxus often has fibre strands in the stem. All genera (Styloceras unknown; Hapt.) have three vascular bundles in the upper part of the petiole. The stomata of some Buxaceae may be described as being laterocytic (Baranova 1983).
The floral morphology of Didymeles has been described by von Balthazar and Endress (2002a) and Von Balthazar et al. (2003). In carpelate flowers there is a little scale between the ovule and the bract; its morphological nature is unclear; is it a third integument (Endress 2011b)? The flowers may be borne in pairs with a small hump between them, perhaps a modified cymose unit, the hump being a reduced terminal flower.
Haptanthus has parietal placentation and odd staminate flowers that were originally described as consisting of a single stamen (Goldberg & Nelson S. 1989). However, there are two stamens each with two thecae attached their length to an elliptic structure with a single arcuate vascular bundle (?very broad filament; ?prophyll, ?perianth member - the former interpretation is preferred interpretation by Shipunov & Oskolski 2011, the latter by Oskolski et al. 2015); the endothecium is massive (Doust & Stevens 2005), as it is in Styloceras, the genus perhaps most similar to Haptanthus (Shipunov & Oskolski 2011). The inflorescence is distinctive, and is described as a botryoid with a central terminal flower (Oskolski et al. 2015).
The carpelate flowers of other taxa have spirally inserted parts, but the basical floral construction of the family is perhaps dimerous (see also Oskolski et al. 2015). Interpretation of inflorescence morphology is not easy, indeed, the nature of the paired "bracts" (?prophylls?) below the carpelate flowers of Buxus is particularly unclear (von Balthazar & Endress 2001, 2002a). The perianth parts have a single vascular bundle (von Balthazar and Endress 2002a). Some species of Buxus have distinctive pollen with two or more orae per colpus that is rather like that of Didymeles; pollen in the family is very variable (Köhler 2006 for references). Both Styloceras and Pachysandra have subdivided carpels; the relative positions of the carpels and perianth is unclear (Wiger 1935).
For general information, see van Tieghem (1897a) and Köhler (2006: as Buxaceae and Didymelaceae): for reaction (compression-type) wood in Buxus, see Timell (1986, vol.1, chap. 8) and references in Aiso et al. (2016), for nectary types, see Vogel (1998c), for pollen of Buxus, very variable, see Köhler (1994 and references), for floral morphology, von Balthazar and Endress (2001, esp. 2002b), and for ovules, etc., see Wiger (1935; also Mauritzon 1935; Wiger 1936; Bhatnagar & Kapil 1994).
Phylogeny. Didymeles is sister to Buxaceae s. str. (representatives of Buxeae and Sarcococceae were included) in the three-gene analysis of Worberg et al. (2007), although Hilu et al. (2003: matK [incomplete sequence] alone) found Didymeles to be embedded in a paraphyletic Buxaceae. The poorly-known Haptanthaceae were tentatively included in Buxales in pre-March 2011 versions of the site based on the anatomy of the staminate flower (Doust & Stevens 2005), but a position within Buxaceae s.l. was strongly suggested by analysis of data from the rbcL gene, although the exact position differed in maximum likelihood and maximum parsimony analyses (Shipunov & Shipunova 2011). For relationships in the family, see also von Balthazar et al. (2000) and von Balthazar and Endress (2002b), and relationships within Stylocerateae are [Sarcococca [Styloceras + Pachysandra]], although Soh et al. (2014) recovered the relationships [[Sarcococca + Styloceras] Pachysandra].
Classification. Including Didymelaceae in Buxaceae was an option in A.P.G. II (2003);the two were combined in A.P.G. III (2009).
Previous Relationships. Buxaceae have often been associated with Euphorbiaceae s.l. (e.g. Cronquist 1981). When the very distinctive Haptanthus was first described, it was not assigned to a family. Shipunov (2003) placed Haptanthaceae in his Violales (= Malpighiales here), close to Salicaceae and Lacistemataceae (see also Goldberg & Alden 2005), probably because of its parietal placentation.
){kind=link}