EMBRYOPSIDA Pirani & Prado
Gametophyte dominant, independent, multicellular, initially ±globular; showing gravitropism; acquisition of phenylalanine lysase [PAL], phenylpropanoid metabolism [lignans +, flavonoids + (absorbtion of UV radiation)], xyloglucans in primary cell wall with distinctive side chains; plant poikilohydrous [protoplasm dessication tolerant], ectohydrous [free water outside plant physiologically important]; thalloid, leafy, with single-celled apical meristem, tissues little differentiated, rhizoids +, unicellular; chloroplasts several per cell, pyrenoids 0; glycolate metabolism in leaf peroxisomes [glyoxysomes]; centrioles in vegetative cells 0, metaphase spindle anastral, preprophase band of microtubules [where cell plate will join parental cell wall], phragmoplast + [cell wall deposition spreading from around the spindle fibres], plasmodesmata +; antheridia and archegonia jacketed, surficial; blepharoplast +, coaxial bicentriole pair separates at midpoint, centrioles rotate, associated with basal bodies of cilia, multilayered structure + [4 layers: L1, L4, tubules; L2, L3, short vertical lamellae], spline + [tubules from L1 encircling spermatid], basal body 200-250 nm long, associated with amorphous electron-dense material, microtubules in basal end lacking symmetry, stellate array of filaments in transition zone extended, axonemal cap 0 [microtubules disorganized at apex of cilium]; male gametes [spermatozoids] with a left-handed coil, cilia 2, lateral; oogamy; sporophyte multicellular, cuticle +, plane of first cell division horizontal [with respect to long axis of archegonium/embryo sac], early embryo developing towards the archegonial neck [from epibasal cell, exoscopic], with at least transient apical cell [?level], initially surrounded by and dependent on gametophyte, placental transfer cells +, in both sporophyte and gametophyte, development of wall ingrowths early, suspensor/foot +, cells at foot tip somewhat haustorial; sporangium +, single, terminal, dehiscence longitudinal; meiosis sporic, monoplastidic, MTOC [MTOC = microtubule organizing centre] associated with plastid, cytokinesis simultaneous, preceding nuclear division, sporocytes 4-lobed, with a quadripolar microtubule system; wall development both centripetal and centrifugal, sporopollenin + laid down in association with trilamellar layers [white-line centred lamellae; tripartite lamellae], >1000 spores/sporangium; nuclear genome size <1.4 pg, main telomere sequence motif TTTAGGG, LEAFY and KNOX1 and KNOX2 genes present, ethylene involved in cell elongation; chloroplast genome with close association between trnLUAA and trnFGAA genes, precursor for starch synthesis in plastid.
Many of the bolded characters in the characterization above are apomorphies of subsets of streptophytes along the lineage leading to the embryophytes, not apomorphies of crown-group embryophytes per se.
All groups below are crown groups, nearly all are extant. Characters mentioned are those of the immediate common ancestor of the group, [] contains explanatory material, () features common in clade, exact status unclear.
STOMATOPHYTES
Abscisic acid, L- and D-methionine distinguished metabolically; sporophyte with polar transport of auxins, class 1 KNOX genes expressed in sporangium alone; sporangium wall 4≤ cells across [≡ eusporangium], tapetum +, secreting sporopollenin, which obscures outer white-line centred lamellae, columella +, developing from endothecial cells; stomata +, on sporangium, anomocytic, cell lineage that produces them with symmetric divisions [perigenous]; underlying similarities in the development of conducting tissue and of rhizoids/root hairs; spores trilete; shoot meristem patterning gene families expressed; MIKC, MI*K*C* and class 1 and 2 KNOX genes, post-transcriptional editing of chloroplast genes; gain of three group II mitochondrial introns, mitochondrial trnS(gcu) and trnN(guu) genes 0.
[Anthocerophyta + Polysporangiophyta]: xyloglucans in the primary cell wall with fucosylated subunits; archegonia embedded/sunken; sporophyte long-lived, chlorophyllous; cell walls with xylans.
POLYSPORANGIOPHYTA†
Sporophyte dominant, branched, branching apical, dichotomous, potentially indeterminate; vascular tissue +; stomata on stem; sporangia several, each opening independently; spore walls not multilamellate [?here].
EXTANT TRACHEOPHYTA / VASCULAR PLANTS
Sporophyte with photosynthetic red light response; (condensed or nonhydrolyzable tannins/proanthocyanidins +); xylans in secondary walls of vascular and mechanical tissue; root hairs +; lignins +; tracheids +, in both protoxylem and metaxylem, G- and S-types; sieve cells + [nucleus degenerating]; plant homoiohydrous [water content of protoplasm relatively stable]; control of leaf hydration passive; plant endohydrous [physiologically important free water inside plant]; endodermis +; leaves/sporophylls spirally arranged, blades with mean venation density 1.8 mm/mm2 [to 5 mm/mm2], all epidermal cells with chloroplasts; sporangia adaxial, columella 0; tapetum glandular; ?position of transfer cells; MTOCs not associated with plastids, basal body 350-550 nm long, stellate array in transition region initially joining microtubule triplets; root lateral with respect to the longitudinal axis of the embryo [plant homorhizic].
[MONILOPHYTA + LIGNOPHYTA]Sporophyte endomycorrhizal [with Glomeromycota]; root cap +, protoxylem exarch, lateral roots +, endogenous; stem apex multicellular, G-type tracheids +, with scalariform-bordered pits; leaves with apical/marginal growth, venation development basipetal, growth determinate; sporangium dehiscence by a single longitudinal slit; cells polyplastidic, MTOCs diffuse, perinuclear; blepharoplasts +, paired, with electron-dense material, centrioles on periphery, male gametes multiciliate; chloroplast long single copy ca 30kb inversion [from psbM to ycf2]; LITTLE ZIPPER proteins.
LIGNOPHYTA†
Sporophyte woody; stem branching lateral, meristems axillary; lateral root origin from the pericycle; cork cambium + [producing cork abaxially], vascular cambium bifacial [producing phloem abaxially and xylem adaxially].
SEED PLANTS†
Plants heterosporous; megasporangium surrounded by cupule [i.e. = unitegmic ovule, cupule = integument]; pollen lands on ovule; megaspore germination endosporic [female gametophyte initially retained on the plant].
EXTANT SEED PLANTS / SPERMATOPHYTA
Plant evergreen; nicotinic acid metabolised to trigonelline, (cyanogenesis via tyrosine pathway); primary cell walls rich in xyloglucans and/or glucomannans, 25-30% pectin [Type I walls]; lignins particularly with guaiacyl and p-hydroxyphenyl [G + H] units [sinapyl units uncommon, no Maüle reaction]; root stele with xylem and phloem originating on alternate radii, cork cambium deep seated; stem apical meristem complex [with quiescent centre, etc.], mitochondrial density in SAM 1.6-6.2[mean]/μm2 [interface-specific mitochondrial network]; eustele +, protoxylem endarch, endodermis 0; wood homoxylous, tracheids and rays alone, tracheid/tracheid pits circular, bordered; mature sieve tube/cell lacking functioning nucleus, sieve tube plastids with starch grains; phloem fibres +; cork cambium superficial; leaf nodes 1:1, a single trace leaving the vascular sympodium; leaf vascular bundles amphicribral; guard cells the only epidermal cells with chloroplasts, stomatal pore with active opening in response to leaf hydration, control by abscisic acid, metabolic regulation of water use efficiency, etc.; axillary buds +, exogenous; prophylls two, lateral; leaves with petiole and lamina, development basipetal, lamina simple; sporangia borne on sporophylls; spores not dormant; microsporophylls aggregated in indeterminate cones/strobili; grains monosulcate, aperture in ana- position [distal], primexine + [involved in exine pattern formation with deposition of sporopollenin from tapetum there], exine and intine homogeneous, exine alveolar/honeycomb; megasporangium indehiscent; ovules with parietal tissue 2+ cells across, megaspore tetrad linear, functional megaspore single, chalazal, sporopollenin 0; gametophyte ± wholly dependent on sporophyte, development initially endosporic [apical cell 0, rhizoids 0, etc.]; male gametophyte with tube developing from distal end of grain, male gametes two, developing after pollination, with cell walls; female gametophyte initially syncytial, walls then surrounding individual nuclei; embryo cellular ab initio, plane of first cleavage of zygote transverse, shoot apex developing away from micropyle [i.e. away from archegonial neck; from hypobasal cell, endoscopic], suspensor +, short-minute, embryonic axis straight [shoot and root at opposite ends; plant allorhizic], cotyledons 2; embryo ± dormant; plastid transmission maternal; ycf2 gene in inverted repeat, whole nuclear genome duplication [ζ - zeta - duplication], two copies of LEAFY gene, PHY gene duplications [three - [BP [A/N + C/O]] - copies], nrDNA with 5.8S and 5S rDNA in separate clusters; mitochondrial trans- nad2i542g2 and coxIIi3 introns present.
ANGIOSPERMAE / MAGNOLIOPHYTA
Lignans, O-methyl flavonols, dihydroflavonols, triterpenoid oleanane, apigenin and/or luteolin scattered, [cyanogenesis in ANA grade?], lignin also with syringyl units common [G + S lignin, positive Maüle reaction - syringyl:guaiacyl ratio more than 2-2.5:1], hemicelluloses as xyloglucans; root apical meristem intermediate-open; stele di- to pentarch [oligarch], pith relatively inconspicuous, lateral roots arise opposite or immediately to the side of [when diarch] xylem poles; origin of epidermis with no clear pattern [probably from inner layer of root cap], trichoblasts [differentiated root hair-forming cells] 0, hypodermis suberised and with Casparian strip [= exodermis]; shoot apex with tunica-corpus construction, tunica 2-layered; starch grains simple; primary cell wall mostly with pectic polysaccharides, poor in mannans; tracheid:tracheid [end wall] plates with scalariform pitting, wood parenchyma +; sieve tubes enucleate, sieve plate with pores (0.1-)0.5-10< µm across, cytoplasm with P-proteins, not occluding pores of plate, companion cell and sieve tube from same mother cell; sugar transport in phloem passive; nodes 1:?; stomata brachyparacytic [ends of subsidiary cells level with ends of pore], outer stomatal ledges producing vestibule, reduction in stomatal conductance with increasing CO2 concentration; lamina formed from the primordial leaf apex, margins toothed, development of venation acropetal, overall growth ± diffuse, secondary veins pinnate, fine venation hierarchical-reticulate, (1.7-)4.1(-5.7) mm/mm2, vein endings free; flowers perfect, pedicellate, ± haplomorphic, protogynous; parts free, numbers variable, development centripetal; P +, ?insertion, members each with a single trace, outer members not sharply differentiated from the others, not enclosing the floral bud; A many, filament not sharply distinguished from anther, stout, broad, with a single trace, anther introrse, tetrasporangiate, sporangia in two groups of two [dithecal], each theca dehiscing longitudinally by a common slit, ± embedded in the filament, walls with at least outer secondary parietal cells dividing, endothecium +, cells elongated at right angles to long axis of anther; tapetal cells binucleate; microspore mother cells in a block, microsporogenesis successive, walls developing by centripetal furrowing; pollen subspherical, tectum continuous or microperforate, ektexine columellate, endexine lamellate only in the apertural regions, thin, compact, intine in apertural areas thick, pollenkitt +; nectary 0; carpels present, superior, free, several, ascidiate [postgenital occlusion by secretion], stylulus at most short [shorter than ovary], hollow, cavity not lined by distinct epidermal layer, stigma ± decurrent, carinal, dry, extragynoecial compitum +; ovules few [?1]/carpel, marginal, anatropous, bitegmic, micropyle endostomal, outer integument 2-3 cells across, often largely subdermal in origin, inner integument 2-3 cells across, often dermal in origin, parietal tissue 1-3 cells across [crassinucellate], nucellar cap?; megasporocyte single, hypodermal, functional megaspore lacking cuticle; female gametophyte lacking chlorophyll, not photosynthesising, four-celled [one module, nucleus of egg cell sister to one of the polar nuclei]; ovule not increasing in size between pollination and fertilization; pollen grains land on stigma, bicellular at dispersal, mature male gametophyte tricellular, germinating in less than 3 hours, pollen tube elongated, unbranched, growing between cells, growth rate (20-)80-20,000 µm/hour, apex of pectins, wall with callose, lumen with callose plugs, penetration of ovules via micropyle [porogamous], whole process takes ca 18 hours, distance to first ovule 1.1-2.1 mm; male gametes lacking cell walls, ciliae 0, siphonogamy; double fertilization +, ovules aborting unless fertilized; P deciduous in fruit; mature seed much larger than fertilized ovule, small [], dry [no sarcotesta], exotestal; endosperm +, cellular, development heteropolar [first division oblique, micropylar end initially with a single large cell, divisions uniseriate, chalazal cell smaller, divisions in several planes], copious, oily and/or proteinaceous, embryo short [<¼ length of seed]; dark reversal Pfr → Pr; Arabidopsis-type telomeres [(TTTAGGG)n]; nuclear genome very small [1C = <1.4 pg, 1 pg = 109 base pairs], whole nuclear genome duplication [ε/epsilon - duplication]; protoplasm dessication tolerant [plant poikilohydric]; ndhB gene 21 codons enlarged at the 5' end, single copy of LEAFY and RPB2 gene, knox genes extensively duplicated [A1-A4], AP1/FUL gene, palaeo AP3 and PI genes [paralogous B-class genes] +, with "DEAER" motif, SEP3/LOFSEP and three copies of the PHY gene, [PHYB [PHYA + PHYC]]; chloroplast chlB, -L, -N, trnP-GGG genes 0.
[NYMPHAEALES [AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]]: wood fibres +; axial parenchyma diffuse or diffuse-in-aggregates; pollen monosulcate [anasulcate], tectum reticulate-perforate [here?]; ?genome duplication; "DEAER" motif in AP3 and PI genes lost, gaps in these genes.
[AUSTROBAILEYALES [MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]]]: vessel elements with scalariform perforation plates in primary xylem; essential oils in specialized cells [lamina and P ± pellucid-punctate]; tension wood + [reaction wood: with gelatinous fibres, g-fibres, on adaxial side of branch/stem junction]; tectum reticulate; anther wall with outer secondary parietal cell layer dividing; nucellar cap + [character lost where in eudicots?]; 12BP [4 amino acids] deletion in P1 gene.
[MONOCOTS [[CHLORANTHALES + MAGNOLIIDS] [CERATOPHYLLALES + EUDICOTS]]] / MESANGIOSPERMAE: benzylisoquinoline alkaloids +; sesquiterpene synthase subfamily a [TPS-a] [?level], polyacetate derived anthraquinones + [?level]; outer epidermal walls of root elongation zone with cellulose fibrils oriented transverse to root axis; P more or less whorled, 3-merous [?here]; pollen tube growth intra-gynoecial [extragynoecial compitum 0]; carpels plicate [?here]; embryo sac bipolar, 8 nucleate, antipodal cells persisting; endosperm triploid.
[MONOCOTS [CERATOPHYLLALES + EUDICOTS]]: (extra-floral nectaries +); (veins in lamina often 7-17 mm/mm2 or more [mean for eudicots 8.0]); (stamens opposite [two whorls of] P); (pollen tube growth fast).
[CERATOPHYLLALES + EUDICOTS]: ethereal oils 0.
EUDICOTS: (Myricetin, delphinidin +), asarone 0 [unknown in some groups, + in some asterids]; root epidermis derived from root cap [?Buxaceae, etc.]; (vessel elements with simple perforation plates in primary xylem); nodes 3:3; stomata anomocytic; flowers (dimerous), cyclic; protandry common; K/outer P members with three traces, ("C" +, with a single trace); A ?, filaments fairly slender, anthers basifixed; microsporogenesis simultaneous, pollen tricolpate, apertures in pairs at six points of the young tetrad [Fischer's rule], cleavage centripetal, wall with endexine; G with complete postgenital fusion, stylulus/style solid [?here]; seed coat?
[PROTEALES [TROCHODENDRALES [BUXALES + CORE EUDICOTS]]]: (axial/receptacular nectary +).
[TROCHODENDRALES [BUXALES + CORE EUDICOTS]]: benzylisoquinoline alkaloids 0; euAP3 + TM6 genes [duplication of paleoAP3 gene: B class], mitochondrial rps2 gene lost.
[BUXALES + CORE EUDICOTS]: ?
CORE EUDICOTS / GUNNERIDAE: (ellagic and gallic acids +); leaf margins serrate; compitum + [one position]; micropyle?; γ whole nuclear genome duplication [palaeohexaploidy, gamma triplication], PI-dB motif +, small deletion in the 18S ribosomal DNA common.
[ROSIDS ET AL. + ASTERIDS ET AL.] / PENTAPETALAE: root apical meristem closed; (cyanogenesis also via [iso]leucine, valine and phenylalanine pathways); flowers rather stereotyped: 5-merous, parts whorled, calyx and corolla distinct, stamens = 2x K, (often numerous, but then usually fasciculate and/or centrifugal), pollen tricolporate, G [5], G [3] also common, if G [2], carpels superposed, compitum +, placentation axile, style +, stigma not decurrent; endosperm nuclear; fruit dry, dehiscent, loculicidal [when a capsule]; whole genome triplication; RNase-based gametophytic incompatibility system present.
[DILLENIALES [SAXIFRAGALES [VITALES + ROSIDS s. str.]]]: stipules + [usually apparently inserted on the stem].
[SAXIFRAGALES [VITALES + ROSIDS]] / ROSANAE Takhtajan / SUPERROSIDAE: ??
[VITALES + ROSIDS] / ROSIDAE: anthers articulated [± dorsifixed, transition to filament narrow, connective thin].
ROSIDS: (mucilage cells with thickened inner periclinal walls and distinct cytoplasm); embryo long; chloroplast infA gene defunct, mitochondrial coxII.i3 intron 0.
ROSID I / FABIDAE / [ZYGOPHYLLALES [the COM clade + the nitrogen-fixing clade]]: endosperm scanty.
[the COM clade + the nitrogen-fixing clade]: ?
[CELASTRALES [OXALIDALES + MALPIGHIALES]] / the COM clade: seed exotegmic, cells fibrous.
[OXALIDALES + MALPIGHIALES]: ?
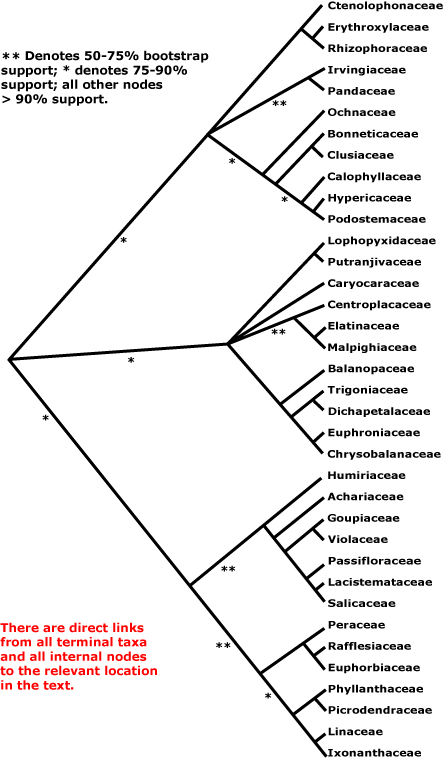
MALPIGHIALES Martius Main Tree, Synapomorphies.
Vessel element type?; (sieve tubes with non-dispersive protein bodies); lamina margin toothed [teeth with a single vein running into a congested ± deciduous apex]; stigma dry. - 39 families, 716 genera, 15935 species.
Includes Achariaceae, Balanopaceae, Bhesa, Bonnetiaceae, Calophyllaceae, Caryocaraceae, Centroplacaceae, Chrysobalanaceae, Clusiaceae, Ctenolophonaceae, Dichapetalaceae, Elatinaceae, Erythroxylaceae, Euphorbiaceae, Euphroniaceae, Goupiaceae, Humiriaceae, Hypericaceae, Irvingiaceae, Ixonanthaceae, Lacistemataceae, Linaceae, Lophopyxidaceae, Malpighiaceae, Malesherbiaceae (= Passifloraceae-Malesherboideae), Medusagynaceae (= Ochnaceae), Ochnaceae, Pandaceae, Passifloraceae, Peraceae, Phyllanthaceae, Picrodendraceae, Podostemaceae, Putranjivaceae, Quiinaceae (= Ochnaceae-Quiinoideae), Rafflesiaceae, Rhizophoraceae, Salicaceae, Trigoniaceae, Turneraceae (= Passifloraceae-Turneroideae), Violaceae.
Note: Possible apomorphies are in bold, (....) denotes a feature common in the clade, exact status uncertain, [....] includes explanatory material. Note that the particular node to which many characters, particularly the more cryptic ones, should be assigned is unclear. This is partly because homoplasy is very common, in addition, basic information for all too many characters is very incomplete, frequently coming from taxa well embedded in the clade of interest and so making the position of any putative apomorphy uncertain. Then there are the not-so-trivial issues of how character states are delimited and ancestral states are reconstructed (see above).
Evolution. Divergence & Distribution. Crown Malpighiales probably began radiating some time in the Cretaceous-late Aptian, some 101-114 m.y.a. ([119.4-]113.8[-110.7]/[105.9-]101.6[-101.1] m.y. - high and low estimates: Davis et al. 2005a); initial diversification seems to have been rapid. Other estimates are rather younger. The age of crown group Malpighiales was estimated as (93-)92, 90(-89) m.y. (two penalized likelihood dates), the stem group age being (107-)103(-99) and (95-)91(-87) m.y.; Bayesian relaxed clock estimates were slightly older, to 106 and 112 m.y. respectively (Wang et al. 2009). Wikström et al. (2001) suggested an age for the stem group of only (91-)88(-85) m.y., and for the crown group some (80-)77(-74) m.y. before present, nevertheless, stem groups of many families were evident before the beginning of the Caenozoic. Magallón and Castillo (2009: Celastrales sister to Malpighiales) estimated ages of ca 89.3 m.y. for both relaxed and constrained penalized likelihood datings for the divergence of crown Malpighiales, 98.6 and 98.9 m.y. (relaxed and constrained again) for the stem.
The order contains ca 7.8% eudicot diversity (Magallón et al. 1999) and show moderately high diversification rates (Magallón & Catillo 2009). Xi et al. (2012: see different methods of analysis) examined diversification rates throughout the clade, and found about eight clades in which the rates of diversification decelerated and about five in which they accelerated; these are mentioned individually below.
Ecology. Malpighiales are particularly important in tropical rainforests where they are a major component of the diversity of the understory; they account for up to some 28% of the species and 38% of the total stems there (Davis et al. 2005a), and members of Ericales are another major component of this vegetation. Note that this forest may not have developed until early in the Caenozoic (Burnham & Johnson 2004, see Caenozoic Diversification), somewhat at odds with the dates just mentioned.
Plant-Animal Interactions. Caterpillars of outgroups to Nymphalidae-Nymphalinae, -Melitaeini, etc., are quite common on Mapighiales (Nylin & Wahlberg 2008). The butterfly Cymothoë has hosts widely scattered in this order (Ackery 1988), although also found on Bignoniaceae (one species) and Rhamnaceae (sometimes another species). Phyllonorycter leaf-mining moths (Lepidoptera-Gracillariidae - Phyllocnistinae) seem to have diversified on this clade (and especially Fagales) some time in the region of 50.8-27.3 m.y.a., well after the Malpighiales diversified, and after the genus itself evolved, some 76.3-50.3 m.y.a. (Lopez-Vaamonde et al. 2006).
Chemistry, Morphology, etc. Paracytic stomata may characterise a sizeable clade in Malpighiales, and three-carpellate gynoecia are known friom many families. Articulation of the pedicels is another feature that may be common to the order. The intron in the atpF gene has been lost several times in Malpighiales, alone among angiosperms, however, this varies within Euphorbiaceae, Phyllanthaceae, and Picrodendraceae (Daniell et al. 2008).
See Endress and Matthews (2006b) for petal appendages, etc., in the order, while Matthews and Endress (2008) discuss other floral variation and Tokuoka and Tobe (2006) integrate testa anatomy and embryology with phylogeny. Tobe and Raven (2011: see also supplement) provide an invaluable summary of embryological data for the whole order although, as they note, many families are poorly known; they plot the distribution of some characters of embryology and seed on a phylogenetic tree, much of which is unresolved. Oginuma and Tobe (2010) provide the first chromosome counts for four families in the order. Furness (2011) looked at pollen development, focusing on the parietal-placentation clade; the massive amount of detail that she found is difficult to optimize on a tree, partly at least because of the still poor sampling.
Phylogeny. Although Malpighiales are now strongly supported as being monophyletic (e.g. Davis et al. 2005a; Wurdack & Davis 2009; Xi et al. 2012), relationships within them were initially poorly understood (e.g. Soltis et al. 2007a). Studies on some groups within Malpighiales suggested relationships within particular clades, e.g. Litt and Chase (1999), Schwarzbach and Ricklefs (2000), Chase et al. (2002), and Davis and Chase (2004), and these were in general agreement with relationships apparent in broader studies. Davis et al. (2005a) clarified some relationships in Malpighiales in a four-gene (all three compartments) analysis, in particular suggesting an association between the families with parietal placentation (and also Goupiaceae) and that Centroplacus should be recognised as a separate family (see also Korotkova et al. 2009 and Soltis et al. 2011 for relationships in Malpighiales). The isolated Ctenolophonaceae were linked with Erythroxylaceae and Rhizophoraceae, Bhesa with Centroplacaceae, etc. (Wurdack & Davis 2009). A [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Chrysobalanaceae + Euphroniaceae]]] clade had strong support, e.g. Davis et al. (2005a), Tokuoka and Tobe (2006) and Korotkova et al. (2009).
However, in 2011 there were still nine clades composed of two or more families along with seven separate families that together formed a very substantial basal polytomy in Malpighiales (Wurdack & Davis 2009; Xi et al. 2010; Soltis et al. 2011). However, relationships apparent in Xi et al. (2012) are much more resolved. The major analysis in this study used 78 protein-coding plastome genes and four ribosomal genes; families not included were Lophopyxidaceae, Malesherbiaceae and Rafflesiaceae (the last-named for obvious reasons). Other analyses included many more taxa but less complete sampling of genes (see Xi et al. 2012 for details). Malpighiales can now be divided into three main clades, the Salicaceae-Euphorbiaceae, Rhizophoraceae-Clusiaceae, and Malpighiaceae-Chrysobalanaceae clades, all with substantial molecular support (>80% ML bootstrap, 1.0 p.p.) and even with a modicum of morphological support. Although at the next level of the tree the second two of these clades have polytomies and the first an only weakly-supported dichotomy, overall the improvement of resolution in the tree is substantial (Xi et al. 2012), and the relationships suggested there are followed here.
Clade 1. [[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]] [[Peraceae [Rafflesiaceae + Euphorbiaceae]] [[Phyllanthaceae + Picrodendraceae] [Linaceae + Ixonanthaceae]]]].
Although support for the [Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]] clade is not strong (Xi et al. 2012), the clade excluding Humiriaceae (= the parietal clade) has very strong support. Goupiaceae are certainly to be included here, although their association with Violaceae is only weakly supported, as is the position of the combined clade (Xi et al. 2012). Major relationships in the rest of this clade have strong support (Xi et al. 2012).
Molecular evidence that the whole group of families with parietal placentation and (often) three carpels is monophyletic had initially not been compelling (e.g. see Savolainen et al. 2000a; Chase et al. 2002), although part of the rpS 16 gene is absent from Passifloraceae-Passifloroideae and -Turneroideae, Violaceae, and Salicaceae s. str. (and also Linaceae and Malpighiaceae, so really a feature of Malpighiales?: see Downie & Palmer 1992). Salicaceae were weakly associated with Passifloraceae, and in turn with Humiriaceae and Pandaceae, and Violaceae were weakly associated with Achariaceae (and Goupiaceae, Lacistemataceae and Ctenolophonaceae) in Chase et al (2002). Tokuoka and Tobe (2006) found a weakly-supported relationship between the Passifloraceae group and Violaceae (see also Soltis et al. 2007a), and strongly supported relationships between Lacistemataceae and Salicaceae. However, Davis et al. (2005a) found a moderately supported association of these taxa with parietal placentation (59% bootstrap, 1.00 posterior probability), and also Goupiaceae, with axile placentation, and a similar grouping is also evident in e.g. Wikström et al. (2001), Wurdack and Davis (2009), Korotkova et al. (2009: 83% jacknife, 1.00 pp, Goupiaceae not included) and Soltis et al. (2011: details of relationships unclear). Ixonanthes was rather surprisingly embedded in Achariaceae in the Bayesian analysis of Soltis et al. (2007a), but that was due to misidentification of the material, which was a species of Hydnocarpus (K. Wurdack, pers. comm.).
Indeed, classical morphological studies had been suggesting a grouping that included Salicaceae, Achariaceae and Violaceae, and sometimes also Passifloraceae and its segregates, Malesherbiaceae and Turneraceae, in part because of their common possession of parietal placentation, some sort of corona or scales in the flower, nectaries outside the stamens, etc. (e.g. Nandi et al. 1998). Furthermore, it was known that species of the old Flacourtiaceae had one of two kinds of seed coat: the exotegmen was either more or less fibrous - taxa with this kind of exotegmen are now mostly in Salicaceae - or massive and non-fibrous - taxa with this exotegmen are now in Achariaceae (Corner 1976). It was also commonly recognized that Salicaceae were simply an extreme morphology reflecting the wind pollination common in that family, and that they could be linked with some of the old Flacourtiaceae. Distinctive cyclopentenoid cyanogenic glucosides and/or cyclopentenyl fatty acids, including gynocardin, also occur sporadically here (Webber & Miller 2008). The inclusion of Goupiaceae in this clade is the only real surprise since it is morphologically rather distinct.
The other weakly supported clade in Clade 1 is [[Peraceae [Rafflesiaceae + Euphorbiaceae]] [[Phyllanthaceae + Picrodendraceae] [Linaceae + Ixonanthaceae]]]], the euphorbioids. This is an unexpected clade in that the fruits of a rather broadly delimited Euphorbiaceae (inc. both Phyllanthaceae and Putranjivaceae) are very distinctive, with the walls falling away leaving the persistent columella, and that was one of the main characters that I used to recognize herbarium material of the family. It is hardly surprising that Merino Sutter and Endress (1995) argued for a broad circumscription of the family. However, the clade [[Phyllanthaceae + Picrodendraceae] [Linaceae + Ixonanthaceae]] is strongly supported, as is the [Peraceae [Rafflesiaceae + Euphorbiaceae]] clade (Xi et al. 2012). The inclusion of Rafflesiaceae in Malpighiales follows the recent findings of Barkman et al. (2004, 2007), Davis and Wurdack (2004), and in particular Davis et al. (2006), who place it with strong support as sister to Euphorbiaceae s. str.
Clade 2. [[Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]], [Irvingiaceae + Pandaceae], [Ochnaceae [[Clusiaceae + Bonnetiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]]].
Weak support for an association of [Caryocaraceae [Linaceae + Irvingiaceae]] with [Rhizophoraceae + Erythroxylaceae] (Soltis et al. 2007a), has not been strengthened, although they have a number of features in common, such as a basally connate androecium, epitropous ovules with an endothelium, etc. (Matthews & Endress 2007). Although Ctenolophonaceae, etc., might also be associated, their floral similarities did not seem to be so great. However, Wurdack and Davis (2009) found support for the clade [Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]], but further relationships were unclear. In the recent study by Xi et al. (2012), the clade [Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]] (= rhizophoroids) had strong support, [Pandaceae + Irvingiaceae] (pandoids) had weak support.
The clade [Ochnaceae [[Clusiaceae + Bonnetiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]]] has only weak support (70% ML bootstrap, 0.81 p.p.: Xi et al. 2012), but its composition is consistent with morphology; Ochnaceae and Clusiaceae et al. (the latter = clusioids) also have a generally similar flavonoid spectrum (Hegnauer 1990). For relationships in the core Bonnetiaceae-Podostemaceae clade (= clusioids: Xi et al. 2012), Calophyllaceae being recognized, see Wurdack and Davis (2009; esp. Ruhfel et al. 2011).
Clade 3. [[Lophopyxidaceae + Putranjivaceae], Caryocaraceae, [Centroplacaceae [Elatinaceae + Malpighiaceae]], [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Chrysobalanaceae + Euphroniaceae]]]].
Although this clade has strong support in Xi et al. (2012), relationships within it are poorly understood. The [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Chrysobalanaceae + Euphroniaceae]]] and [Putranjivaceae + Lophopyxidaceae] clades (= chrysobalanoids and putranjivoids respectively) are well supported, but the [Centroplacaceae [Elatinaceae + Malpighiaceae]] clade (malpighioids) has poor support. The particular position of the distinctive Caryocaraceae is unclear, althougth there is little question that it belongs here (Xi et al. 2012). There was some support for Picrodendraceae as sister to the chrysobalanoids in Soltis et al. (2007a: as Pseudanthaceae, Phyllanthaceae not included), but this relationship has not been confirmed.
Classification. A.P.G. thought that it would be useful to adopt a narrow circumscription for families that used to be included in Flacourtiaceae and Euphorbiaceae s.l. Even if future work were to suggest reaggregation of genera that had been placed in those two families, the composition of the clades recognized would be different from those in nominally the same families by previous classifications. Indeed, the realignments caused by the break-up of the old Flacourtiaceae and integration with Salicaceae and Achariaceae correlate well with a number of morphological and anatomical characters (Wurdack & Davis 2009). Furthermore, these earlier decisions are compatible with the tree in Xi et al. (2012), for instance, to restore Euphorbiaceae to close to its old broad circumscription would require the inclusion of Linaceae, Ixonanthaceae and Rafflesiaceae.
Previous Relationships. The problems faced when working out the circumscription and relationships of the small family Ixonanthaceae, here sister to Linaceae (Clade 1, with strong support), are a microcosm of similar problems taxonomists had faced - yes, there are distinctive characters, but which reliably indicate relationships? Thus Robson and Airy Shaw (1962) drew attention to the "spiral convolution of the filaments and style" of Cyrillopsis (Ixonanthaceae: Clade 1) which, they thought, were points of similarity between this genus and Irvingiaceae (Clade 2). However, in Ixonanthaceae Allantospermum and some species of Ochthocosmus have flowers very similar to those of Cyrillopsis, with the thin calyx reflexed after anthesis (Phyllocosmus, Ixonanthes), while other species of Ochthocosmus have persistent, erect, almost scarious-looking sepals, as is common in Linaceae. Takhtajan (1997) included Allantospermum in Irvingiaceae; both have flowers with two carpels and seeds with copious endosperm, and the inflorescences of some Ixonanthaceae are very like those of Irvingiaceae. Bove (1997), on the other hand, suggested that Ixonanthaceae and Humiriaceae (Clade 1, but not immediately related) were sister taxa, both having ellagic acid, a "free" disc encircling the ovary, and an entire stigma. In the context of Linales (also including Linaceae, Hugoniaceae, Erythroxylaceae [also Clade 2, not immediately related to Erythroxylaceae]: see Cronquist 1981), Ixonanthaceae, Bove thought, were rather different in their free stamens, semi-inferior ovaries and pollen grains with supratectal spines. Recent work (2016) suggests that Allantospermum is properly to be included in Irvingiaceae, and overall, characters are consistent with this position.
Synonymy: Linineae Shipunov, Rhabdodendrineae Shipunov, Rhizophorineae Shipunov - Balanopales Engler, Chailletiales Link, Chrysobalanales Link, Elatinales Martius, Erythroxylales Link, Euphorbiales Berchtold & J. Presl, Flacourtiales Martius, Garciniales Martius, Homaliales Martius, Hypericales Berchtold & J. Presl, Irvingiales Doweld, Lacistematales Martius, Linales Berchtold & J. Presl, Malesherbiales Martius, Marathrales Dumortier, Medusagynales Reveal & Doweld, Ochnales Berchtold & J. Presl, Pandales Engler & Gilg, Passiflorales Berchtold & J. Presl, Phyllanthales Doweld, Podostemales Lindley, Rafflesiales Martius, Rhizophorales Berchtold & J. Presl, Salicales Lindley, Samydales Berchtold & J. Presl, Sauvagesiales Martius, Scyphostegiales Croizat, Stilaginales Martius, Turnerales Link, Violales Berchtold & J. Presl
[[Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]], [Irvingiaceae + Pandaceae], [Ochnaceae [[Bonnetiaceae + Clusiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]]]: cristarque cells +.
[Ctenolophonaceae [Erythroxylaceae + Rhizophoraceae]]: leaves opposite, stipules enclosing the terminal bud, interpetiolar; pedicels articulated; nectary to outside of A; A 10, of two lengths, antepetalous stamens longer than antesepalous, anthers ± basifixed, connate basally, (minute corona +); G postgenitally united, placentation apical, stigmas capitate/lobed, papillate; ovules 2/carpel, collateral, epitropous, outer integument thinner than the inner, nucellus laterally thin, disintegrates, endothelium +, placental obturator +; K persistent in fruit; seeds arillate, also exotestal; endosperm +.
Chemistry, Morphology, etc. See Matthews and Endress (2011) for details of the floral morphology of this clade. Tobe and Raven (2011) suggest that the three families here have a multiplicative inner integument, rather, at least sometimes it is very thick even at the time of fertilization.
Phylogeny. For relationships in this clade (= rhizophoroids), all well supported, see Xi et al. (2012).
CTENOLOPHONACEAE Exell & Mendonça Back to Malpighiales
Evergreen trees; ellagic acid?; vessel elements with scalariform perforation plates; calcium oxalate as single crystals; cuticle waxes 0; stomata anomo- or anisocytic; petiole bundle arcuate; hairs tufted/stellate; bud scales +; lamina margins entire; inflorescence terminal, ?thyrsoid; K quincuncial, basally connate, (with 1 trace), C protective in bud, contorted, caducous; A adnate to base of C; pollen 3-9 stephanocolporate; G [2], septae thin, style +, branches short; ovules with zig-zag micropyle, integuments lobed, outer integument ca 5 cells across, inner integument ca 11 cells across; fruit a [?kind] capsule, K swollen; seed single, persisting on columella; aril ± hairy [when dry!], exotestal cells large, subpalisade, the outer wall alone thickened, exotegmic cells laterally flattened, tracheidal; endosperm copious, cotyledons very large, folded; n = ?.
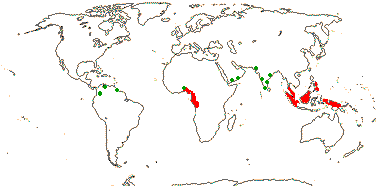
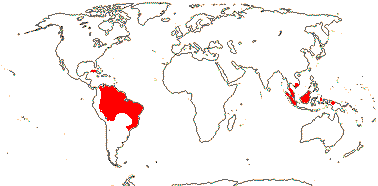
1[list]/3. W. Africa, Malesia (map: from van Hooren & Nooteboom 1988b; fossils [green] from Krutzsch 1989).
Evolution. Divergence & Distribution. Ctenolophonaceae may have diverged in the Cretaceous-Albian 111-100 m.y.a. ([109.6-]101.8[-96.6]/[97.1-]91.0[-88.1] m.y.: Davis et al. 2005a). The distinctive pollen is known as fossils from South America and India, the earliest records being from Africa in the Upper Cretaceous (Muller 1981; Krutzsch 1989). However, the diversification rate in this clade may have decreased (Xi et al. 2012).
Chemistry, Morphology, etc. Like Humiriaceae, there are "marginal" stomata on the disc and the anthers have a broad connective (Link 1992b); the wood anatomy is also similar. Takhtajan (1999) perhaps implies that there may be an endothelium, but embryology, etc., are largely unknown.
Some information is taken from van Hooren and Nooteboom (1988b) and Kubitzki (2013), general; for seed anatomy, see Huber (1991).
Previous Relationships. "Ctenolophon was almost universally recognized as belonging to the Linaceous alliance" (van Hooren & Noteboom 1988: p. 629).
[Erythroxylaceae + Rhizophoraceae]: tropane [hygroline] and pyrrolidine alkaloids, non-hydrolysable tannins +; sieve tube plastids with protein crystalloids; mucilage cells common; stomata paracytic; lamina vernation involute, colleters +; inflorescence cymose; K valvate, postgenitally united, C ± clawed, conduplicate-valvate, each C enclosing a stamen/stamens; median G adaxial, style somewhat impressed; (micropyle endostomal); fruit a septicidal capsule; exotestal cells enlarged, thick-walled, ± tanniniferous; endosperm starchy, embryo green.
Evolution. Divergence & Distribution. This clade may have diverged in the Cretaceous-Aptian around 114-110 m.y.a. ([119.3-]113.8[-110.2]/[105.7-]101.6[-102.1] m.y.: Davis et al. 2005a). The rate of diversification may have increased in Erythroxylaceae (Xi et al. 2012).
Chemistry, Morphology, etc. Although an unexpected family pair when contrasting Erythroxylum with mangrove Rhizophoraceae, when comparing Aneulophus (Erythroxylaceae) with non-mangrove Rhizophoraceae, the differences are then less obvious, and as noted above the two families are united by several synapomorphies. For floral development, see Matthews and Endress (2007).
Previous Relationships. Rhizophoraceae used to be placed in Myrtales (Cronquist 1981) or Myrtanae (Takhtajan 1997), largely because of their vestured pits and inferior ovary, but they are well supported as sister to Erythroxylaceae (e.g. Setoguchi et al. 1999; Schwarzbach & Ricklefs 2000; Chase et al. 2002; Korotkova et al. 2009).
ERYTHROXYLACEAE Kunth, nom. cons. Back to Malpighiales
Smallish trees and shrubs; mycorrhizae 0; ellagic acid 0; vessel elements with simple perforation plates; wood commonly with SiO2 grains; nodes with lateral bundles originating well before the central, forming cortical bundles; sclereids +; petiole bundle arcuate to annular with medullary and adaxial bundles; branching from previous flush; buds perulate; leaves usu. two-ranked (spiral), stipules usu. intrapetiolar; inflorescence often fasciculate; (pedicel not articulated - Aneulophus?), heterostyly common; (hypanthium + - Nectaropetalum); K connate basally, C protective in bud, with fringed bilobed ligule (0); nectary glands on outside of A tube; A obdiplostemonous, latrorse, (connective not thickened); pollen trinucleate; G [(2-)3(-4)], (adaxial only fertile), (short), (stylar canal +), branches well developed; ovule (1/carpel), outer integument 2-5 cells across, inner integument (ca 3?-)5-9 cells across, parietal tissue 2-4 cells across, nucellus below embryo sac extensive, endothelium +, hypostase 0, 2 vascular bundles in raphe; fruit often a 1-seeded drupe, A also persistent; (aril 0), tegmen multiplicative or not; (endosperm 0); n = 12, x = 7 (?6).
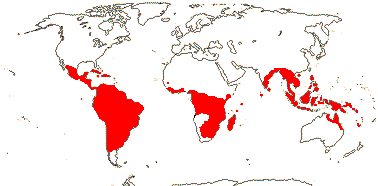
4[list]/240: Erythroxylum (230). Pantropical, esp. American (map: from van Steenis and van Balgooy 1966; Heywood 1978). [Photo - Flower, Fruit.]
Plant-Animal Interactions. Cocaine is sequestered by the larvae of Eloria noyesii, a lymanitrid moth.
Chemistry, Morphology, etc. The nodes were described as being unilacunar by Sinnott (1914), however, there are lateral traces although their gaps may be inconspicuous and the traces themselves may depart from the vascular cylinder well before the central trace (Rury 1982). Erythroxylum sometimes has milky exudate. Are the lamina teeth theoid? The leaves of Erythroxylum coca were described as being revolute by Cullen (1978); they are involute (e.g. Peyritsch 1878; Weberling et al. 1980; Rury 1982; Keller 1996). Matthews and Endress (2011) described the complexity of the postgenital fusion of the petals.
Aneulophus has seeds with a thick testa, thin tegmen, and aril, opposite leaves with colleters and inter/intrapetiolar stipules, and a septicidal capsule; from petal length, it appears that the flowers are monosymmetric.
See Bittrich (2013: general), for chemistry, see Hegnauer (1966, 1989) and Aniszewski (2007), for ovule and seed, see Rao (1968) and Boesewinkel and Geenen (1980).
Previous Relationships. This has been linked with Linaceae and Humiriaceae, and thence to Geraniales (Narayana & Rao 1978b), or the three families together are placed in Linales (Cronquist 1981).
Synonymy: Nectaropetalaceae Exell and Mendonça
RHIZOPHORACEAE Persoon, nom. cons. Back to Malpighiales
Trees; ellagic acid +; vessel elements with simple and/or scalariform perforation plates; true tracheids +; pits vestured; subepidermal laticifers in flower; cristarque cells 0; branching from current flush; inflorescence axis often evident; K (3-)4-5(-16), C small, often hairy, variously lobed, fringed, or with filiform appendages, aristate; A (= or) 2X C (more), anthers ± dorsifixed, (fasciculate; free); nectary inside A, on ovary or hypanthium; G [2-many], opposite sepals, when 2, lateral, septae often thin/disintegrating, style + (branched - Gynotroches), stigma also ± punctate, ?type; (micropyle also zig-zag), outer integument 3-6 cells across, inner integument 4-8(-20?) cells across, (endothelium 0), parietal tissue 1-3 cells across; megaspore mother cells several; (endotesta crystalliferous); endosperm with micropylar and chalazal haustoria + [?distribution], embryo (short), green; n = (13), 14, 16, 18, 21, x = 7; germination epigeal, cotyledonary node unilacunar.
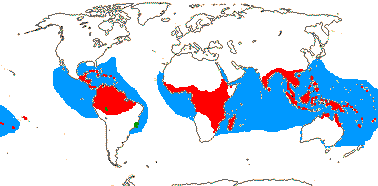
16[list]/149. Three groups below. Pantropical (map: from Ding Hou 1958; van Steenis 1963; Fl. Austral. 8. 1984; Tomlinson 1986; Juncosa & Tomlinson 1988a; Levin 1992). [Photo - Flower, Flower, Fruit.]
[Macariseae + Paradrypetes]: ?
1. Macariseae
Nodes 1:1 + split laterals [?all]; calcium oxalate crystals solitary; (leaves "alternate"), stipules valvate; (hypanthium +); K open; anthers latrorse; stigma not lobed; (seeds winged at micropylar end; arillate).
7/94: Cassipourea (62), Dactylopetalum (15). Tropical America and Africa, also peninsula India and Sri Lanka.
Synonymy: Cassipoureaceae J. Agardh, Legnotidaceae, nom. illeg., Macarisiaceae J. Agardh
2. Paradrypetes Kuhlmann
Raphides +; lamina with long, zig-zag intersecondary veins; plant dioecious; inflorescence epiphyllous, on petiole; flowers small; P +, 3-4; staminate flowers: pollen grains spiny; nectary 0; pistillate flowers: style 0; ovule with placental obturator; fruit a drupe; seed coat vascularized; endosperm starchy, abundant, cotyledons plicate, broad.
1/2. Brasil.
[Gynotrocheae + Rhizophoreae]: stilt roots present; rootlets without root hairs; leaves bijugate, stipules imbricate; hypanthium +; ovary ± inferior; obturator 0; fruit indehiscent; aril 0, testa vascularized.
3. Gynotrocheae
(Stilt roots 0); (petals entire); G often more than K, placentation axile [check]; ovules (to 8/carpel), tenuinucellate, outer integument 2-3 cells across, inner integument 2-4 cells across; megaspore mother cell 1); fruit a berry; exotesta mucilaginous, tanniniferous, other testal cells crystalliferous, tegmen 0, or fibrous to palisade, meso- and endotegmen persist; cotyledons short, or large, involute [Carallia, Pallacalyx].
4/30: Crossostylis (10). Indo-Malesia, Madagascar.
4. Rhizophoreae
Nodes 5:5, 7:7, + split-laterals; stomata cyclocytic; abaxial hypodermis +; sclerenchymatous sheath of midrib at most weakly developed; lamina vernation supervolute, margins entire; (C postgenitally united above base); (anthers locellate - Rhizophora); endothelium 0; fruit indehiscent, 1-seeded;seed coat undifferentiated, tegmen not persisting; (endosperm overflows from seed); (cotyledons convolute - Rhizophora, Bruguiera); seeds germinating on tree; cotyledonary node tri- or multilacunar.
4/17: Rhizophora (?9). Pantropical, centred on the eastern Indian Ocean, introduced into the central Pacific and Hawaii (see the blue area in the map above; Spalding et al. 2010).
Synonymy: Mangiaceae Rafinesque
Evolution. Divergence & Distribution. See below.
Ecology & Physiology. Mangrove taxa in Rhizophoraceae are derived within the family (e.g. Schwarzbach & Ricklefs 2000) and are most diverse in the Southeast Asia-Malesian area. Their seeds have little endosperm and are viviparous (aquatic/marine/mangrove plants in general quite commonly have large embryos, e.g. Feller et al. 2010), and in all genera except Bruguiera the endosperm overflows from the seed, pushing open the micropyle as it does so. After the seed falls from the tree it may float in the water, the hypocotyl straightening and establishment of the seeding being by the development of lateral roots (Juncosa & Tomlinson 1988b). Depending on the genus, there are either stilt roots, plank roots, or pneumatophores. Axillary buds soon die so the plants cannot regenerate when cut, or if the twigs are killed by frost, etc. (see Tomlinson 1986 for much useful information). Robert et al. (2009) discuss the hydraulic architecture of the wood of Rhizophora.
Rhizophoraceae-Rhizophoreae are a major component in the mangrove ecosystem, but overall only few species of flowering plants grow there, and apart from Rhizophoreae they are largely unrelated. A mere 34 species in nine genera and five families dominate the vegetation (half are Rhizophoraceae), while there are another 20 species in 11 genera and ten families (only one also including dominant species) that are quite common (Tomlinson 1986, estimates in Spalding et al. 2010 are 38 core species, 73 species of true mangroves). Mangroves can be divided into two groups, the much more speciose eastern group, from east Africa to the western Pacific, which includes ca 40 species, ca 14 of which are Rhizophoraceae, and the western group, from west Africa to the Americas, with only eight species, three of which are Rhizophoraceae; depending on how species limits are drawn, no common mangrove species is common to the two areas (Tomlinson 1986). Families include Primulaceae-Myrsinoideae (Aegiceras), Lythraceae (Sonneratia), Acanthaceae (Acanthus ilicifolius, Avicennia), Tetrameristaceae (Pelliceria) and Combretaceae (Lumnitzera). Of the dominant species, Nypa fruticans in particular forms monospecific stands growing along rivers to the upper limits of tidal influence. For the evolution of the mangrove ecosystem, which also involves diversification of clades of molluscs, etc. (Reid et al. 2008), see Ellison et al. (1999) and especially Plaziat et al. (2001 and references). A division of mangroves into two largely exclusive areas, the more diverse Indo-West Pacific and the less diverse Caribbean-West Atlantic areas, seems to have occurred by ca 20 m.y.a. (Plaziat et al. 2001).
Fossil and current distributions of these mangrove plants seem to have little to do with each other, and the history of individual mangrove species is complicated. By the Eocene, ca 50 m.y.a., many mangrove genera are known from the fossil record, and several, including Pelliciera, are known from both the Old and the New World (but see Martínez-Millán 2010). Pelliciera is now Central American, although growing in Europe in the past (Plaziat et al. 2001). Nypa (Arecaceae, q.v. for fossils), today found only in the Indo-Malesian area, appeared in the Upper Cretaceous ca 70 m.y.a. and by the early Palaeocene ca 55 m.y.a. was found in both the Old and New Worlds. Even though Rhizophora is known from the Caribbean in the late Eocene, the common ancestor of the existing populations there may have arrived in the New World some 40 m.y. later, only ca 11 m.y.a. (Graham 2006); there may be considerable genetic differentiation within Atlantic populations of mangrove species (Takayama et al. 2008a, b). Fossil hypocotyls identified as Ceriops and preserved with good anatomical detail are known from the Lower Eocene London Clay (Wilkinson 1981), although Collinson and van Bergen (2004) noted that the fossils did not show distinctive curvature of modern mangroves (Rhizophoreae) seedlings. At the other geographic extreme, Rhizophoreae are known from the Early Eocene 55-48.5 m.y.a. in western Tasmania, Australia (Pole 2007).
The mangrove ecosystem is very productive and has high carbon flux rates, and it also stores much carbon, especially below ground - at about 1,000 Mg C ha-1, storage is about three times as much as in temperate, boreal or tropical upland forests. Mangroves occupy 13.7-15.2 million hectares, and they may store 4-20 PgC globally (Bouillon et al. 2008; Donato et al. 2011 and references). Other estimates are that they bury 17.0-23.6 TgCy-1, their gross primary productivity is 2087 gCm2y-1, global primary productivity is 417 TgCy-1, but with a rather lower net ecosystem production (221 gCm2y-1> and globally 44 TgCy-1) because of a relatively high respiration rate, at least as compared with sea grasses (Duarte et al. 2005: area estimated at 20 million hectares). Mangrove peat can become very thick, the carbon in it being thousands of years old (McKee et al. 2007). See also Feller et al. (2010) for mangrove physiology, carbon flux, etc., and also Clade Asymmetries.
Pollination Biology. Pollen in Rhizophoreae is deposited on to the hairy petals, so there may be secondary pollen presentation, but pollination is basically explosive, the stamens being held in groups by the petals until the flower is tripped by the pollinator. These petals often have an arista or other appendages, and are shaped like a tiny bivalve mollusc (Endress & Matthews 2006b). The pollen grains are very small, and in Rhizophora in particular pollination may be by wind (Juncosa & Tomlinson 1988b).
Chemistry, Morphology, etc. Growth in a number of Rhizophoraceae may be continuous, although growth patterns in Macarisieae are unknown. Cork initation in the root is superficial in at least some taxa, perhaps just those with stilt roots (see von Guttenberg 1968 for Carallia). The leaf teeth are theoid.
There is considerable variation in merosity in the family, Carallia having K5 C5 A5 G5, and both carpel and stamen number vary (Matthews & Endress 2011). The stamens in polystemonous flowers arise from ring primordia (Ronse de Craene & Smets 1992b). Rhizophora has transversely arranged carpels (Eichler 1876). Variation in testal morphology in Gynotrocheae in particular is considerable, Gynotroches and Pellacalyx, with strongly exotegmic seeds, differing so much from Carallia, which lacks an exotegmen, that Corner (1976) preferred to segregate the former as Legnotidaceae - a comprehensive survey of seed anatomy in the family is desirable.
See also Juncosa and Tomlinson (1988) and Schwarzbackh (2013), general, Howard (1970) and Baranova and Jeffrey (2006), anatomy, Tobe and Raven (1987e, 1988b: seed coat anatomy), Endress and Matthews (2006b: petal morphology) and Carey (1934: embryology); for information on Paradrypetes, see Levin (1986, 1992) and Radcliffe Smith (2001 - as Euphorbiaceae).
Phylogeny. Schwarzbach and Ricklefs (2000) found strong phylogenetic structure in the family, with three major clades. At least some Macarisieae have stamens of two lengths and well-developed anther connectives (D. Kenfack, pers. comm.), probably plesiomorphic features. Crossostylis, with dehiscent fruits and arillate seeds, is embedded in Gynotrocheae, which otherwise have fleshy, indehiscent fruits and seeds without arils. Fleshy indehiscent fruits may have evolved in parallel within Gynotrocheae, or the arillate seed, etc., of Gynotroches is a reversal.
Rather surprisingly, molecular data also place Paradrypetes (ex Euphorbiaceae) here (e.g. Davis et al. 2005a), strongly supported as sister to Cassipourea (Wurdack & Davies 2008: only one species from each tribe included). Paradrypetes has a rather unexpected combination of characters and is highly apomorphic (see above).
Classification. Schwarzbach and Ricklefs (2000) suggested that three tribes be recognized for the three major clades that were apparent in their phylogeny of the family.
Previous Relationships. Rhizophoraceae have often been associated with Myrtales (e.g. Cronquist 1981; Takhtajan 1997), and they have sometimes included or been closely associated with (Takhtajan 1997) Anisophylleaceae, here in Cucurbitales.
[Irvingiaceae + Pandaceae]: leaves two-ranked, at least on plagiotropic axes; lamina vernation involute; flowers small; K connate basally; anthers basifixed; ovule 1/carpel, apical, pendulous, epitropous; fruit indehiscent; exotesta and endotegmen tanniniferous; n = 15, x = 7.
Evolution. Divergence & Distribution. The rate of diversification of this clade - it contains ca 25 species - may have decreased (Xi et al. 2012).
Phylogeny. Support for this clade (= pandoids) was rather weak in Xi et al. (2012: 64% ML bootstrap, 0.97 PP).
IRVINGIACEAE Exell & Mendonça Back to Malpighiales
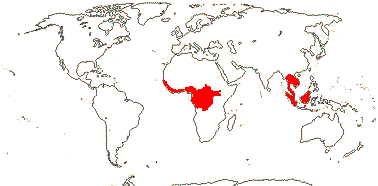
Trees; ellagic acid, myricetin +; vessel elements with simple perforation plates; nodes ?multilacunar; (sclereids +); petiole bundle annular and with associated bundles; mucilage cells in epidermis and ducts elsewhere in leaf/(0); stomata paracytic; branching from previous flush; leaves two-ranked, lamina margins entire, secondary veins strong, rather close and subparallel, tertiary veins also ± parallel and at right angles to the secondary veins, stipules very long (not), intrapetiolar and encircling terminal bud, deciduous; inflorescences racemose, branched, axillary or terminal; pedicels basally articulated; K cochlear; C protective in bud, cochlear or quincuncial, free, with 3 traces; A (9) 10, latrorse, filaments folded in bud; nectary massive, disk-like; G [(2) 5], G median (when 2) or opposite sepals, style single, stigma subcapitate-papillate, ?type; ovules sessile, attachment broad, micropyle bistomal, outer integument 2-3 cells across, inner integument 3-4 cells across, parietal tissue 3-4 cells across, (nucellar cap +, weak), epidermis at nucellar apex with radially elongated cells, placental obturator +, hypostase 0; embryo sac long; fruit a 1-seeded berry, 1- or 5-seeded drupe, or samara, or septicidal/part loculicidal capsule with columella [Allantospermum]; K deciduous or not; hilum long [Irvingia], outer (esp.) and inner integuments multiplicative, (testa with fascicles each of small bundles concentrically arranged in the antiraphal area, thick, inner part sclerotised - Irvingia), exotegmen fibrous/tracheidal, the rest ± collapsed; endosperm copious to 0; cotyledons large, cordate; x = 7, chromosomes 0.7-1.4 µm long; germination epigeal
4 [list]/12: Irvingia (7). Africa; South East Asia to W. Malesia (map: from Harris 1996). [Photo - Fruit]
Evolution. Divergence & Distribution. Stem-group Irvingiaceae may have diverged in the Cretaceous-Albian some 111-100 m.y.a. (Davis et al. 2005a).
Chemistry, Morphology, etc. Keller (1996) suggests that the leaves are involute in bud; this should be confirmed. Netolitzky (1926) is unclear about exactly where the fibrous layer is in the seeds of Desbordea and Klainedoxa, suggesting that the latter is exotestal, although Boesewinkel (1994) calls it exotegmic, which seems more likely.
See also Harris (1996: monograph) and Kubitzki (2013b) for general information, also Jadin (1901), Rojo (1968) and van Tieghem (1905a: stomata anomocytic?), all anatomy, Nooteboom (1967: chemistry), Weberling et al. (1980: stipules), Link (1992c: nectary), and Tobe and Raven (2011: stamen, ovules and seed); details of floral orientation are taken from Eckert (1966).
Previous Relationships. All over the place! Irvingia was included in Simaroubaceae-Sapindales by Cronquist (1981) and, kept separate as Irvingiaceae, placed in Rutales, in the same general area, by Takhtajan (1997). Irvingia is sister to Erythroxylum in a tree presented by Fernando et al. (1995), and the stipules of Irvingiaceae, Erthroxylaceae and Ixonanthaceae are similar (Weberling et al. 1980); Irvingiaceae are weakly associated with Putranjivaceae in Chase et al (2002a) and with Linaceae in Davis et al. (2005a). Prior to xii.2015, Allantospermum was in Ixonanthaceae...
PANDACEAE Engler & Gilg, nom. cons. Back to Malpighiales
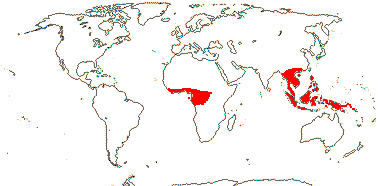
Trees to shrubs; cork?; vessels in radial multiples, vessel elements with scalariform (and simple - Galearia) perforation plates; rays 2-9 cells wide; sieve tubes with non-dispersive protein bodies; pericycle also with sclereids; druses and crystals +; petiole bundles D-shaped to (incurved-)arcuate; cuticle waxes 0; leaves spiral and reduced on orthotropic axes, lamina with a single vein running into the opaque persistent tooth apex, one stipule higher than the other on the stem; inflorescences various; plant dioecious; (K free), C valvate or imbricate, petals usu. thick, hooded to flat; nectary 0; staminate flowers: stamens = and opposite sepals, 10, or 15, in one or two series, basifixed, connective produced or not; pistillode +; carpellate flowers: staminodes 0; G [2-6], style 0, stigmas spreading, laciniate or entire; ovule (straight - Panda), micropyle endostomal, outer integument 3-5 cells across, inner integument 3-5 cells across, nucellar cap ca 6 cells across, obturator 0; fruit a 2-5-seeded drupe, stone surface often irregular; exotegmen tracheoidal, (many layered - Panda); endosperm ?development, +, cotyledons incumbent, thin and flat, oily; x = 7.
3[list]/15: Microdesmis (10). Tropics, Africa to New Guinea (map: in part from Léonard 1961; van Welzen 2011).
Evolution. Divergence & Distribution. Pandaceae seem to be a very old and isolated clade, dating back perhaps to the late Aptian (Cretaceous) 114-112 m.y.a. ([118.7-]113.8[-110.2]/[105.5-]101.6[-101.9] m.y.: Davis et al. 2005a).
Chemistry, Morphology, etc. Panda smells like onions. Microdesmis has punctate leaves. The plagiotropic branches have been confused with compound leaves, especially in the derived Galearia and Panda; the stipules may be asymmetrically placed, as in Panda. If the pedicels are articulated, they are articulated only at the very base.
For information - although there is little knowledge of embryology, etc. - mostly as Euphorbiaceae, see Forman (1966), Radcliffe-Smith (2001: genera), van Welzen (2011) and Kubitzki (2013b), all general, Vaughan and Rest (1969), Hegnauer (1969: chemistry), Stuppy (1996: seed anatomy), Nowicke et al. (1998: pollen), and Tokuoka and Tobe (2003: ovules and seeds).
Phylogeny. What is known about wood anatomy suggests that Galearia and Panda are close, while pollen suggests that Galearia and Microdesmis are close... (van Welzen 2011). The relationships [Microdesmis [Galearia + Panda]] are strongly supported by molcecular data (see Xi et al. 2012).
Classification. For a checklist and bibliography, see Govaerts et al. (2000, vol. 4).
Previous Relationships. Pandaceae are still often included in Euphorbiaceae, e.g. Govaerts et al. (2000) and Radcliffe-Smith (2001), but they differ from even the uniovulate taxa (Peraceae and Euphorbiaceae s. str.) in several respects, including their indehiscent fruits. Rays of Euphorbiaceae are only 1-5 cells wide (Hayden & Hayden 2000); Pandaceae lack obturators, while Euphorbiaceae have them - another difference. Dicoelia (Euphorbiaceae - Dicoelieae) and Galearia both have stamens in depressions in the petals. However, Dicoelia has a low, thin-walled testa, a massive exotegmen, and a moderately thickened mesotegmen (Stuppy 1996), and it is to be placed in Phyllanthaceae (Kathriarachchi et al. 2005). Centroplacus is also not included, although it is placed sister to Pandaceae, but without much support, by Wurdack et al. (2004); see Centroplacaceae here.
Engler had trouble with Panda, mistaking its plagiotropic branches for compound leaves, so he described it first as a species of Burseraceae, then as a species in Sapindaceae, and after he recognized his mistake, he was still unclear as to its relationships and placed it in a monotypic Pandales (Forman 1966).
[Ochnaceae [[Bonnetiaceae + Clusiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]]: biflavones +; C contorted; A many; nectary 0; (G [5+]); ovules lacking parietal tissue; fruit a septicidal or -fragal capsule; endosperm at most slight.
OCHNACEAE Candolle, nom. cons. Back to Malpighiales
Pits vestured; mucilage cells/canals +; branching from previous flush; lamina with secondary and tertiary venation well developed; pedicels articulated; A ³12 [some staminodial]; K persistent in fruit; x = 14 (?13), nuclear genome [1C] 90.013-)0.706(-39.5) pg.
33/561 - five groups below. Tropical, esp. Brazil.
1. Ochnoideae Burnett
Isoflavonoids +; stem with cortical (and medullary) bundles; (vessel elements with scalariform perforations); nodes also multilacunar; petiole bundle annular, (several, arcuate); stomata also paracytic; lamina with secondary veins strong and close, and/or with parallel tertiary veins, stipules fimbriate or not; flowers (3-)5(-10)-merous; K almost scarious; A (5-many), (development centrifugal), anthers (locellate), dehiscing by pores (not); G [(1-)5(-15)], opposite sepals, when 3 median member adaxial, placentation parietal, style long (short), stigma punctate or slightly lobed (± divided); ovules 1-many/carpel, endothelium +, micropyle endostomal; antipodals persistent; endotesta with small crystalliferous cells; endosperm slight, (embryo curved).
28[list]/495. Tropical, esp. Brasil (map: see Kanis 1971).
1A. Luxembergieae Horaninow
Androecium obliquely zygomorphic in bud, only adaxial A developing, filaments ± connate, anthers connate or not; pollen exine with small perforations; seeds winged; G [3]; n = ?
2/22. Venezuela and Brazil.
Synonymy: Luxemburgiaceae van Tieghem
[Ochneae + Sauvagesieae]: staminodes separate, forming a lobed disc or corolla-like tube, or 0; pollen with striate-rugulate exine.
1B. Ochneae Bartling
Vessel/parenchyma pitting not unilaterally compound; (petiole with inverted medullary bundle and subepidermal fibres); leaves two-ranked, (stipules semi-intrapetiolar - Ouratea); (inner edge of petal enveloping stamens in pairs); A ob/diplostemonous to many, development centripetal, filaments long to short, (anthers dehiscing by slits); (pollen 3-celled); G [2-]5[-15], (short gynophore +), style (gynobasic, receptacle expanded), hollow or not; ovule one/carpel, apotropous, integument single [= 2 fused, except sometimes at tip], 7-17 cells across, or micropyle bi- or endostomal, outer integument 3-4 cells across, inner integument 2-3 cells across [Ochna], hypostase + (/0?); embryo sac with antipodals enlarged; fruit indehiscent, usu. drupaceous, stamens persistent; testa with vascular bundles, lacking layer of small crystalliferous cells, fibrous exotegmen 0; endosperm 0; cotyledons massive, variously arranged, (unequal); n = 12-14.
9/390: Ouratea (inc. Gomphia: 200), Ochna (paraphyletic?: 85), Campylospermum (65). Tropical, especially Brazil. [Photo - Flower, Flower, Fruit.]
Synonymy: Gomphiaceae Schnizlein, Lophiraceae Loudon
1C. Sauvagesieae de Candolle
(Herbs); leaves spiral, (compound - Rhytidanthera), lamina vernation conduplicate-flat; (flowers monosymmetric, monosymmetry developing late, involving A and G); (K with outer members smaller than the rest); (pollen exine with small perforations); G [2, 3, 5], when 3, median member adaxial; ovules with outer integument ca 2 cells across, inner integument ?3-4 cells across; (seeds not winged); exotesta of large cells, ± detached, entotesta with crystalliferous cells; endosperm with aleurone; n = 18.
16/82: Sauvagesia (40). Pantropical, only 2 spp. in Africa, most South American.
Synonymy: Euthemidaceae van Tieghem, Sauvagesiaceae Dumortier, Wallaceaceae van Tieghem
[Medusagyne + Quiinoideae]: leaves opposite; styluli separate, ovary roof well developed, stigma expanded; ovules often 2/carpel.
2. Medusagynoideae Reveal
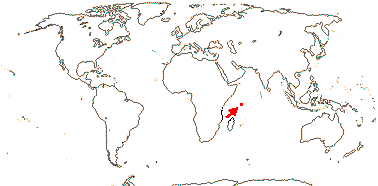
Plant tanniniferous; phloem stratified; true tracheids +; nodes 5:5 + 2 phloic bundles; cristarque cells 0; petiole bundles many, arcuate, variously oriented; hypodermal mucilage cells +; cuticle waxes 0; plant glabrous; buds perulate; leaves opposite, lamina venation very reticulate, stipules 0, colleters +; inflorescence terminal, ?cymose, plant andromonoecious; K basally connate; A spiral, from 5 trunk bundles, anthers basifixed; pollen porate; G [16-25], adnate to central axis, stigmas capitate, ?type; ovules 2-5/carpel, outer and inner integuments 3-4 cells across, funicles long; fruit a ribbed verrucose capsule, carpels pulling away acropetally and opening adaxially, columella persistent; seeds winged; exotesta slightly thickened; endosperm ?development, thin; n = ?
1[list]/1: Medusagyne oppositifolia. Seychelles, very rare.
Synonmy: Medusagynaceae Engler & Gilg, nom. cons.
3. Quiinoideae Luersson
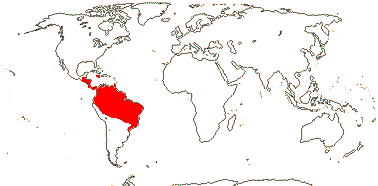
Trees (lianes); mycorrhizae 0; cork?; (vessel elements with scalariform perforation plates); true tracheids +; petiole bundle annular, often complex; stomata anisocytic; (leaves compound), lamina with strong, close secondary venation, tertiary venation paxillate, margins entire to deeply lobed, stipules pubescent, also interpetiolar, large, ± persistent; (plant androdioecious); K 4-5, pubescent, C 4-5(-8), usu. imbricate; A basally connate or not, (adnate to the base of the C), thecae distinct; pollen exine with small perforations; G 3 [2-13], stigmas obliquely expanded, type?; ovules 2/carpel, basal; fruit a berry (follicle), striate when dry, exocarp with lacunae; seeds 1-4, (usu. hairy), unwinged; coat ?; endosperm development?, 0, cotyledons massive; n = ?
4[list]/55: Quiina (25), Lacunaria (12). Tropical America (map: from Schneider et al. 2002). [Photo - Flower, Fruit.]
Synonymy: Quiinaceae Engler, nom. cons.
Evolution. Divergence & Distribution. This clade (= ochnoids: Xi et al. 2012) may have diverged in the Cretaceous-Albian (117-)111(-106()/(104-)99.6(-98.8) m.y.a.: Davis et al. 2005a). The restriction of Medusagyne to the Seychelles is noteworthy since the ocean crust separating India and the Seychelles started forming ca 63.4 m.y. old (Collier et al. 2008); the diversification rate in this clade may have slowed down (Xi et al. 2012).
Chemistry, Morphology, etc. Sauvagesia lacks vestured pits; two other genera in Ochnoideae are recorded as having them (Jansen et al. 2001). Godoya has stratified phloem. There are mucilage cells or mucilage channels, and the plants sometimes have watery juice.
For floral morphology in the whole group, see Matthews et al. (2011, esp. 2012, very useful). There is considerable variation in floral morphology in Ochnaceae-Ochnoideae. Sauvagesia has numerous linear staminodes, five petal-like staminodes opposite the petals, and five stamens opposite the sepals. The antesepalous primordia of Ochna (Ochnoideae-Ochneae) show centripetal androecial development (Pauzé & Sattler 1978), while the androecia of members of the other two tribes have centrifugal development (Amaral & Bittrich 1998). Zygomorphy is largely the result of the unequal later development of the androecium, but in Philacra and Luxembergia it is evident early in development (Amaral & Bittrich 1998). Although the anthers are often porose, there is still an endothecium (quite often absent in such situations), and this perhaps facilitates reversal from the porose condition (Amaral & Bittrich 2004).
There is also considerable variation in the ovule, etc., of Ochna, alternatively, some reports must be incorrect. Chikkannaiah and Mahalingappa (1974) suggest that there is no endothelium, but the nucellar epidermis seems to take over that function. Lophira has unequally accrescent sepals, two members forming wings (there are only two carpels, each with many ovules, and the testa is thin). Batygina et al. (1991, p. 222) show Sauvagesia erecta as having a much enlarged endotesta with thick walls.
For further information on Ochnoideae, see van Tieghem (1902: general, esp. embryo), Amaral (1991), and Amaral and Bittrich (2013), all general, Hegnauer (1966, 1989: chemistry), Dickison (1981: anatomy), and Narayana (1975) and Guèdès and Sastre (1981), both embryology.
In Medusagyne the upper ovules are ascending and epitropous, the lower ovules descending and apotropous (Batygina et al. 1991; Doweld 1998b). Additional information on Medusagyne is taken from Robinson et al. (1989) and Dickison and Kubitzki (2013), general, Dickison (1990a, 1990b: morphology and anatomy), and Fay et al. (1997a: relationships and morphology). For comparison of the fruit dehiscence of Medusagyne with that of some Ochnaceae, particularly some Sauvagesioideae, see Fay et al. (1997a); the anatomy of the fruits is similar to that of Caryocaraceae (Dickison 1990a).
The venation of the leaves of Quiinoideae is very distinctive, although not that dissimilar from that of other Ochnaceae, and it has been studied in detail by Foster (1952 and references). Veinlets ending free in the mesophyll can be few or even absent. The stomata are described as being paracytic by Schneider et al. (2002). For general information, see Kubitzki (2013b).
Medusagyne and Quiinoideae are largely unknown embryologically, etc.
Phylogeny. There is good molecular support for a monophyletic Ochnaceae s.l., e.g. Fay et al. (1997a), Nandi et al. (1998), Savolainen et al. (2000a), Chase et al. (2002) and Korotkova et al. (2009). Relationships between the three clades that make Ochnaceae up were less clear. However, Xi et al. (2012: as families) found moderate (75% ML bootstrap; 1.00 p.p.) support for a [Medusagynoideae + Quiinoideae] clade. Within Quiinoideae, Froesia is sister to the rest of Quiinoideae (Schneider et al. 2006, see also 2002 for a morphological phylogeny); it has separate carpels, follicles (apomorphies), and glabrous seeds (a plesiomorphy). Relationships among the other three genera are unclear.
Classification. Including Ochnaceae, Medusagynaceae and Quiinaceae in Ochnaceae s.l. was an optional arrangement in A.P.G. II, and they have much in common; Ochnaceae s.l. are recognized in A.P.G. III (2009).
Previous relationships. Diegodendraceae, included in Ochnaceae by Cronquist (1981), are placed in Malvales (see also Amaral 1991). Medusagyne is morphologically very distinctive. Comments on the species cover of Medusagyne at the Royal Botanical Gardens, Kew, ca 1985: "c.f. Actinidia. - Would be much better placed in Guttiferae or Hypericaceae - !!!!! - this plant allied to Myrtales. - Nonsense! - oh yes it is!". Hardly surprisingly, it was placed in a monotypic Medusagynales (Theanae) by Takhtajan (1997) and generally associated with Theales (e.g. Cronquist 1981); the latter was already such an heterogeneous group that the further inclusion of practically anything made little difference to its description... Van Tieghem (1902) thought that on balance Clusiaceae s.l. and Ochnaceae might be close, largely because of the polystemony of the former and also some of the latter.
Thanks. For discussion, and for comments on relationships within Ochnoideae, I am grateful to Maria Amaral and Volker Bittrich.
[[Bonnetiaceae + Clusiaceae] [Calophyllaceae [Hypericaceae + Podostemaceae]]]: flavones, flavonols, (ellagic acid), biphenyls, xanthones and dimeric xanthones, benzophenones, acylphloroglucinol derivatives, quinones +; vessel elements with simple perforation plates; schizogenous canals or cavities +; nodes 1:1; cristarque cells 0; stomata paracytic; leaves opposite, with colleters, lamina margins entire, stipules 0; inflorescence cymose; (A fasciculate, fascicles opposite C), anthers basifixed; G opposite sepals [check], or median member adaxial, stigma papillate; ovules many/carpel, micropyle exostomal; exotegmen with anticlinal walls sinuous, low, lignified; embryo ± fusiform.
Evolution. Divergence & Distribution. The stem of this clade seems to have diverged in the Cretaceous-Albian, 111-100 m.y.a., Clusiaceae (there sister to the rest of the clade) in turn diverging perhaps in the Cenomanian (104-)94(-92)/(95-)89(-87) m.y.a. (Davis et al. 2005a).
There are several potential morphological synapomorphies for the clade (= clusioids: Xi et al. 2012), and it is recovered even in morphological phylogenetic analyses (e.g. Luna & Ochoterena 2004 - Hypericaceae not included). Xanthones are uncommon elsewhere, being known from Gentianeae, some Moraceae, etc. The xanthones of Podostemaceae are similar to those both of Gentianaceae (in their -6-0-glucosides) and of Clusiaceae (in their isoprenyl substitutions). The morphology of Podostemaceae is so highly derived that finding synapomorphies with Hypericaceae is difficult.
Chemistry, Morphology, etc. For a summary of the chemistry of the group, see Crockett and Robson (2011); exactly where on the tree particular classes of secondary metabolites are to be placed will depend on more detailed sampling.
Given the likely phylogenetic relationships above, anatomical studies of Bonnetiaceae are needed to clarify the apparent absence - or near absence - of secretory tissues there. Takhtajan (1993) describes the pith as having secretory canals, as in Clusiaceae, but c.f. Baretta-Kuipers (1976). It is quite common for the calyx to be small relative to the corolla in bud, so the corolla has taken over the protective function, although in taxa like Calophyllum and Clusia the bud is at first completely enclosed by the sepals.
Phylogeny. Relationships within this clade were initially unclear (see also Soltis et al. 1999b; Gustaffson et al. 2002; Davis et al. 2005b), although Wurdack and Davis (2009) and particularly Ruhfel et al. (2011, see also Xi et al. 2012) have recently confirmed the paraphyly of the old Clusiaceae, necessitating the separation of Calophyllaceae (= Clusiaceae-Kielmeyeroideae of versions 8 and before). Support for all relationships is strong, although that for the [Bonnetiaceae + Clusiaceae] clade is the weakest (Xi et al. 2012). The branch leading to Podostemaceae is rather long. The xanthones of Podostemaceae are similar to those both of Gentianaceae (in the -6-0-glucosides) and of Clusiaceae (in the isoprenyl substitutions).
Previous Relationships. Morphological data in particular (most of the features above) initially seemed to suggest a grouping of Elatinaceae + Bonneticaceae + Clusiaceae/Hypericaceae (e.g. see versions of this site prior to version 5). This was not a monophyletic group in Savolainen et al. (2000a), indeed, Ploiarium is there placed in Malvales (but see Wurdack & Davis 2009), although testa anatomy, etc., are strongly against such a position. Analyses in Chase et al. (2002) weakly link Elatinaceae and Bonnetiaceae + Clusiaceae + Podostemaceae, but the evidence now suggests that Elatinaceae are sister to Malpighiaceae (Davis & Chase 2004; Davis et al. 2005a; Tokuoka & Tobe 2006; Wurdack & Davis 2009), and some morphological data support this.
Classification. The old Clusiaceae were strongly paraphyletic, so their continued recognition would entail the inclusion of Bonnetiaceae and Podostemaceae, and also Hypericaceae. For the "price" of recognizing Calophyllaceae, we have five coherent and moderately easy recognizable clades.
[Bonnetiaceae + Clusiaceae]: ?
BONNETIACEAE Nakai Back to Malpighiales
Evergreen shrubs; biphenyls, biflavones 0 (nodes 3:3); schizogenous cavities 0?; mucilage cells common; plant glabrous; leaves spiral, lamina vernation supervolute, margins minutely toothed by setae, petiole short; C protective in bud; G [3-5], style long, hollow, or style branches ± separate, stigma papillate; cotyledons usu. small (-50% the embryo); n = ca 150 [Bonnetia cubensis].
3[list]/35: Bonnetia (30). Cambodia, Malesia (mostly Western), Cuba, South America. [Photo - Flower] [Photo - Flower]
Chemistry, Morphology, etc. Bonnettia s.l. has trilacunar nodes, a mucilaginous epidermis, a foliar endodermis, and foliar sclereids; Archytaea and Ploiarium have unilacunar nodes and lack the distinctive epidermis, foliar endodermis and sclereids (Dickison & Weitzman 1996). The absence of any secretory system should be confirmed. Keller (1996) described the leaf vernation as being involute.
Bonnetia also often has 3-trace petals while the bracetoles sometimes have only a single trace (Dickison & Weitzman 1998). For Archytaea, Wawra de Fernsee (1886) shows a floral diagram in which both the five carpels and the stamen fascicles are drawn opposite the calyx. Ovule morphology in Bonnetiaceae appears to be unknown.
For chemistry, see Hegnauer (1969, as Theaceae), and for a general account, see Weitzman et al. (2006).
Previous Relationships. Savolainen et al. (2000b) found that Ploiarium was placed within Thymelaeaceae. Morphologically and anatomically this position would seem rather unlikely - probably the leaf was from Gonostylus...
CLUSIACEAE Lindley, nom. cons.//GUTTIFERAE Jussieu, nom. cons., nom. alt. Back to Malpighiales
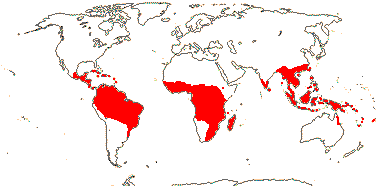
Evergreen trees or shrubs (lianes), (epiphytes); isoflavones, diterpenes; (vessel elements with scalariform perforations); lamina vernation often flat (conduplicate), margins entire, exudate in ± branched canals, (paired "stipular glands" on the stem); plant dioecious (flowers perfect - Symphonia, etc.); flowers (3-)4-5-merous, K and C decussate (contorted), (0-)4-5(-8); A (5-15), connate or not, (anthers extrorse), (with small glands), filaments as thick as anthers (not); G [2-5(-12)], often opposite petals, (placentation apical, basal, parietal), (ovary with a roof - Clusieae), style short, single or styluli widely separate, stigmas much expanded to punctate and porose, wet; ovules (1-few/carpel), (micropyle bistomal), outer integument 2-8 cells across (18-22 cells, Pentadesma), inner integument 2-7 cells across (10-15 cells, Pentadesma), 1(-2) lateral nucellar cells [Garcinia]; (fruit a berry); seeds few-many, (arillate, aril vascularized or not); (testa multiplicative), (exotegmen 0, then testa usu. much developed); embryo green or white, hypocotyl much enlarged, cotyledons minute; n = 7 (?6), nuclear genome [1C] ?; germination crypto(phanero-)cotylar, hypogeal (epigeal).
14 [list]/595: Garcinia (240: esp. Old World), Clusia (300-400: entirely American), Chrysochlamys (55). Throughout the tropics (map: from Gustaffson et al. 2007; Fang et al. 2011; various African floras...). [Photo - Staminate flower, Fruit.]
Evolution. Divergence & Distribution. For discussion on an interesting and well-preserved late-Cretaceous fossil ca 90 m.y. in age that is possibly assignable to Clusiaceae, see Crepet & Nixon (1998). The seeds are described as being arillate, but the morphology of the aril is unlike that of extant Clusiaceae (it is adjacent to the seed, rather than surrounding it), and it may even be an aborted seed.
Pollination Biology. Variation in the androecium and gynoecium in Clusia and Garcinia in particular is extreme. In Clusia, resins (polyisoprenylated benzophenones, mixed with fatty acids) are quite commonly a floral reward, and floral resin production may have evolved four times (as well as in Clusiella [Calophyllaceae]: Gustaffson & Bittrich 2002). Euglossine and especially stingless Trigona (meliponine) bees have been observed at Clusia flowers, interestingly, species of Clusia at higher altitudes, where bees are less common, produce nectar as a floral reward (Armbruster 1984). In general, resins are an uncommon floral reward (but see also Dalechampia [Euphorbiaceae] and Maxillaria [Orchidaceae]). Fragrant oils are produced from the filaments of the flowers of Tovomita; these attract male euglossine bees, and the composition of the fragrances in three different species growing in the Ducke Nature Reserve was quite different (Noguiera et al. 1998). In Symphonia pollen is caught in a droplet that exudes through the pore at the tip of the stylar branches, and is sucked back into the pore (Bittrich & Amaral 1996); the same unusual mechanism probably occurs in other Symphonieae all of which have similar stigmas.
Ecology & Physiology. Garcinia is one of the five most diverse genera in West Malesian l.t.r.f. (Davies et al. 2005). Clusia is a very diverse genus and includes epiphytes and stranglers many of which are more or less leaf succulents, and a number of species grow at elevations up to 3500 m in altitude (Gustafsson et al. 2007). For the general ecology of Clusia, see papers in Lüttge (2007). Crassulacean acid metabolism has been reported from at least some of these epiphytes (Holtum et al. 2004), and its development may be promoted by phosphorus deficiency; whether or not the plant is mycorrhizal also affects the plant's phosphorus and carbon metabolism (Maiquetía et al. 2009).
Chemistry, Morphology, etc. Roots of Clusia, at least, have superficial phellogen, as is fairly common in epiphytic taxa in general. For the morphology of the variable androecium in Garcinia and its immediate relatives, see Leins and Erbar (1991), Sweeney (2008, 2010) and Leal et al. (2012); the nectary is unlikely to be staminodial. Leal et al. (2012) and Tobe and Raven (2011) describe the ovules as being bitegmic (c.f. Corner 1976).
For chemistry, see Hegnauer (1966, 1989) and for general information, see Stevens (2006c).
Synonymy: Cambogiaceae Horaninow, Garciniaceae Bartling
[Calophyllaceae [Hypericaceae + Podostemaceae]]: exudate in pale [yellowish] gland dots or unbranched canals.
CALOPHYLLACEAE J. Agardh Back to Malpighiales
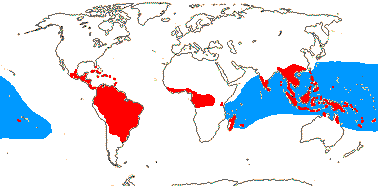
Evergreen trees or shrubs; (vessel elements with scalariform perforations); (leaves spiral, two-ranked), lamina vernation often flat (conduplicate; supervolute - Kielmeyera), (paired "stipular glands" on the stem; colleters 0); flowers 4-5-merous, C (contorted), (protective in bud), (0-)4-5(-8); A not obviously fasciculate, (connate), anthers often with complex or simple glands; G (?1) [2-5], (placentation apical, basal), style usually long, stigmas much expanded to punctate, wet; ovules (1-few/carpel), outer integument 20-30+ cells across [Calophyllum], inner integument 2-3 cells across; (fruit a berry or drupe); seeds 1-many, (testa multiplicative, Calophyllum), (exotegmen 0); embryo green or white, cotyledons moderate sized to huge [Calophyllum, Mesua, etc.]; n = 8 (?18, ?9); germination phanerocotylar, epigeal/cryptocotylar, hypogeal.
13[list]/460: Calophyllum (190), Kayea (70), Mammea (70), Kielmeyera (50). Throughout the tropics (map: in part see Stevens 1980 - blue is Calophyllum inophyllum). [Photo - Flower, Flower.]
Chemistry, Morphology, etc. Marila asymmetralis, alone in the whole family group, has obliquely monosymmetric flowers. The glands on anthers of genera like Caraipa are large, paired and crateriform, perhaps because the contents have been removed, while in other genera like Kayea they are small and rounded. Alternate-leaved genera form a clade; these genera also have capsular fruits, often with winged seeds, and their embryos have cotyledons with cordate bases. Although Calophyllum has opposite leaves, a few species have seedlings with alternate leaves.
For chemistry, see Hegnauer (1966, 1989, as Guttiferae), for general information, see Stevens (2006c, as Clusiaceae), and for the distinctive foliar fibres of many species of Mammea, see Dunthorn (2009).
Phylogeny. Clusiella is to be included in this clade (Gustaffson et al. 2002). Its seeds and vegetative anatomy (including the deep-seated phellogen of the root) are consistent with this position, although the flowers are a little odd, since they do indeed look like those of Clusia.
Previous Relationships. Many Theaceae also have spiral leaves, capsular fruits, winged seeds, and flowers with many stamens, and alternate-leaved Calophyllaceae seemed superficially to be similar and so used to be placed in that family. Another example of "intermediates" between groups that turn out not to be closely related at all (c.f. Baretta-Kuipers 1976).
[Hypericaceae + Podostemaceae]: ?
Evolution. Divergence & Distribution. Divergence between Podostemaceae and Hypericaceae may have occurred in the Campanian, (82-)76(-69)/(74-)72(-66) m.y.a. (Davis et al. 2005a).
HYPERICACEAE Jussieu, nom. cons. Back to Malpighiales
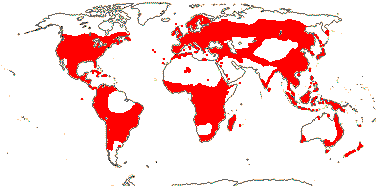
Small trees to shrubs or annual to perennial herbs; lignans, flavones, flavonols, (ellagic acid) +; cork pericyclic; polyderm widespread; dark glands also present; flowers 4-5-merous, C (imbricate), (protective in bud); A (5-15), often fasciculate, centrifugal, anthers dorsifixed, often with simple glands; G [3-5], (placentation parietal), style single, or styles separate, stigma papillate (not), dry; ovules (with bistomal micropyle), outer integument ca 2 cells across, inner integument 2-6 cells across; (fruit a berry or drupe); seeds (5-)many, (exotegmen 0); endosperm with chalazal cyst [Hypericum], embryo green or white, cotyledons moderate-sized (to 80% of the embryo); n = 6-12, etc., x = 9 (?10, ?7), nuclear genome (0.05-)0.398(-3.186) pg.
9[list]/560: Hypericum (370), Vismia (55), Harungana (50). World-wide (map: from Hultén & Fries 1986; Meusel et al. 1978 - still incomplete for Africa). [Photo - Flower]
Evolution. Divergence & Distribution. For the fossil record of the family - rather skimpy - see Meseguer and Sanmartin (2012); seeds of Hypericum antiquum don't seem to be notably hypericaceous.
Plant-Animal Interactions. Production of hyperforins (acylphloroglucinols, present in pale grands) and hypericins (phototoxic anthraquinones, present in dark glands) increased in Hypericum perforatum when eaten by generalist herbivores, but not by specialists (Sirvent et al. 2003).
Seed Dispersal. Vismia is an important food source for phyllostomid bats such as Carollia (Lobova et al. 2009).
Bacterial/Fungal Associations. The naptha-dianthrone hypericin may be synthesized by an endophytic fungus close to Chaetomium (Kusari et al. 2008).
Chemistry, Morphology, etc. For ovules, see Guignard (1893) and Nagaraja Rao (1957), for chemistry, see Hegnauer (1966, 1989, as Guttiferae), for androecial development, etc., see Leins and Erbar (1991, 2010), Leins (2000) and Ronse de Craene and Smets (1991e: Harungana), for secretory structures in vegetative parts of Hypericum, see Lotocka and Osinska (2010). For general information, see Stevens (2006c).
Phylogeny. Nürk and Blattner (2010) discuss relationships and evolution in Hypericum evident in an analysis of morphological characters; the groupings have little support.
Classification. Generic limits need attention, with those of Hypericum to be expanded and those of Harungana in particular probably to be restricted (Ruhfel et al. 2009, esp. 2011; c.f. in part Stevens 2006c, Nürk & Blattner 2010). For a monograph of Hypericum, see Robson (2010) and references.
Synonymy: Ascyraceae Plenck
PODOSTEMACEAE Kunth, nom. cons. Back to Malpighiales
Annual (perennial) herbs of fast-flowing water; quinones 0, benzophenones, etc.?; plant ± thalloid, stem root and leaf often not distinguishable, plant attached to substrate by haptera, basic construction sympodial; primary root 0, other roots (dorsiventrally flattened), photosynthetic, exogenous or endogenous, (apical meristem 0); shoots endogenous, [also at least sometimes flowers], branching extra-axillary; cork?; vessels usu 0; ?resin cells +; epidermal SiO2 bodies common; cuticle waxes 0; when leaves present spiral, opposite, 2- or 3-ranked, leaf base broad or not, stipules petiolar or 0; (flowers solitary: monosymmetric); P +, in single whorl; A 1-many (2 whorls, inner extrorse), filaments often basally connate (connective prolonged); pollen microechinate, infratectum granular; when G equals P, opposite to it, style + (0), stigma linear; ovules (2-few/carpel), (embryo sac protruding), outer integument 2(-4) cells across, inner integument ca 2 cells across, nucellus plasmodial before fertilization; embryo sac monosporic, from the subchalazal spore, tetranucleate [Apinagia type], polar nuclei degenerate; capsule ribbed, about the same size as the ovary, pedicels elongating; exotesta thick-walled, often mucilaginous, (exo- and) endotegmen ± lignified; no double fertilization, embryonic suspensor haustorial; cotyledons large; n = 10, x = 10 (?9, ?20).
48[list]/270 - three subfamilies below. Usually tropical, esp. America.
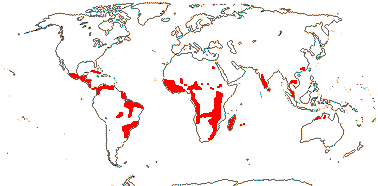
1. Tristichoideae Engler
Xanthones?; (root 0), (root cap 0); (stem flattened); stomata?; P 3, connate; A (1-)3, anthers sagittate; pollen pantoporate; G [3]; integuments develop simultaneously; hypocotyl 0.
3/4-10. India and Southeast Asia to Australia, Tristicha trifaria in Africa and America (map: from van Royen 1953; Cusset & Cusset 1988a; Kito & Kato 2004; Kato 2009).
Synonymy: Philocrenaceae Bongard, Tristichaceae J. C. Willis
[Weddellinoideae + Podostemoideae]: G [2], with apical septum only.

2. Weddellinoideae Engler
Plant with scales; flowers single, terminal; P (4) 5 (6), 1-veined; A 5-25, anthers X-shaped; pollen ?development, smooth, ?infratectum?; stigma single, globose; integuments develop simultaneously; capsule not ribbed; tegmen [?layer] thick walled; hypocotyl +.
1/1: Weddellina squamulosa. N. South America (map: from van Royen 1953).
3. Podostemoideae Engler
Shoot apical meristem 0; apical meristems of root on the underside of the thallus, roots foliose or ribbon-like; ("laticiferous" tubes +);
stomata 0, "epidermal" cells with dimorphic chloroplasts; "leaves" ± endogenous, often
2-ranked, ensiform, but bifacial, (digitate), (some leaves dithecous [double-sheathed, one sheath on both sides]), (leaves with 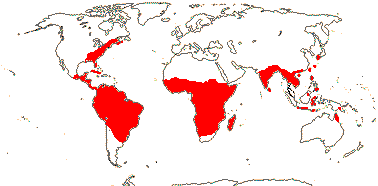 axillary branches, not dithecous - Thelethylax), (stipulate); flowers or groups of flowers enveloped by a
non-vascularized spathella, (spathella of non-terminal flower open - Diamantina), (flowers monosymmetric), (flowers inverted in bud - some African taxa); P 2-25, often 2-3 on one side, lobes narrow, sometimes replaced by stamens; A 1-3(-many), often sagittate, (extrorse);
(microsporogenesis successive [tetrads tetragonal]), pollen often in dyads (tetrads), (a)calymmate, 3(-5)-colpate; G also [3(-7)], (unilocular), gynophore + (0), style short, branches long, (2 styluli - Diamantina); ovule with outer integument that develops first, (apex of nucellus exposed); (embryo sac bisporic [Polypleurum and Podostemon types]);
embryo lacking radicle, (plumule 0; cotyledon 1); (n = 14). Floral Diagram.
axillary branches, not dithecous - Thelethylax), (stipulate); flowers or groups of flowers enveloped by a
non-vascularized spathella, (spathella of non-terminal flower open - Diamantina), (flowers monosymmetric), (flowers inverted in bud - some African taxa); P 2-25, often 2-3 on one side, lobes narrow, sometimes replaced by stamens; A 1-3(-many), often sagittate, (extrorse);
(microsporogenesis successive [tetrads tetragonal]), pollen often in dyads (tetrads), (a)calymmate, 3(-5)-colpate; G also [3(-7)], (unilocular), gynophore + (0), style short, branches long, (2 styluli - Diamantina); ovule with outer integument that develops first, (apex of nucellus exposed); (embryo sac bisporic [Polypleurum and Podostemon types]);
embryo lacking radicle, (plumule 0; cotyledon 1); (n = 14). Floral Diagram.
45/260: Apinagia (50: perhaps paraphyletic, see Philbrick et al. 2001). Pantropical (map: from van Royen 1951; van Steenis 1972; Kato 2009). [Photo - Marathrum Flower]
Synonymy: Marathraceae Dumortier
Evolution. Bacterial/Fungal Associations. Although there have been suggestions that Podostemaceae are attached to rocks by means of a special glue that they produce, it is more likely that it is materials in a biofilm produced by associated cyanobacteria that attach the plant to the substrate. There are hooked hairs on the lower side of the thallus that stick to the cyanobacterial filaments and associated biofilm. Indeed, these cyanobacteria may even produce nitrogen used by the plant; Podostemaceae usually grow in oligotrophic rivers flowing over gneiss or granite, being absent in rivers over limestone (Jäger-Zürn & Grubert 2000).
Pollination Biology & Seed Dispersal. Some Podostemaceae self pollinate, the pollen tubes growing through the tissue of the flower to the ovules (Sehgal et al. 2009).
There is considerable variation (and a corresponding amount of controversy) over the development of the embryo sac in particular, perhaps especially in Weddellina (see e.g. Battaglia 1971; Arekal & Nagendran 1975, 1977a, b; Nagendran et al. 1976; Murguía-Sánchez et al. 2002; Sehgal et al. 2011a). Is it mono- or bisporic, and does it have 3, 4 or 5 cells at maturity? However, there is general consensus that there is no double fertilization, the single polar nucleus degenerating (Sehgal et al. 2011b). During development of the embryo, the nucellus becomes amoeboid and is involved in the nutrition of the embryo; there are different pathways by which the nucellus becomes amoeboid (see Jäger-Zürn 1997).
For germination and establishment, see Grubert (1970, 1976); mucilage from the testa firmly attaches the seed to a rock. Although the embryo usually lacks a plumule and radicle, these were reported for Malaccotristicha sp. (Kita & Kato 2005), and the seedling of Zeylanidium olivaceum has a hypocotyl; more information is needed on embryo morphology (but see Koi & Kato 2010).
Vegetative Variation. Interpretations of the plant body of Podostemaceae, the "thallus", vary, and saltational evolution has been invoked to explain how very different and distinctive morphologies in related taxa have evolved (Koi & Kato 2010). It has been suggested that the thallus is a highly modified but ultimately fairly conventional plant body (Jäger-Zürn 2005), or that it cannot be compared with any other plant structure. Since Podostemaceae are sister to Hypericaceae, detailed studies of the latter may provide clues for the evolution of the growth of the former, particularly building on the developmental studies of Katayama et al. (2010).
Podostemaceae with ribbon-like roots have opposite branching, those with a crustose or foliose growth form have endogenous shoots born singly on the upper surface. The evolution of the remarkable flattened roots of some Podostemoideae and Tristichoideae, which lack caps and have meristematic regions on both sides of the root, from the more ordinary-looking roots found in Weddelinoideae and some other Tristichoideae has been carefully documented by Koi et al. (2006). The exogenous or superficial origin of roots of some Podostemoideae is very unusual, since roots are normally endogenous, being initiated inside the pericycle; Cladopus has both exogenous and endogenous lateral roots (Rutishauser & Pfeifer 2002). Whatever their origin, roots often have root caps.
The apex of the stem has a tunica-corpus construction. There is some controversy over whether normal axillary branching occurs or not (e.g. Rutishauser et al. 2005; Jäger-Zürn 2009a). Some taxa have shoots arising endogenously in the cortex (e.g. Moline et al. 2007). Dithecous leaves usually terminate growth of the axis that bears them; the leaf bases have two concave sheaths facing in opposite directions and in the axils of each a flower or branch bud arises (Rutishauser et al. 2003); for the optimisation of these dithecous leaves on a phylogeny of Podostemoideae, see Moline et al. (2007: note the adaxial position of the prophyll of the axillary dithecal vegetative shoot illustrated). Jäger-Zürn (2009b) also depicts the dithecous leaf as being adaxial on the axillary shoot that bears it.
Katoyama et al. (2010) found from gene expression patterns in two Podostemoideae that it is almost as if a determinate "leaf" caps the indeterminate stem, new leaves/branches developing endogenously from the base of the "leaf". Since separation of the young "leaves" is by cell death, they will lack an epidermis, hence, perhaps, the absence of stomata in the subfamily - although they might be found on the flowers, in which there is more normal development (e.g. Katayama et al. 2008). The "epidermal" cells have dimorphic chloroplasts; smaller chloroplasts are found against the outer periclinal walls and much larger chloroplasts against the inner periclinal walls (Fujinami et al. 2011).
Chemistry, Morphology, etc. Grubert (1976) noted distinctive contents in cells of young plants of several Podostemaceae. For SiO2 bodies, see Machado da Costa (2011).
The nature of the spathella is unclear. Unlike bracts in Podostemoideae examined, its development is that of a "typical organ of leaf homology" (Katayama et al. 2010, see also Katayama et al. 2008), although Eckardt and Baum (2010) suggest more specifically that it is calycine. However, if there is more than one flower per spathella, then it cannot be calycine. Some species of Dalzellia (Tristichoideae) have a cupule at the base of the pedicel that is formed by leafy shoot axes (Mathew et al. 2001).
Tristicha - A 1, adaxial, median carpel abaxial? (Schnell 1998). The position of the stamens may suggest an obliquely monosymmetric flower (Cusset & Cusset 1988b), while Razi (1955; see also Endress & Matthews 2006a) described the flowers of Zeylanidium olivaceum, which have a spathella, two stamens and two carpels, as being monosymmetric. There is sometimes an apical meristem in the vegetative body of Podostemoideae and the spathella may be produced by the connation of two foliar structures (Jäger-Zürn 2005b). The outer integument develops early and the nucellus protrudes beyond the inner integument. The amoeboid nucellus has been described as a pseudo-embryo sac. Tobe and Raven (2011) describe the tegmen as being unspecialised.
Much information is taken from Rutishauser (1997); see also Hegnauer (1969, 1990), Contreras et al. (1993) and Kato et al. (2005) - all these chemistry, Graham and Wood (1975), Barlow (1986: roots), Cusset and Cusset (1988a), Rutishauser and Huber (1991), Lobreau-Callen et al. (1998: pollen), Rutishauser and Grubert (1993 [Mourera], 2000 [Apinagia]), Passarelli (2002: pollen), Suzuki et al. (2002: seedlings), Sehgal et al. (2002: seeds, etc.), Ameka et al. (2002: general), Koi and Kato (2003: roots, 2007: hypotheses on nature of shoots and leaves), Jäger-Zürn (2003: apical septum, 2005b: interpretation of the thalloid plant body, 2007: shoot apex; 2011: possible new characters), Rutishauser et al. (2004: Diamantina), Rutishauser and Moline (2005: emphasis on "homology"), Jäger-Zürn et al. (2006: microsporogenesis), Cook and Rutishauser (2006: general), Sehgal et al. (2007: organ identity), Jäger-Zürn (2008: Thelethylax), Kato (2008: general), Thiv et al. (2009: African Podostemoideae), Ghogue et al. (2009: Djinga, morphology), Koi and Kato (2010: vegetative body, Hydrodiscus et al.), and de Sá-Haiad et al. (2010: floral morphology of Podostemon).
Phylogeny. the basic phylogenetic structure of the family is [Tristichoideae [Weddellinoideae + Podostemoideae], all aldes with very strong support (Kita & Kato 2001; see also Kita 2002: phylogeny and morphology; esp. Koi et al. 2012). See Moline et al. (2007) for the phylogeny and evolution of African Podostemoideae, Koi and Kato (2010) that of Asian Podostemoideae, and Koi et al. (2009) for that of Tristichoideae (and the description of a distinctive new genus), and Tippery et al. (2011) for that of New World Podostemoideae, where morphological analyses allowed the recovery of a few small generic clades, but molecular data resolved quite a number of nodes. The recently-described Diamantina appeared to be sister to all other Podostemoideae (Ruhfel et al. 2009), although that genus was not studied by Tippery et al. (2011), who found Podostemon and a paraphyletic Mourera as successively sister to the rest. Koi et al. (2012: very good sampling) confirmed the position of Diamantina, although support was weak; further large scale structure in Podostemoideae mostly had very little support, and monophyly of genera seems to be almost a foreign concept there, while groupings in Tristichoideae are well supported and genera are monophyletic.
Classification. Many genera are monotypic, the morphology of the thallus being so bizarre. For generic limits in some African Podostemoideae, see Thiv et al. (2009).
Previous Relationships. Prior to molecular work, systematists were largely at a loss as to where the relationships of Podostemaceae were to be found. The micropylar suspensor haustorium seemed like that of Crassulaceae, perhaps suggesting a link between the two families (e.g. Les & Philbrick 1996; Ueda et al. 1997a), but very different positions have also been suggested. Indeed, Podostemaceae have sometimes been set apart from all other angiosperms (e.g. Cusset & Cusset 1988b).
[[[Lophopyxidaceae + Putranjivaceae], Caryocaraceae, [Centroplacaceae [Elatinaceae + Malpighiaceae]], [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]]] [[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]]] [[Peraceae [Rafflesiaceae + Euphorbiaceae]] [[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]]]]: ovules 2/carpel, apical, pendulous, epitropous.
[[Lophopyxidaceae + Putranjivaceae], Caryocaraceae, [Centroplacaceae [Elatinaceae + Malpighiaceae]], [Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]]]: outer integument 5-7 cells across, inner integument 5-6 cells across.
Evolution. Although ovule number can be optimised to this node, note that both the position and orientation of the two ovules varies...
[Lophopyxidaceae + Putranjivaceae]: stomata paracytic; hairs unicellular; flowers imperfect; style 0; fruit 1-seeded.
Evolution. Divergence & Distribution. This clade may have separated in the Cretaceous-Albian 111-100 m.y.a., Lophopyxidaceae and Putranjivaceae themselves diverging at end Coniacian or thereabouts ca 85 m.y.a. (Davis et al. 2005a).
Phylogeny. Davis et al. (2005a) found a strong association between these two families; they may in turn be associated with the group of families with parietal placentation. However, the position here has strong support in Xi et al. (2012: = putranjivoids).
LOPHOPYXIDACEAE H. Pfeiffer Back to Malpighiales
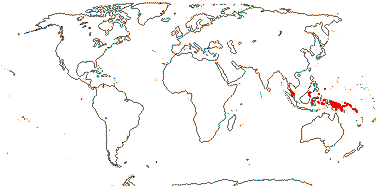
Lianes, climbing by leaf tendrils; chemistry?; branches with lateral bud at base; secondary thickening anomalous, with included phloem; vessel elements with simple perforation plates; phloem stratified; nodes ?; petiole bundles arcuate and with wing bundles; leaves spiral; plant monoecious; inflorescence branched, flowers in clusters; flowers sessile, small; K connate basally, valvate, C small, shorter than K, cordate or reniform nectary glands adnate to C; staminate flowers: stamens = and opposite sepals; carpellate flowers: nectary glands connate at the base; staminodes 0; G [(4) 5], opposite petals, stigmas subulate; ovules with small funicular obturator, ?integument; fruit a 5-winged samara; seed coat?; endosperm ?development, +, cotyledons long; n = ?
1/1: Lophopyxis maingayi. Malesia to the Solomon and Caroline Islands (map: from Sleumer 1971b).
Evolution. Divergence & Distribution. The diversification rate in Lophopyxidaceae seems to have slowed down (Xi et al. 2012).
Chemistry, Morphology, etc. Lophopyxis lacks information on everything from gross morphology to embryology. Sleumer (1971b) described the tendrils as being leaves and also bud-bearing branches; the ultimate spirally-recurved portion seems to be foliar.
For general information, see Kubitzki (2013).
Previous Relationships. Lophopyxidaceae were included in Celastraceae by Cronquist (1981) and Hutchinson (1973), in Celastrales by Takhtajan (1997), but placed close to Pandaceae (represented by Microdesmis) by Savolainen et al. (2000a; see also Chase et al. 2002).
PUTRANJIVACEAE Meisner Back to Malpighiales
Evergreen trees; cucurbitacins [triterpenes], glucosinolates, biflavonoyls +; cork?; vessel elements with scalariform perforation plates; petiole bundles elliptic; leaves two-ranked, (lamina with veins running into opaque deciduous teeth, or spines); plant dioecious; inflorescence fasciculate; P +, whorled, 4-5(-7), staminate flowers: A (2-)3-20(-many; extrorse); nectary + or 0; carpellate flowers: G [(1-)2-4(-9)], stigmas flap-like, ?type; ovules with micropyle (exo/)endostomal, outer integument 3-9 cells across, inner integument 6-14 cells across, (integument single, 6-9 cells across - Drypetes macrostigma), endothelium +, parietal tissue ca 2 cells across, disintegrating early, placental obturator +, hypostase massive; megaspore mother cells 2-3; fruit a drupe; testa vascularized, exomesotesta sclereidal, tegmen ± multiplicative, 6-24 or more cells thick, exotegmic cells sclereidal, flat-lying; endosperm copious; n = (19) 20 (21).
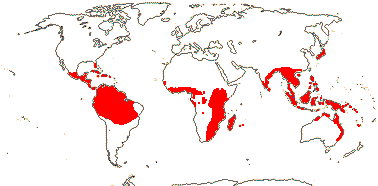
3[list]/210: Drypetes (200). Tropical, esp. Africa and Malesia (map: from FloraBase 2005; Andrew Ford, pers. comm). [Photo - Flower, Fruit]
Evolution. Divergence & Distribution. the age of Putranjivaceae is some 58 m.y. (Wikstrom et al. 2001). Some analyses suggest that the diversification rate in Putranjivaceae increased (Xi et al. 2012).
Evolution. Plant-Animal Interactions. Perhaps not surprisingly, caterpillars of pierid butterflies have quite often (23/2690 records) been recorded eating Putranjivaceae (see also Brassicales and Fabaceae) - nothing so far is known about Lophopyxidaceae - species of the Indo-Malesian Appias subgenus Catophaga (albatrosses) feeding more or less indiscriminately on Drypetes (Putranjivaceae) and Capparaceae (Yata et al. 2010). In Drypetes natalensis, at least, consitutively-released isothiocyanates are part of the floral odour, but what exactly they do is unclear (Johnson et al. 2009a).
Chemistry, Morphology, etc. For general information, see Levin (2013), for chemistry, Hegnauer (1966, 1989, as Euphorbiaceae), for wood anatomy, Hayden and Brandt (1984 - it is like that of Aporosa, etc. [= Phyllanthaceae]), for embryology and seed anatomy, Singh (1970), Stuppy (1996), and Tokuoka and Tobe (1999, 2001 - Lingelsheimia included, but tegmen 3-4 cells thick and testa vascularized, to be placed in Phyllanthaceae - see Kathriarachchi et al. 2005). For a checklist and bibliography, see Govaerts et al. (2000).
Previous Relationships. Putranjivaceae have usually been included in Euphorbiaceae (as by Webster 1994b, in Phyllanthoideae), but can be distinguished i.a. by their chemistry, embryology, and fruit. They are certainly not to be placed with the rest of the glucosinolate families in Brassicales (e.g. Rodman et al. 1997, 1998).
CARYOCARACEAE Voigt, nom. cons. Back to Malpighiales
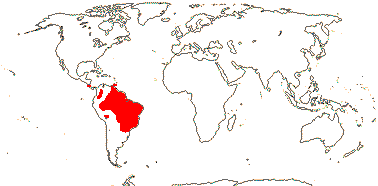
Evergreen trees to shrubs; ellagic acid, triterpenoid saponins + [Caryocar]; vessel elements with simple (scalariform) perforations; nodes 5 or more:5 or more; branched sclereids +; cuticle waxes as smooth to irregular platelets; colleters +; leaves opposite [Caryocar] or spiral, trifoliate, leaflets ± articulated, (margins entire), stipellate or not, stipules ± intrapetiolar [Anthodiscus] or inter-intrapetiolar; inflorescences terminal, racemose(-corymbose); pedicels articulated, bracts 0; flowers large [>5 cm across], (6-merous), K imbricate, (small, ± connate, lobed - Anthodiscus), C protective in bud, with 3 traces, connate below, or forming a deciduous calyptra; A many, connate at base and adnate to C, (in 5 bundles), filaments long, with wart-like tubercles/vesicles towards apex, inner stamens staminodial, anthers basifixed; nectary at base of G/0 [Anthodiscus]; G [4-20], placentation basal, styluli impressed, with 2 vascular bundles, stigmas puctate-impressed; ovule 1/carpel, basal, erect, campylotropous to anatropous, sessile, attachment broad, micropyle endostomal, outer integument 2-3 cells across, inner integument 3-4 cells across, (unitegmic, integument 4-5 cells across - Anthodiscus), weakly crassinucellate, nucellus below embryo sac massive, epidermal cells of nucellar apex radially elongated, obturator 0; fruit a drupe, (radiating fibres - Caryocar), stone separating into 1-seeded units; seeds reniform, hilum large; coat undistinguished, testa vascularized, aerenchymatous or not, exotegmen?; endosperm type?, at most thin, (hypocotyl very large, oily, spirally-twisted - Anthodiscus); n = 23.
2[list]/21: Anthodiscus (15). Tropical America, esp. Amazonia (map: from Prance & Freitas da Silva 1973). [Photo - Flower] [Photo - Flower, Fruit]
Evolution. Divergence & Distribution. Stem Caryocaraceae may have diverged from other Malpighiales in the Cretaceous-Albian 111-100 m.y.a. ([117.6-]112.3[-108.5]/[103.8-]101.7[-99.5] m.y.: Davis et al. 2005a).
Chemistry, Morphology, etc. .he fruits of both genera can be used as fish poisons.
Prance and Freitas da Silva (1973) described Anthodiscus as lacking stipules. See Dickison (1990c) for details of the complex floral vasculature and other floral features; in Anthodiscus each style receives a vascular bundle from adjacent carpels, so is presumably commissural, while the style of Caryocar is vascularized from a single carpel. The androecium may form a ring primordium (Ronse de Craene & Smets 1992b).
See Prance (2013) for general information, Hegnauer (1964, 1989, the latter also under Lecythidaceae) and Chisté and Mercadante (2012) for chemistry.
Previous Relationships. Both Cronquist (1981) and Takhtajan (1997) included Caryocaraceae in Theales.
[Centroplacaceae [Elatinaceae + Malpighiaceae]]: pedicels articulated; K persistent in fruit.
Phylogeny. for relationships in this area, see Zhang et al. (2009a, 2009b, 2010) and Wurdack and Davis (2009); Xi et al. (2012: = malpighioids) found that support for the inclusion of Centroplacaceae in this clade was only weak (63% ML bootstrap; 0.51 PP).
CENTROPLACACEAE Doweld & Reveal Back to Malpighiales
Evergreen trees; inflorescence branched; A 5, opposite sepals; styles widely diverging, stigmas little expanded; ovules collateral; capsule with one seed/loculus; seeds arillate, exotegmic cells laterally compressed and ribbon-shaped, thick-walled; embryo short.
2[list]/6. West Africa, Indo-Malesia.
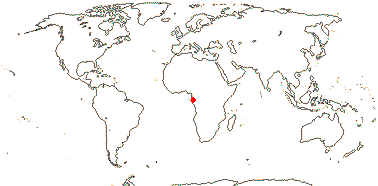
1. Centroplacus
Chemistry?; cork?; vessel member perforations?; sclereids +; stomata anisocytic; leaves two-ranked, stipules cauline; plant dioecious; flowers small; nectary? lobes alternating with K and outside A; staminate flowers: anther dehiscence oblique-apical, connective well developed; pollen psilate, perforate; pistillode +; carpellate flowers: C 0; ?staminodes minute; G [3]; ovules subapical, ?morphology; fruit a septicidal (and loculicidal) capsule opening from the base; seed carunculate; exotesta rather tall, outer wall thickened, mesotegmic cells flattened and at right angles, endotegmen ± thick-walled; n = ?
1/1: Centroplacus glaucinus. W. Africa.
2. Bhesa
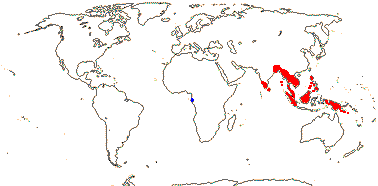
Chemistry?; (cork mid-cortical); vessel elements with scalariform perforation plates; paratracheal parenchyma +; nodes 5:5; calcium oxalate as crystals [?always]; petiole with bundles forming a U or flattened-annular, 2-3 medullary bundles, (also wing bundles); stomata laterocylic; leaves spiral, lamina vernation conduplicate, margins entire, tertiary venation closely scalariform, petiole ± pulvinate apically, stipules almost encircling the stem, colleters +; inflorescence racemose; C contorted; A extrorse to introrse; pollen finely striate; nectary lobed or not; G [2]; ovules basal, apotropous, micropyle exostomal, outer integument 6-8 cells across, inner integument 4-5 cells across, hypostase +; fruit loculicidal; aril exostomal-funicular, sheet-like; exotegmic cells massive, exotegmen cells laterally compressed tracheidal; n = ?; germination epigeal.
1/5. Indo-Malesia (map: from Ding Hou 1962).
Evolution. Divergence & Distribution. Centroplacaceae may be sister to Ctenolophonaceae, diverging in the Cretaceous-Albian 111-100 m.y.a. ([109.6-]101.8[-96.6]/[97.1-]91.0[-88.1] m.y.: Davis et al. 2005a). Xi et al. (2012) found that diversification rate in this clade slowed down in some analyses.
Bacterial/Fungal Associations. Bhesa is reported to be ectomycorrhizal (Smits 1994).
Chemistry, Morphology, etc. The ribbon-shaped exotegmic cells of Bhesa are longer than those of Centroplacus, and its integument is much thicker. The endostome of Centroplacus is lignified and more or less protruding.
For general information, see Kubitzki (2013b), Pierre (1894), Ding Hou (1962, both Bhesa, and Wurdack and Davis (2009). For seed and vegetative anatomy of B. ceylanica, see Jayasuriya & Balasubramaniam 3107, for seeds of B. robusta, see Corner (1976). For more information about Centroplacus, see Forman (1966: general), Stuppy (1996: seed anatomy and good discussion, not Euphorbiaceae s.l.), Tokuoka and Tobe (2001: seed anatomy, Euphorbiaceae-Phyllanthoideae, but with some doubt), and Radcliffe-Smith (2001: generic description). However, studies of the embryology, etc., of both genera are much needed.
Phylogeny. In a molecular study by Wurdack et al. (2004) Centroplacus was associated with Pandaceae, although with very little support, however, in Davis et al. (2005a) it is separate from Pandaceae and weakly associated with Ctenolophonaceae. Zhang and Simmons (2006) found that Bhesa fell among the few Malpighiales they included in their analysis of Celastrales, and Ken Wurdack (pers. comm.) suggested that a position around about here may be appropriate.
Classification. Recognising the two genera as a family seems most reasonable.
Previous Relationships. Although Centroplacus glaucinus was often placed in Pandaceae (Takhtajan 1997; Mabberley 1997), Webster (1994b) and Radcliffe-Smith (2001) included it in Euphorbiaceae, but only with hesitation and with little certainty as to where it should be placed within the family. There is no obturator, unlike Euphorbiaceae.
Bhesa was distinctive in morphological analyses of Celastraceae, in which it used to be included (Simmons & Hedin 1999; Matthews & Endress 2005b), if sometimes with some doubt (e.g. Pierre 1894 [he thought it might be in a separate family]; Metcalfe & Chalk 1950; Ding Hou 1962). Its huge stipules, distinct styles, vessels with scalariform perforation plates, etc., were somewhat out of place there, although Celastraceae were so heterogeneous that a strong case could not be made for its removal. The seed coat, with its massive exotegmic cells, is also very different, as is its pentalacunar nodes.
[Elatinaceae + Malpighiaceae]: vessel elements with simple perforation plates; sieve tube plastids lacking starch and protein inclusions; leaves opposite, with glands[?], (lamina margins entire); inflorescence cymose; flowers with inverted orientation; nectary 0; when G 3 median member adaxial, ?integument; fruit septifragal; endosperm slight; x = 6; duplication of CYC genes [?level].
Evolution. Divergence & Distribution. The common ancestor of these two families may have diverged towards the beginning of the Cretaceous-Albian at around 111-100 m.y., the Malpighiaceae and Elatinaceae themselves separating towards the end of this period some [113-]98[-89]/[100-]89[-85] m.y.a. (Davis et al. 2005a).
Depending on the interpretation of variation, both the evolution of monosymmetric flowers and flowers with an inverted orientation can be pegged to this node (Zhang et al. 2010).
Chemistry, Morphology, etc. For the foliar glands and resin and latex production in Elatinaceae and Malpighiaceae, neither well understood, see Vega et al. (2002) and Davis and Chase (2004). Vega et al. (2002) suggested that the laticifers of Galphimieae might be a symplesiomorphy with Euphorbiaceae, although laticifers are not even an apomorphy of that family which anyhow is not immediately related to this group (see below); I know nothing of the composition of the latex of Malpighiaceae. Zhang et al. (2009a) surmised that the flowers of the ancestor of [Elatinaceae + Malpighiaceae] might have been monosymmetric.
Phylogeny. For relationships between Malpighiaceae and Elatinaceae, see Davis and Chase (2004), Tokuoka and Tobe (2006), Korotkova et al. (2009), Wurdack and Davis (2009), Wang et al. (2009), Xi et al. (2012), etc. - support is strong.
Previous Relationships. It had been suggested that Malpighiaceae were rather weakly associated with Peridiscaceae and were perhaps near Clusiaceae et al. (Chase et al. 2002); for the current position of Peridiscaceae, here in Saxifragales, see e.g. Davis and Chase (2004).
ELATINACEAE Dumortier, nom. cons. Back to Malpighiales
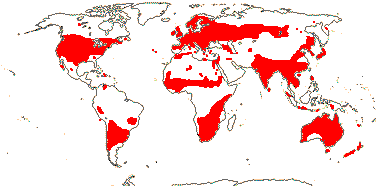
Herbs to subshrubs of moist/wet habitats; flavonols, ellagic acid +; plant resinous; (cork from inner cortex); nodes 1:1; mucilage cells +; leaves with colleters, stipules scarious; ?pedicel articulation; flowers (single), (2-)5-6-merous, K free to connate, (with an apical "gland"), C contorted; stamens = and opposite sepals, or 2x K; (pollen trinucleate - Elatine); G [2-5], opposite sepals, stigma papillate; ovules many/carpel, micropyle zigzag, outer integument 2-3(-5) cells across, inner integument 2-3 cells across, parietal tissue ca 4 cells across; megaspore mother cells several; fruit capsular; exotegmic cells with low lignified sinuous anticlinal walls; embryo ± fusiform; n = (9), x = 10 (?14, ?5).
2[list]/35: Bergia (25), Elatine (10). Worldwide, most tropical, not arctic (map: from Meusel et al. 1978; FloraBase 2006).
Chemistry, Morphology, etc. Eichler (1878) draws three-merous flowers of this family with the odd sepal abaxial, i.e., in the monocot position, and Zhang et al. (2010: Supplement 1) consider that this orientation also occurs in flowers of the 5-merous Bergia texana.
See Kubitzki (2013b) for general informatio, Tobe and Raven (1983b) for some embryology.
Synonymy: Cryptaceae Rafinesque
MALPIGHIACEAE Jussieu, nom. cons. Back to Malpighiales
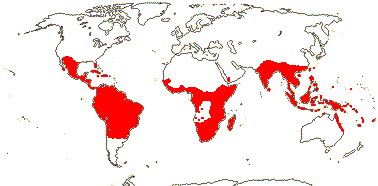
Lianes to trees; (inulin +), ellagic acid 0; (cork ?near endodermis); secondary thickening often anomalous; pits vestured; (nodes 1:1); petiole bundle arcuate; cuticle waxes as rosettes; stomata usu. paracytic; branching from current flush; hairs unicellular, ± T-shaped, surface rough; leaves (spiral), glands common, abaxial or petiolar, stipules cauline, intrapetiolar or petiolar (0); inflorescence various; (flowers polysymmetric); K with large paired abaxial oil glands (0 - esp. Old World taxa), C clawed, often crumpled in bud, often fringed, one adaxial-lateral often different to the others; A (2; = and opposite sepals) 10, obdiplostemonous, (15), often basally connate; tapetal cells multinucleate; G [(2) 3(-5)], (inferior), styles +, (style single), stigma dry; ovule 1/carpel, apical, pendulous. apotropous[?], micropyle exo/endo/bistomal or none, outer integument 2-4 cells across, inner integument 3-6 cells across, nucellar beak +; embryo sac often tetrasporic, 16-nucleate, (bisporic, 8-nucleate); K and A often persistent in fruit (K accrescent, wing-like); (fruit separating into mericarps); exotegmen 0 (fibrous - Thryallis), endotegmic cells (elongated), lignified; chalazal endosperm haustoria +, (embryo curved); n = 6 (?5), nuclear genome (0.069-)0.918(-12.261) pg.
68[list]/1250 - two groups below. Tropical and subtropical, especially American (map: C. C. Davis, from Arènes 1957). [Photo - Flower, Flower, Fruit.]
1. Byrsonimoideae W. R. Anderson
(Articulated laticifers + - Galphimieae); style subulate, stigma terminal; (fruit baccate); n = 6.
Byrsonima (150). American Tropics.
2. Malpighioideae Burnett
Pollen globally symmetric [4-polyporate]; style various, stigma usu. not terminal, asymmetrically capitate; fruit winged, (bristly, unwinged); n = (9) 10.
Malpighia (?130), Heteropterys (120), Stigmaphyllon (100), Banisteriopsis (90), Bunchosia (55), Mascagnia (50), Malpighia (40). Tropical and subtropical, especially the Americas.
Evolution. Divergence & Distribution. Malpighiaceae are known fossil from the Northern Hemisphere, e.g. from the Eocene Claiborne Formation in Tennessee, U.S.A., in deposits ca 34 m.y. old (Taylor & Crepet 1987: Eoglandulosa, see also Friis et al. 2011). The rate of diversification may have increased in Malpighiaceae (Xi et al. 2012). An origin of the family in South America during the Late Cretaceous (ca 68 m.y. before present) has been suggested, with several - it now appears to be nine - subsequent dispersal events to the Old World followed by the loss of oil secretion by the sepals (Davis et al. 2002a, b, 2004; esp. Davis & Anderson 2010).
Ecology. Malpighiaceae are one of the three ecologically most important groups of lianes in the New World tropics (see also Bignoniaceae-Bignonieae and Sapindaceae-Sapindoideae: Gentry 1991).
Pollination Biology. The family is noted for having oil flowers, a trait that may have evolved 75-64 m.y.a. (Renner & Schaefer 2010). Oil is secreted by paired calyx glands (epithelial elaiophores) and removed by the legs of the bees. Several genera of Apidae and Centridini-Anthophoridae are involved, and the latter, at least, grasp the narrow base of the banner petal of the functionally inverted monosymmetric flowers with their mandibles as they collect the oil from the glands, which are on the abaxial surface of the sepals; this banner petal is often distinctively coloured, and may change colour as it ages (Renner & Schaefer 2010 and references). Oil is secreted in New World taxa only; some taxa there are buzz pollinated (Sigrist & Sazima 2004). The flowers of Oncidiinae orchids may mimic those of Malpighiaceae, and sometimes they even provide fatty acids as a reward that are similar to those of Malpighiaceae (Reis et al. 2007). Galphimia brasiliensis may have small glands on the sides of the sepals towards the base; these glands have the same anatomy as glands on the margins of the leaf towards the base, and both secrete oil (Castro et al. 2001). However, Lobreau-Callen (1989) recorded the leaf/bracteole glands of G. bracteata as producing sugars.
In some Malpighiaceae growing in the Old World the "oil" glands secrete nectar (Vogel 1974, 1990), although in most pollen is the only obvious reward (Davis & Anderson 2010); the anthers may be porose. The orientation of the flowers has sometimes reverted to the normal condition for a core eudicot with the odd petal abaxial (Zhang et al. 2010). Flowers of Old World Malpighiaceae tend to be more or less polysymmetric, the petals are less strongly clawed, and the style branches longer and more widely spreading (Davis & Anderson 2010 for illustrations).
Zhang et al. (2010) suggested that the functional inversion of the flower in New Worl Malpighiaceae is because of a 36o rotation of the flower; the result is that monosymmetry is associated with a monocot-type orientation of the flower, with the odd member of the inner whorl adaxial. There have been duplication(s) of CYC genes, although there is no local genome duplication, and CYC2 genes are expressed only in the adaxial part of the flower in monosymmetric Malpighiaceae - although details of exactly when gene duplication occurred and what changes in pattern of gene expression there have been are unclear (Zhang et al. 2010).
Self-fertilization is common in species of Gaudichaudia, Janusia and relatives; it occurs by pollen tubes growing through the tissues of the flower to the embryo sac (Anderson 1980). Apomixis - nucellar polyembryony - is common.
Chemistry, Morphology, etc. Acridocarpus has spiral, exstipulate leaves. Some species of Stigmaphyllon have leaves with palmate venation and toothed margins; some taxa, especially when young, have almost fimbriate lamina margins, albeit distantly so (the fimbriae are ca 4 mm long). Stipules are very diverse, being petiolar in Hiraea and cauline in many vines and also in Malpighia; in the latter genus they may be lobed or toothed.
The flowers of Acridocarpus have an inferior ovary with only two fertile carpels. Testa anatomy may repay investigation. In Mascagnia macrodisca there is a layer of thin-walled, slightly elongated cells with brown contents over a layer of more or less isodiametric, lignified cells with somewhat more thickened and straight anticlinal walls; the latter layer has a "frosted" appearance; the origin of these cell layers is not known (pers. obs.). Given that the main protective part of the propagule is not the seed coat, testa and tegmen may be rather reduced. There are reports that endotegmic fibres may be quite conspicuous (Silva & Trombert 2006), but since fibres in other Malpighiales are exotegmic and Elatinaceae also have exotegmic seeds, what is going on in Malpighiaceae is unclear.
Some information on chemistry is taken from Hegnauer (1969, 1989: iridoids have been reported from Stigmatophyllum) and on fruit and seed is taken from Takhtajan (2000). C. Anderson et al. (2006 onwards) provide general information, especially phylogeny and nomenclature, on the whole family.
Phylogeny. Information on relationships within the family is taken from Davis et al. (2001) and Cameron et al. (2001); within Malpighiodeae, for the most part well-supported branches including Acridocarpus, McVaughia, Barnebya, Ptilochaeta, Bunchosia and Hiraea are successively sister to the remaining taxa. A recent study by Davis and Anderson (2010: four genes, all genera sampled) returned the same set of relationships, and with good support.
Previous Relationships. Malpighiaceae were included in Vochysiales by Takhtajan (1997) and in Polygalales by Cronquist (1981).
[Balanopaceae [[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]]: hairs simple [unicellular always?]; ovules collateral, micropyle bistomal, outer integument ³5 cells across, inner integument ³5 cells across, nucellus evanescent by maturity, endothelium +; endosperm at most slight.
Evolution. Divergence & Distribution. This clade may have diverged at the beginning of the Cretaceous-Albian 111-100 m.y.a., Balanopaceae separating from the rest at the end of this period ([106.2-]99.6[-95.5]/[94.9-]90.2[-88.5] m.y.: Davis et al. 2005a).
Chemistry, Morphology, etc. For a comparison of floral morphology within the whole clade, see Matthews and Endress (2006a); for ovule position, see Merino Sutter and Endress (2003). Tobe and Raven (2011) suggested that there is a multiplicative inner integument; although it is thick, it does not usually become thicker after fertilization. There is insufficent information to know if a vascularized testa might be an apomorphy for the clade.
Phylogeny. For relationships, see especially Litt and Chase (1999); this group (= chrysobalanoids: Xi et al. 2012) has held together strongly in subsequent studies.
BALANOPACEAE Bentham & J. D. Hooker, nom. cons. Back to Malpighiales
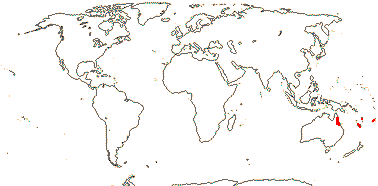
Evergreen trees; ellagic acid?, prob. tanniniferous; vessel elements with scalariform perforations; parenchyma diffuse; sclereid nests with rhomboidal crystals in bark; petiole bundle?; bud scales +; cuticle waxes 0 (platelets); stomata usu. laterocytic; leaves spiral, lamina tooth ?type, stipules minute; plant dioecious; staminate plants: inflorescence catkinate; P +, of small teeth; A (1-)3-6(-14), anthers much longer than filaments; pollen 3-5-colpate, exine columellate-granulate; pistillode common; carpellate plants: inflorescence a fascicle; flowers with cupule made up of spirally arranged bracts; P 0; G [(2)], styles long, once or twice bifid, stigma adaxial on styles, ?type; ovules subbasal, apotropous but facing laterally, outer integument 5-7 cells across, inner integument 5-9 cells across, parietal tissue ca 2 cells across; fruit a drupe with 2-3 stones; testa vascularized, persistent, cell walls not much thickened; endosperm type?, slight, embryo green, large, cotyledons cordate; n = 20 (21), x = 7 (?6, ?11); germination epigeal, phanerocotylar.
1[list]/9. S.W. Pacific, especially New Caledonia (map: from van Steenis & van Balgooy 1966). [Photo - Fruits © Andrew Ford, CSIRO.]
Evolution. Divergence & Distribution. Some analyses suggest that the diversification rate in Balanopaceae decreased (Xi et al. 2012).
Chemistry, Morphology, etc. The leaves are often described as being dimorphic (Carlquist 1980; Cronquist 1983), but they are no more so than in many plants that have bud scales. Cristarque cells occur in some species. Endress and Merino Sutter (2002) present details of the morphology of carpellate flowers; pollen descriptions in Feuer (1991) and Herendeen et al. (1995) differ somewhat.
see Kubitzki (2013b) for general information, Batygina et al. (1991) for testa anatomy, and Merino Sutter and Endress (2003) for floral morphology. Little is known about embryology.
Previous Relationships. Relationships of Balanopaceae have long been problematic. Cronquist (1983) compared their wood anatomy with that of Hamamelidaceae; Balanopales were included in Daphniphyllanae by Takhtajan (1997). Merino Sutter and Endress (2003) suggested that the floral morphology of Balanopaceae is closer to that of Euphorbiaceae than to other families of this group.
[[Trigoniaceae + Dichapetalaceae] [Euphroniaceae + Chrysobalanaceae]]: vessel elements with simple perforation plates; vestured pits +; mucilage cells +; stomata paracytic; lamina margins entire, (flat surface glands or glandular hairs +); pedicels articulated; flowers obliquely monosymmetric; K basally connate, with epidermal mucilage cells, quincuncial, 2 outer members shorter; fertile stamens abaxial, ± connate, anthers with a little pit where the filament joins, connective well developed abaxially with endothecium continuous there, staminodes adaxial; G with longitudinal furrows, unicellular unlignified hairs +, style +, stigmas commissural; ovules with zig-zag micropyle, outer integument 2-5 cells across, inner integument 3-8 cells across, parietal tissue?, obturator +.
Chemistry, Morphology, etc. Matthews and Endress (2008), which see for many more details, elaborate the floral morphology of this clade and suggest synapomorphies for its members.
Previous Relationships. Including these four families in Chrysobalanaceae s.l. was optional in A.P.G. II (2003), and specimens of Chrysobalanaceae and Dichapetalaceae are quite often misidentified as the other family (G. T. Prance, pers. comm.), however, the families are kept separate in A.P.G. III (2009).
[Trigoniaceae + Dichapetalaceae]: (vessel elements with scalariform perforations); petiole bundle arcuate; lamina with strongly looping secondary veins; inflorescences cymose; K with mesophyllar mucilage cells; nectary with lobes or scales, semi-annular [staminodial?]; ovary and lower style completely synascidiate; outer integument <5 cells across; testa multiplicative.
TRIGONIACEAE A. Jussieu, nom. cons. Back to Malpighiales
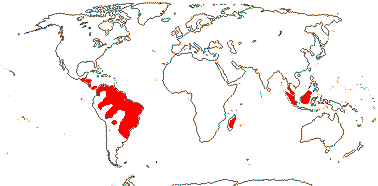
Evergreen trees or lianes; helical thickening in ray and axial parenchyma; wood fluorescing [1 sp]; parenchyma in apotracheal bands; branched sclereids +; (nodes with split laterals); hairs T-shaped, unicellular[?]; leaves opposite, spiral or two-ranked, lamina with dense whitish hairs below (not), stipules interpetiolar [when leaves opposite]; (inflorescence racemose - Isidodendron); C contorted, adaxial-lateral petal is basally spurred or saccate, the standard, plicae in abaxial + abaxial-lateral petals form the keel, or these petals saccate; A 5-13, filaments ± connate, fertile stamens 4-9, staminodes 2-6; pollen 3-5-porate; nectary of 1 or 2 [and then each to 3-lobed] glands at base of standard (glands on base of staminodes - Isododendron); G [(4)], median member adaxial, placentation also parietal, stigma capitate to slightly trilobed; ovules 1-10/carpel, (apotropous), micropyle zig-zag, outer integument 2-3 cells across, inner integument 4-6 cells across, whole inner integument endothelial; fruit a septicidal capsule, carpels opening internally, central fibrous strands persisting, (hairs from endocarp- Trigoniastrum), or samara; seeds (winged); tegmen also multiplicative, exotesta with thickened outer walls, with long lignified hairs or not, endotegmic cells tanniniferous, walls slightly thickened; endosperm development?; embryo green [Trigonia], cotyledons large; n = ca 10; germination epigeal, phanerocotylar.
5[list]/28: Trigonia (24). Central and South America, Madagascar (Humbertiodendron), W. Malesia (Trigoniastrum) (map: from van Steenis 1949c; Lleras 1978). [Photo - Flower]
Chemistry, Morphology, etc. Trigonia has opposite leaves, interpetiolar stipules, and split lateral vascular bundles; lamina glands are not obvious. The bracts of Trigoniastrum are more or less glandular, with large glands on the abaxial surfaces, Trigonia has stalked glands variously on pedicels or petioles and margins of bracts and leaves, while Humbertiodendron has concave marginal glands towards the base of the leaf blades.
For floral morphology, I follow the interpretations of Warming (1875) and Eichler (1878) rather than that of Cronquist (1981). Schnizlein (1843-1870: fam. 233) draws the flowers as being more or less vertically monosymmetric, while the orientation shown by Schatz (2001) is difficult to work out; in the latter it appears that the corolla may be quincuncial. Warming (1875) draws the nectary glands as being part of the androecial whorl; Lleras (1978) describes them as being "disc glands". In any event, the androecium at least sometimes seems to have more than 10 stamens. Testa anatomy is similar to that of Linaceae, where there is similar variation in ovules but the two are not immediately related.
Some information is taken from Lleras (1978), Takhtajan (2000), Fernández-Alonso et al. (2000) and Bittrich (2013); see Hegnauer (1973) for chemistry.
Previous Relationships. Trigoniaceae were included in Vochysiales by Takhtajan (1997), while Cronquist (1981) placed the family in Polygalales.
DICHAPETALACEAE Baillon, nom. cons. Back to Malpighiales
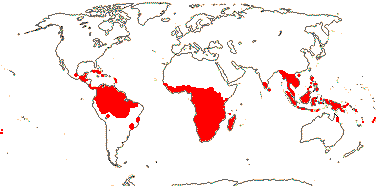
Evergreen trees or lianes; sieve tubes with non-dispersive protein bodies; parenchyma ± paratracheal; pericyclic sheath interrupted; fibres common; branching from current flush; hairs warty[?]; leaves spiral, (lamina with flat abaxial glands), stipules fimbriate or not; inflorescence epiphyllous, from petaiols, (not); flowers small, (polysymmetric), (4-merous); C (connate; 3 small petals forming a "lip"), petals bifid (unlobed), drying black; nectary a ring, or lobes opposite petals; A = and opposite sepals, (3 + 2 staminodes; connate; adnate to C), (anthers without pits); pollen "small"; G [(2-4)] (inferior), (styles separate), stigmas ± punctate, wet, papillate; ovules with outer integument 3-5 cells across, inner integument 6-8 cells across, hypostase 0, funicular obturator +; fruit a flattened drupe, 1(-3, then often lobed) locular; 1 seed/loculus; testa vascularized, only enlarged tanniniferous (divided) exotestal cells and remains of vascular bundles persist, exotegmen 0; embryo green or orange, oily; n = 10, 12.
3[list]/165: Dichapetalum (130: often very poisonous because of fluoracetic acid). Pantropical, few in Malesia (map: see Prance 1972b; Leenhouts 1957a; van Steenis 1963; Heywood 1978 [Africa]). [Photo - Flower] [Photo - Fruit]
Evolution. Divergence & Distribution. Some analyses suggest that the diversification rate in Dichapetalaceae may have increased (Xi et al. 2012).
Chemistry, Morphology, etc. Dichapetalum at least has fluoracetic and related acids in its seed oils (e.g. Badami & Patil 1981). The petiole bundles are arcuate above the insertion of the inflorescence. The flowers of Tapura in particular are quite complex (Prance 1972b).
Some general information is taken from Prance (1972b, inc. anatomy, 2013); see also Hegnauer (1966, 1989) for chemistry, Barth (1896) for petiole anatomy, Punt (1975) for pollen morphology, and Boesewinkel and Bouman (1980) for ovule and seed.
Previous Relationships. Dichapetalaceae were placed in Celastrales by Cronquist (1981) and in Euphorbiales by Takhtajan (1997).
Synonymy: Chailletiaceae R. Brown
[Euphroniaceae + Chrysobalanaceae]: hypanthium +, nectary on inside; C clawed, with lignified hairs.
Chemistry, Morphology, etc. Whether or not this clade has a spur in the floral cup is a matter of perspective; I prefer to interpret the flower in many Chrysobalanaceae as having the gynoecium "adnate" to one side of the hypanthium rather than being spurred as appears to be the case in a l.s. of the flower. Hairs have been found in the ovary loculi in flower, but they are not apparent in the capsule, even before it has opened (pers. obs.).
EUPHRONIACEAE Marcano-Berti Back to Malpighiales
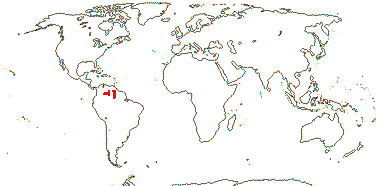
Tree; parenchyma ± aliform-confluent; petiole bundles annular or arcuate, (not joining immediately with stele); cortical and foliar sclereids +; hypodermis mucilaginous; leaves spiral, lamina vernation revolute, white tomentose below; inflorescence terminal; C 3 [abaxial-lateral and abaxial C absent], contorted; stamens = and opposite sepals (4-7), in two groups, adnate to C, filaments basally connate, staminodes 1, long, abaxial-lateral, and 4-5 small and dentate; G with median carpel adaxial, stigma subcapitate; ovule apotropous; fruit a septicidal capsule, columella persisting; seeds winged, coat?; endosperm development?; n = ?
1[list]/1-3. The Guyana Shield, South America (map: from Steyermark 1987).
Evolution. Divergence & Distribution. Some analyses suggest that the diversification rate in Euphroniaceae decreased (Xi et al. 2012).
Chemistry, Morphology, etc. Ovule morphology, etc., of Euphronia is still very poorly known. Lleras (1976) suggested that the (long) staminode of Euphronia was in the position of the fertile stamen of Vochysiaceae (that was a time when the two were thought to be related), Marcano-Berti (1995) that there is a staminal tube and four stamens of two different lengths. Some information is also taken from Warming (1875: general), Barth (1896: general anatomy), and de Pernia and ter Welle (1995: wood anatomy). Some more data come from Euphronia guianensis: Colonnello-Medina 712, vegetative anatomy.
Previous Relationships. Euphronia has been included in Trigoniaceae (Airy Shaw 1966; Hutchinson 1973; Takhtajan 1997) or Vochysiaceae (Cronquist 1981; Mabberley 1997). However, Euphroniaceae and Trigoniaceae differ in a number of features, including those of wood anatomy (see Lleras 1976 for a table) and other aspects of vegetative anatomy (e.g. Trigoniaceae lack the mucilaginous hypodermis of Euphronia: Sajo & Rudall 2002) and are not sister taxa.
CHRYSOBALANACEAE R. Brown, nom. cons. Back to Malpighiales
Evergreen trees or shrubs; trihydroxyflavonoids, distinctive unsaturated fatty acids in the seeds +, ellagic acid 0; (cork ± deep-seated); true tracheids +; wood siliceous and with SiO2 grains; parenchyma in apotracheal bands; nodes 5:5; petiole vasculature annular, often with medullary plates, etc., wing bundles +; branching from previous flush; (foliar sclereids +); leaves often two-ranked, lamina vernation (flat-)conduplicate, often with flat abaxial glands, esp. near base of lamina, (margins toothed), (stipules petiolar or intrapetiolar); inflorescence various; (flowers almost polysymmetric); (C 0); A (2-)5-many, usually long-exserted, abaxial members best developed, (connate); pollen also colpate, angled; usu. only abaxial carpel developed, (loculus divided; all three carpels fertile), often borne on side of tube, style ± gynobasic, stigma punctate (3-lobed); ovules ± basal, outer integument 5-12 cells across, inner integument 5-12 cells across, micropyle bistomal (zig-zag); megaspore mother cells several, embryo sac lacking antipodals; fruit a 1-seeded drupe, endocarp densely hairy (not), medium-sized to large; (seed ruminate), testa (multiplicative), vascularized, undistinguished or mesotestal, exotesta collapsed-fibrous, (tanniniferous), tegmen multiplicative; embryo large; n = 10, 11, x = 11 (?10); germination cryptocotylar, hypogeal.
17[list]/460: Licania (170), Hirtella (105), Couepia (70), Parinari (45). Pantropical, especially American (map: from van Balgooy 1993; Prance & Sothers 2003a, b). [Photo - Flower]
Evolution. Seed Dispersal. For seed dispersal, generally by animals, see Prance and Mori (1983).
Chemistry, Morphology, etc. Syllepsis is uncommon both here (Keller 1994), and probably more generally in the whole group of five families.
For more information, see Morvillez (1918: petiole vasculature), Hegnauer (1973, 1990: chemistry), Badami and Patil (1981: seed fatty acids), Tobe and Raven (1984: embryology), Prance and White (1988: nicely illustrated variation in the family) and LaFrankie (2011: field characters).
Phylogeny. A recent molecular study (Yakandawala et al. 2010) yielded poor resolution of relationships, but suggested that groupings apparent in earlier morphological "taximetric" studies (Prance et al. 1969; Prance & White 1988) should be reexamined. There is some support for Atuna being sister to the rest of the family (see also Wurdack & Davis 2009); Sothers et al. (2014) found Parinari and Maranthes to be basal, although Atuna was not included; Couepia and Licania in particular, but also Hirtella, showed more or less extreme dgrees of para/polyphyly.
Previous Relationships. The flowers of some Chrysobalanaceae look rather like those of Prunus, and Chrysobalanaceae and Rosaceae were often considered to be close (e.g. Cronquist 1981; Takhtajan 1997), either as separate families more or less adjacent in the sequence, or Chrysobalanaceae might even be included as a subfamily of Rosaceae. However, there are numerous differences between them (see table in Prance 1972a).
Classification. See Prance (1989: New World Taxa), and Prance and Sothers (2003a, b: world monograph). However, genera are being reorganized, see e.g. Sothers et al. (2014) for Couepia.
Synonymy: Hirtellaceae Horaninow, Licaniaceae Martynov
[[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]]] [[Peraceae [Rafflesiaceae + Euphorbiaceae]] [[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]]]: ?
[Humiriaceae [Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]]]: nectary +; endosperm persistent.
HUMIRIACEAE A. Jussieu, nom. cons. Back to Malpighiales
Trees; ellagic acid +; cork subepidermal; vessel elements with scalariform perforation plates; true tracheids +; vestured pits +; sieve tube plastids with protein crystals and starch; nodes 5-lacunar; petiole bundles annular, with wing bundles; mucilage cells frequent; stomata various; branching from previous flush; leaves often two-ranked, lamina vernation involute, tooth ?type, (margins entire), petiole short, stipules small or 0; inflorescence cymose; K connate, at least at base, quincuncial, (C quincuncial/cochlear; with 3 traces - Vantanea); A 10-many, filaments ± connate at least basally, with interdigitated hairs higher up, forming a tube, (obdiplostemonous), anther thecae with separated superposed loculi, connective broad, prolonged; pollen exine usu. microreticulate; nectary from base of filaments to base of G, prominent, raised, annular; G [(4-7)], opposite sepals (petals), style undivided, stigma slightly lobed, ?type; ovules usu. 1/loculus, (with 5 traces), micropyle exo(endo)stomal, outer integument 2-3 cells across, inner integument 2-3 cells across, parietal tissue 3-6 cells across, nucellar cap +; fruit a drupe, stone operculate, 1- or 2-seeded, surface sculpted, with "resin" cavities; exotestal cells thick-walled, lignified, tegmen multiplicative [ca 5 cells thick], cross layer of fibres beneath exotegmen; endosperm copious [?always], perisperm slight, embryo somewhat curved, green; n = 12.
8[list]/50: Vantanea (16), Humiriastrum (12). Tropical America, W. Africa (Saccoglottis, also American) (map: from Thorne 1973). [Photo - Flower, Fruit.]
Evolution. Divergence & Distribution. Humiriaceae may have diverged from other Malpighiales in the Cretaceous-Albian 111-100 m.y.a. ([117.2-]112.2[-108.9]/[102.8-]101.6[-100.3] m.y.: Davis et al. 2005a). Herrera et al. (2010) rejected all fossils placed in this family other than some from South America, suggesting that the family originated there.
Seed Dispersal. The fruits are dispersed by bats or by water, empty cavities in the stone affording bouyancy.
Chemistry, Morphology, etc. There is no endothelium. Although D. A. Link is sometimes cited as the author of a paper on the nectaries of Humiriaceae, and promised such a paper himself, it seems never to have appeared.
Some information is taken from Herrera et al. (2010: general), Mauritzon (1934d: ovules), Narayana and Rao (1977 and references: floral morphology), Bove and Melhem (2000: pollen), and Boesewinkel (1985a: ovule and seed).
Phylogeny. Within Humiriaceae, Vantanea is sister to the other genera; it has three or more staminal whorls (Bove 1997). Herrera et al. (2010: more detailed morphological analysis of 40 characters) suggested that Vantanea and Humiria were successively sister to the remainder of the family, although support for this topology was weak; the [Humiria + the rest] clade had quincuncial corollas and unilocular anthers.
Previous Relationships. Bove (1997) suggested that Ixonanthaceae were sister to Humiriaceae, both having ellagic acid, a "free" nectariferous disc encircling the ovary, and an undivided style with an entire stigma. Humiriaceae have also been linked with Linaceae and Erythroxylaceae, and thence to Geraniales (Narayana & Rao 1978b), or the three families together are placed in Linales (Cronquist 1981).
[Achariaceae [[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]]: crystals in ray cells; sieve tubes with non-dispersive protein bodies; cuticle waxes usu. 0; (foliar glands +); pedicels articulated; nectary outside A; G with median member abaxial, placentation parietal, placentae raised; ovules several/carpel, nucellus massive; seeds arillate; endotegmen persistent; endosperm oily.
Evolution. Divergence & Distribution. All six major clades in this group (Malesherbiaceae + Turneraceae + Passifloraceae, = Passifloraceae s.l., make a single clade) may have diverged in the Cretaceous-Albian 111-100 m.y.a., or somewhat later (Davis et al. 2005a, details are given for the individual clades); Wikström et al. (2001) suggest that many of the clades do not diverge until (well) after 63 m.y.a..
Plant-Animal Interactions. Larvae of butterflies such as Nymphalidae-Acraeinae and N.-Nymphalinae-Heliconiini, -Vagrantini and -Argynnini commonly eat members of this group (Ehrlich & Raven 1964; see also Dahlgren & van Wyk 1988; Arbo 2006; Simonsen 2006; Silva-Brandão et al. 2008; Nylin & Wahlberg 2008; etc.); this is also discussed under individual families below. Some Acraeinae in particular may cue on the presence of the cyanogenic glucoside gynocardin in potential food plants, indeed, that larvae of Acraea horta, normally living on the woody Kiggelaria africana, ate herbaceous Achariaceae prompted the successful search for that compound in the latter family (Steyn et al. 2002). Toxic compounds like gynocardin may be sequestered by the larva and passed on to the adult. Interestingly, given the relationships evident in Wurdack and Davis (2008), the distinctive cyclopentenoid glycosides seem to have evolved more than once.
Phylogeny. For relationships in this clade (= the parietal clade), which has strong support, see Xi et al. (2012).
Chemistry, Morphology, etc. There is much information on seed anatomy in Takhtajan (1992) while Krosnick et al. (2006) briefly discuss the evolution of polyandry in this group - in some cases, at least, the numerous stamens form a single whorl. See Mauritzon (1936b) for some information on embryology and Furness (2011) for pollen development and ultrastructure.
Previous Relationships. It was commonly agreed that the old Flacourtiaceae presented major taxonomic problems. "Flacourtiaceae as a family is only a fiction; only the tribes are homogeneous" (Hermann Sleumer, the monographer of the family, in Miller 1975: 79) - indeed, it was a fiction. Some of the old Flacourtiaceae are now in Achariaceae, a few in Lacistemataceae, while Flacourtiaceae-Berberidopsideae are in Berberidopsidales (as Berberidopsidaceae) and Aphloia (Aphloiaceae) is in Crossosomatales. The name Flacourtiaceae is now no longer in use, and remainder of this family is placed in Salicaceae. Variation in chemistry, leaf teeth, floral morphology, and seed coat anatomy is largely correlated with this division. When the possession of parietal placentation was considered to be very important, other families such as Caricaceae (Brassicales), Cucurbitaceae (Cucurbitales), etc., might also be grouped here.
ACHARIACEAE Harms, nom. cons. Back to Malpighiales
Shrubs to trees ((climbing) herbs); cyclopentenoid cyanogenic glucosides and/or cyclopentenyl fatty acids [gynocardin], ellagic acid [Kiggelaria] +; vessel elements with simple or scalariform perforation plates; fibres septate; axial parenchyma usu. 0; ray cells with scalariform perforations [?distribution]; petiole bundle annular, with two wing/adaxial strands, (inverted medullary plate - Lindackeria); ?stomata; leaves spiral or two-ranked, lamina margins entire (serrate), (stipules 0), petiole often geniculate; (plant dioecious); inflorescence spicate or cymose (fasciculate); K and C spiral or not, not in a simple alternating relationship, K 2-5, C 4-15 (3-4, connate - Acharieae), often in two series, (adaxial scales +); (nectary 0); A 5-many, opposite petals or irregular, initiation centripetal or simultaneous, (from a ring meristem), anthers basifixed, elongate (barely so - Chiangodendron; dehiscing by pores; locellate); pollen also tricolporoidate; G [2-10], median member?, style (short), branched or not, stigma capitate-peltate; ovules (sessile, attachment to placenta broad, (1/carpel), (straight - Xylotheca, Hydnocarpus, Lindackeria, Scaphocalyx), micropyle endo- or bistomal or zigzag, integuments 3-8 cell layers across, (outer integument lobed), parietal tissue massive, 4+ cells across, nucellar cap +, epistase +, ring/cap of tracheids in chalaza; (megaspore mother cells 2), embryo sac penetrating chalaza, forming a caecum below tracheids, (sac bisporic [chalazal dyad], eight-celled: Allium-type - Acharieae); fruit also a berry; (seed arillate); seed coat thick, pachychalazal [not Hydnocarpus laurifolius], testa multiplicative, vascularized, sarcotestal and with stomata (Acharieae), (with crossing layers in middle - Pangium), (mesotesta with sclerenchyma, palisade layer, and sclereids - Scaphocalyx), endotesta lignified, cells sclereidal (radially elongated), tegmen multiplicative, exotegmic cells elongated, massive, sclereidal; endosperm copious, suspensor 0, embryo green; n = 10, 12, 23.
30[list]/145: Hydnocarpus (40). Pantropical. (map: from Sleumer 1954, 1980; Şerban Procheŝ, pers. comm. [Africa]; Andrew Ford, pers. comm. [Australia])[Photo - Flower, Fruit, Fruit, Acharia tragodes - Leaves.]
Evolution. Plant-Animal Interactions. The feeding behaviour of Acraeini butterfly larvae are consistent with the expanded family limits adopted here (Steyn et al. 2002, 2003 and references). Species of Ryparosa consistently produce food bodies, and in a number there are associations of varying closeness with ants (Webber et al. 2007).
Chemistry, Morphology, etc. There are large and medium intervascular pits; the wood also has solitary pores and lacks tracheids (Miller 1975). Lindackeria has superficial cork cambium. Pollen variation is considerable (Wendt 1988), as is that in ovule development and seed coat anatomy (Dathan & Singh 1979;van Heel 1973 1974, 1977, 1979; Steyn et al. 2002a, b, 2003).
Information is taken from Hegnauer (1966, 1989, as Flacourtiaceae: chemistry), Endress and Voser (1975: floral development of Caloncoba), Spencer and Seigler (1985b: chemistry), Lemke (1988), Gavrilova (1998: pollen), and Groppo et al. (2010: general); see also Judd (1997a) and especially Chase et al. (2002). Bernhard and Endress (1999) discuss androecial initiation.
Phylogeny. For the circumscription of Achariaceae, see Chase et al. (2002) and Sosa et al. (2003): Acharieae (more or less herbaceous and viny; no testal bundles; zig-zag micropyle; fibrous exotegmen), Erythrospermeae (Erythrospermum - fibrous exotegmen), Pangieae (inc. Kiggelarieae) and Lindackerieae (Oncobeae minus Oncoba). Note that the family is divided into three strongly-supported clades, largely Hydnocarpus, Erythrospermeae + Lindackerieae, and Acharieae + Pangieae, and support for monophyly of the family as a whole is strong (Sosa et al. 2003), Groppo et al. (2010) questioned some tribal limits in the family.
Classification. I have not followed the classification in Chase et al. (2002) since the four tribes recognised there do not map (in terms of monophyly) on to the tree.
Previous Relationships. The bulk of Achariaceae had almost universally been included in Flacourtiaceae s.l. until recently (e.g. Cronquist 1981; Takhtajan 1997), most of which = Salicaceae, q.v.
Botanical Trivia. Immature fruits of Australian Ryparosa have the highest concentrations of cyanogenic glucosides known - 12 mg g-1 dry weight (Webber & Woodrow 2004).
Thanks. I thank Sue Zmarzty for comments.
Synonymy: Erythrospermaceae Doweld, Kiggelariaceae Link, nom. inval., Pangiaceae Hasskarl
[[Goupiaceae + Violaceae] [Passifloraceae [Lacistemataceae + Salicaceae]]]: stamens = and opposite sepals; G [3]; exotegmen of ± cuboidal cells.
[Goupiaceae + Violaceae]: petiole with ± annular and wing bundles; cuticle waxes 0; connective ± developed apically; stigma proper ± punctate, receptive area small, appearing recessed/hollow.
GOUPIACEAE Miers Back to Malpighiales
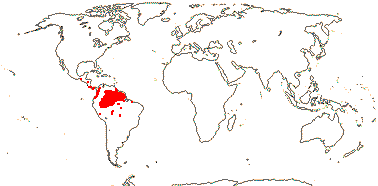
Evergreen trees; plants Al-accumulators, otherwise chemistry unknown; vascular cylinder and pith 4-5-angled; vessel elements with scalariform perforation plates; petiole with inverted medullary bundle; branched sclereids +/0; hairs thick-walled, with pitted bases; leaves two-ranked, lamina tooth ?type, secondary veins actinodromous, 3ary veins scalariform; inflorescences umbellate, axillary, pedicel articulation?; C induplicate-valvate, long, apical part inflexed; nectary annular; connective stout, shortly prolonged, with long hairs; pollen with endexinal folds; G [5], opposite petals, placentation basal-axile, ovary with roof, styluli short, adaxially channeled, on outer shoulders of carpels, stigma type?; ovules few/carpel, ?morphology; fruit a berry; seeds not arillate, testa reticulate, both it and tegmen ca 3 cells across, exotegmen ridged, with 1 layer of sclereids, wall thickenings U-shaped; endosperm copious; n = ?
1/2. Central and N. South America (map: from Tropicos xii.2010).
Evolution. Divergence & Distribution. Diversification in the Goupiaceae clade seems to have slowed down (Xi et al. 2012).
Chemistry, Morphology, etc. The family is poorly known, especially embryologically. It is often suggested that only seedlings have dentate leaves, those of the adult being entire, but leaves of flowering specimens are in fact frequently toothed. () noted that there was a lignified endocarp, so the fruit could technically be called a drupe.
Information on pollen is taken from Lobreau-Callen (1977, 1980) and Furness (2011), on anatomy from den Hartog and Baas (1978), on the seed, from Takhtajan (2000: apparently not entirely accurate), and for general information, inc. anatomy, see Hoyos (2011).
Previous Relationships. Cronquist (1981) included Goupiaceae in Celastraceae, Takhtajan (1997) in Celastrales, A.-L. de Jussieu and others have placed it in Rhamnaceae. Furness (2011: pollen size) suggested that Goupiaceae were closest to the Lacistemataceae-Salicaceae clade.
VIOLACEAE Batsch, nom. cons. Back to Malpighiales
Evergreen trees; vessel elements with simple or scalariform perforation plates; calcium oxalate often as crystals; petiole bundles arcuate; pedicels articulated; flowers weakly monosymmetric; K quincuncial; A with connective forming apical scales; K persistent in fruit; exotesta subpalisade to tabular, ± thickened, (mesotesta sclerenchymatous), endotesta usu. crystalliferous; exotegmen cells tracheidal, lignified, thickened on all walls; x = 8 (?7, ?6), genome size [1C] (0.362-)1.135(-3.562) pg.
23[list]/800. World-wide.
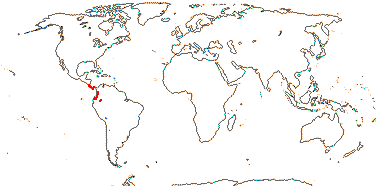
1. Fusispermoideae Hekking
Pith with thin-walled cells; nodes 5:5; petiole with an elliptical medullary bundle, phloem internal; C contorted; disc fleshy, 5-lobed, lobes alternating with A, filaments ± adnate to inner surface at indentations; anther thecae cordate/trapezoid, confluent apically?, connective as short paired fringed apical scales; capsule ca 3 mm long; seeds elongated, longitudinally winged, aril 0, ?exotegmen only moderately developed, of somewhat elongated cells; n = ?
1/3. Costa Rica, Panama, Columbia, Peru (Amazonas) (map: from S. Hoyos, pers. comm.).
2. Violoideae Beilschmied
nectary opposite A; connective lobed, as long as and broader than anther, ± free, margin entire or erose.
2A. ex Rinorea
Nectary as thick semicircular lobes at base of A; anther connective subapical, thecae visible from abaxial side; style broadening subapically; ovule 1/carpel; n = ?
1/3. Costa Rica to Peru.
(Herbs and lianes); plants often Al accumulators; (cylcotide proteins +), tannins 0 [woody members?]; vessel elements with simple or scalariform perforation plates; calcium oxalate often as crystals; (petiole bundles arcuate); stomata also para- or anisocytic; leaves spiral or two-ranked (opposite), lamina margins involute, colleters +, (stipules petiolar; lobed); flowers often strongly monosymmetric, (papilionoid); C quincuncial, abaxial C spurred or not; A (3), basally connate or not, nectariferous appendage on abaxial surface of filaments (on 2 abaxial A only), (filaments connate), anthers connivent, thecae (horizontal), obscured by connective from abaxial side, (connective ± 0); G [(2-5)], (when 5 opposite K), styles separate or style +, straight or curved, apex subcapitate, asymmetric or not, (stigma hollow); ovule (1/carpel), with zig-zag micropyle (endostomal), outer integument 2-4 cells across, inner integument ca 3 cells across, parietal tissue 2-3 cells across, nucellar cap ca 2 cells across, hypostase +; (fruit a berry, nutlike); seeds (winged), (not arillate/carunculate), (exotesta ± thick-walled), (mesotesta sclerenchymatous), endotesta crystalliferous, fibre layer 1-3 cells across, endotegmen crystalliferous; embryo (small), green [Viola]; n = 6(?plesiomorphic)-13+.
22/795: Viola (400-600, Rinorea (160-270), Hybanthus (90-150). World-wide; woody taxa esp. in the lowland tropics (map: from Hultén 1958, 1971; Fl. Austral. ?. 1982; Hultén & Fries 1986; Hekking 1988 - incomplete for South America). [Photo - Leonia, Alexis fruit and flowers, Viola.]
Synonymy: Alsodeiaceae J. Agardh, Leoniaceae A. L. de Candolle
Evolution. Divergence & Distribution. Marcussen et al. (2012) disentangle the complex reticulate history of the polyoploid northern hemisphere species of Viola where hybridization began ca 14 m.y.a.. This group includes the radiation on Hawaii (woody species!), the ancestry of which is to be sought in west North American species. The genus may have had its origin in Andean South America (Ballard et al. 1998).
Plant-Animal Interactions. Violaceae are the preferred hosts of the majority of fritillaries (Nymphalidae: Argynnini - see Simonsen 2006).
Pollination Biology & Seed Dispersal. Cleistogamy is widespread in Viola.
The largely temperate Viola is myrmecochorous (Lengyel et al. 2010), and myrmecochory as well as dispersal by larger animals and by wind also occurs in more tropical taxa.
Genes & Genomes. Viola tricolor, the pansy, and the related V. arvensis were important subjects of early studies of genetics and speciation.
Chemistry, Morphology, etc. Inulin has been reported from Hybanthus (Beauvisage 1889). From the description of the root of Ionidium (= Hybanthus) ipecacuanha by Beauvisage (1889), the cork cambium may be mid-cortical or superficial. Viola has storied cambium.
For information on the flowers of Fusispermum, see Cuatrecasas (1950) and Hekking (1984), the former describes the scales as being ventral appendages of the connective. Feng and Ballard (2005) suggested that even those Violaceae with polysymmetric adult flowers were monosymmetric earlier in development, so "flowers monosymmetric, at least in bud" may be an apomorphy for all/most of the family (see also Arnal 1946 for monosymmetry). The anthers and stigmas of many species are very complex (e.g. Kuta et al. 2012: Viola). In Anchietea and Decorsella the seeds mature exposed on the open carpels.
For seed anatomy, etc., see Singh (1963), Singh and Gupta (1963), and Dathan and Singh (1974), for embryology, etc., see Singh (1970), for chemistry, see Hegnauer (1970, 1990), for cyclotides, see Burman et al. (2010), for information on the basal pectinations in the family, see Hoyos (2011), for general information, see Hekking (1988), Munzinger and Ballard (2003: also key to genera, two undescribed); see also Leins and Erbar (2010: flowers of Viola). An unpublished thesis by Feng (2005) includes a phylogeny of the family and details of the floral development of seven genera.
Phylogeny. There is good support for the relationships [Fusispermum [Rinorea + the rest]], with the old Leonioideae being embedded within Violoideae (Tokuoka 2008, see also Feng & Ballard 2005; Ballard et al. 2009). Bakker et al. (2006b) discuss relationships in African Rinorea.
Classification. The large genus Hybanthus is polphyletic, to be cut up into nine genera, perhaps... (Ballard et al. 2009).
[Passifloraceae [Lacistemataceae + Salicaceae]]: ?
Phylogeny. This clade (= the salicoids") has strong support in the analysis of Xi et al. (2012).
PASSIFLORACEAE Roussel, nom. cons. Back to Malpighiales
Cyclopentenoid cyanogenic glycosides and/or cyclopentenyl fatty acids +, cyanogenic glycosides derived from valine and isoleucine +; (plant with unpleasant smell); leaves spiral, (foliar glands +), colleters +; K + C together forming a tube, corona or scales towards mouth of tube (0); styluli separate; exotestal cells arranged in lines, endotestal cells large, exotegmen sclereidal-palisade, endotegmen persistent; endosperm persistent, oily; x = 7, x = 7 (?6), genome size [1C] (0.035-)1.098(-34.94) pg; biparental or paternal transmission of plastids, atpF intron lost.
27/935. Tropical, esp. America and Africa, also warm temperate - 3 subfamilies below.
1. Malesherbioideae Burnett
Herbaceous or subwoody; hairs conspicuous, multiseriate, often glandular; plant with unpleasant smell; tannins?; (cork cortical); vessel elements usu. with simple perforation plates; nodes also 1:1; lamina often deeply lobed, (margins entire), (stipules foliaceous or 0), colleters?; K + C tube long, (denticulate corona at the mouth), K valvate, subpetal-like, C valvate; androgynophore +; nectary at base; G [3, 4], styles slender, stigmas capitate-clavate, ?type; ovule with large protrusion at chalazal end, micropyle endostomal; K + C tube persistent; seeds pitted, aril 0; endosperm type?
1[list]/24. Andean South America from Peru S., esp. N. Chile (map: see Gengler-Novak 2002). [Photo - Habit]
Synonymy: Malesherbiaceae D. Don, nom. cons.
[Turneroideae + Passifloroideae]: lamina vernation conduplicate, extrafloral nectaries often on petiole/base of lamina; anthers long; tapetum amoeboid.
2. Turneroideae Eaton
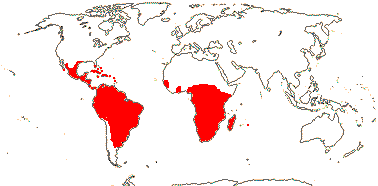
Herbaceous or woody; plant with unpleasant smell; ellagic acid 0; cortical vascular bundles [= leaf traces] common; vessel elements with simple (and scalariform) perforation plates; stomata various; hairs tufted/stellate; colleters +; stipules 0 (+ - e.g. Erblichia); bracteoles often large; (flowers heterostylous); (glands or corona at mouth of K + C tube), C contorted, deliquescent; nectary near base of tube (on sepals; filaments); G [2, 3] (half inferior), stigmas concave, often ± penicillate; micropyle zig-zag, outer integument ca 2 cells across, inner integument c3(-4) cells across, (nucellar cap ca 2 cells across), parietal tissue 2-3 cells across, hypostase +/0; K + C tube deciduous; aril fimbriate; n also = 5 (13).
10[list]/205: Turnera (122), Piriquetia (44). Tropical to warm temperate America and Africa (inc. Madagascar and Rodriguez I.) (map: from Wickens 1976; Heywood 2007, in part; Arbo 2008; Thulin et al. 2012a). [Photo - Flower.]
Synonymy: Piriquetaceae Martynov, Turneraceae Candolle, nom. cons.
3. Passifloroideae Burnett
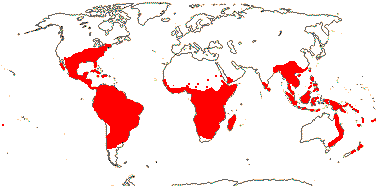
Woody; (lamina margins entire); flowers (3-)5-merous; K ± petal-like, corona +, of (1-)2-several rows of filaments or membranes (0), nectary ± on K/C tube; androgynophore +; (A basally connate), anthers versatile; G [(2-)3(-7)], stigma/s capitate; seeds flattened, surface not smooth, bony; exotesta cells not in lines; endosperm ruminate.
16[list]/705. Tropics to warm temperate, especially Africa and America - two tribes below.
3A. Passifloreae de Candolle
Vines or lianes, climbing by simple branch tendrils; flavonols +, ellagic acid +/0, tannins 0; anomalous secondary thickening quite common; vessel elements with simple perforation plates; wood often fluorescing; supernumerary buds +; leaves (compound), lamina vernation conduplicate, (margins entire), secondary veins often palmate, glands common on petiole or on lamina surface; (plant dioecious), inflorescence cymose; (flowers monosymmetric; (C 0, 1), corona of (1-)2-several rows of filaments or membranes (0), nectary ± on K/C tube, (A basally connate); pollen to 12-colporate; (gynophore +); (G [4]), stigmas capitate or divided [Adenia], with multicellular papillae; ovules with bistomal or zig-zag micropyle, outer integument 2-5 cells across, inner integument 3-5 cells across, nucellus protrudes through micropyle, parietal tissue 6-20 cells across, (nucellar cap +), hypostase +, funicle often long; fruit a berry, (capsule - Passiflora section Xerogona); seeds hairy or not, often sculpted; testa multiplicative, sarcoexotestal, or palisade, endotesta crystalliferous, lignified or not; n = 6 (7) 9(-12); rpl22 gene transferred from chloroplast to nucleus [Passiflora].
10/675: Passiflora (525), Adenia (100). Tropics to warm temperate, especially Africa and America (map: from van Balgooy 1975; Fl. Austral. ?. 1982). [Photo - Collection]
3B. Paropsieae de Candolle
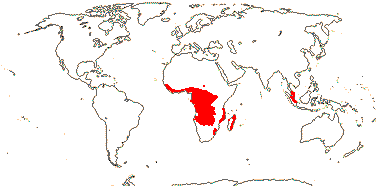
Trees or shrubs; vessel elements in multiples, with scalariform perforation plates; leaves reduced [orthotropic axes], two-ranked [plagiotropic axes], lamina with glands especially on margin and apex, (stipules 0); inflorescence racemose; (androgynophore 0); (A-30, partly connate); (pollen 6-porate); nectary 0 (annular); (gynophore 0); (style single [Barteria]); (fruit dry, indehiscent); seeds scrobiculate; n = ?
6/ca 22: Paropsia 12. Tropical, esp. West Africa (map: from Sleumer 1970; de Vos & Breteler 2009).
Synonymy: Modeccaceae Horaninow, Paropsiaceae Dumortier, Smeathmanniaceae Perleb
Evolution. Divergence & Distribution. Most species of Turneroideae are New World, most genera are African; crown Turneroideae are dated to (42.5-)32.3(-42.5) m.y. (Thulin et al. 2012a). The rate of diversification may have increased in Passifloroideae or [Turneroidae + Passifloroideae] (Xi et al. 2012).
Thulin et al. (2012a) suggest that Mathurina, the only Turneroideae with wind-dispersed seeds, is older than the Rodrigues Island, to which it is now restricted.
Plant-Animal Interactions. The cyclopentenoid glycosides common in Passifloraceae may be sequestered by caterpillars feeding on the plants and perhaps used in defence and/or even as nitrogen sources; Achariaceae also have such glycosides.
Caterpillars of Heliconius butterflies use Passiflora and its relatives as their main food (Fordyce 2010 for references and diversification rates). The plants show great variation in leaf morphology and foliar glands. Some of the glands are egg mimics (Vanderplank 2007 for references) and/or are epithelial extrafloral nectaries (Krosnick et al. 2011 for a summary), etc., indeed, it has been suggested that extrafloral nectaries are a key innovation for Passiflora (Krosnick et al. 2011). Butterflies lay eggs on plants that lack eggs, hence the mimicry of the glands. Heliconiine butterflies may have diversified on the foothills and lower sloped on the eastern Andes from Peru northwards (Rosser et al. 2012). Heliconius itself is also closely associated with Psiguria (Cucurbitaceae) and relatives, and perhaps some other plants, which it pollinates; unusually, the pollen is a source of nutrients for the butterfly. The larvae of some Acraeinae, another nymphalid butterfly, and also of brightly-coloured Notodontidae-Dioptininae (moths) are also often found on Passiflora (Miller 1992; Silva-Brandão et al. 2008), and at least the former are also found on Barteria. Turneroideae are the hosts of caterpillars of several genera of Nymphalidae, alternate hosts include Salicaceae, Passifloroideae, and Violaceae (Arbo 2006 and references).
Details of the association between the African ant-plant Bartera fistulosa and the ant Tetraponera aethiops are given by Dejean et al. (2008).
Vegetative Variation. For the considerable anatomical variation in Adenia as well as variation in life form, see Hearn (2006, 2009a); Hearn (2009b) suggested that the place of development of vascular strands and associated parenchymatous storage tissue in root and/or stem varied spatially in the plant, hence helping to generate the diversity of growth forms in the genus.
Pollination Biology & Seed Dispersal. A little is known about the development of the complex series of fimbriae and membranes at the base of the androgynophore and on the KC tube in Passiflora. In subgenus Passiflora at least, genes normally involved in stamen development are expressed in the centripetally developing fimbriae, although the limen, a rim structure at the base of the androgynophore protecting the nectar, is from this point of view an organ sui generis (Hemingway et al. 2011). Species of Passiflora like P. ampullacea have monosymmetric flowers, with stamens, etc, adaxially positioned, and in at least some of these species the odd sepal is abaxial (Macdougal 1994, they also have four carpels); P. unipetala has but a single petal in the adaxial position. Sazima and Sazima (1978) note that the bat-pollinated flowers of Passiflora mucronata become zygomorphic as the stamens move after the flowers opens (see also Endress and Matthews 2006a).
It has been suggested that there is floral mimicry between Turnera and Malvaceae in Argentina (Benitez-Vieyra et al. 2007). Heterostyly is common in Turnera, Piriqueta and some other genera of Turneroideae.
Myrmecochory occurs in Turnera (Lengyel et al. 2010).
Chemistry, Morphology, etc. Cyanogenic glycosides in this family have a variety of precursors, both protein and non-protein amino acids (Miller et al. 2006 for references); glycosides of individual groups may be distinctive, e.g. Malesherbioideae, which has teraphyllin (Spencer & Siegler 1985a).
Cronquist (1981) suggested that Malesherbioideae lack stipules. In species of Passiflora with strongly bilobed leaves, vernation may be modified conduplicate: The blade makes a V with an inverted V at the end of each arm. The tendril is an axillary shoot and flowers can arise from prophyllar buds on it. For the floral and extrafloral nectaries of Passifloraceae, see Krosnick et al. (2008a, b, 2011). The latter are anatomically quite different from the former (i.a. they lack nectarostomata) and the CRABS CLAW genes is not expressed in them (Krosnick et al. 2008a), and so they are arguably not "homologous".
Species of both Turnera and Piriqueta have epiphyllous flowers. Although the tubular flowers of Passifloraceae s.l. are often described as having a hypanthium, the tube is nearly always formed from calycine and corolline elements only. For androgynophore evolution, see Tokuoka (2012). The styles of Malesherbioideae are shown as being commissural by Schnizlein (1843-1870: fam. 198). For a discussion on aril development, see Kloos and Bouman (1980); although the aril is often described as funicular, they incline to call it raphal. Turneroideae have biparental or paternal transmission of plastids, as may Passifloroideae (Shore et al. 1994).
The old Flacourtiaceae-Paropsieae (Barteria, Paropsia, etc.) are to be included with Passifloroideae (Chase et al. 2002). Anatomically the two are rather similar, indeed, the major variation in the subfamily seems to be associated with habit - lianes versus trees (Ayensu & Stern 1964). Passiflora and its immediate relatives have stem collenchyma, cymose inflorescences, and branches developing from an accessory (superposed) bud, as is common in taxa that have axillary tendrils with non-basal prophyllar buds. In Passifloreae these prophyllar buds may produce additional tendrils, hence branched tendrils, or flowers. Adenia is rather different from other Passifloroideae, perhaps being more like the two other subfamilies, e.g. in having an only moderately developed corona and tricolporate pollen (e.g. see Feuillet & MacDougal 2006). Adenia also thas a nectary often made up of separate glands, a hollow style, and its stigma lacks multicellular papillae (Bernhard 1999a, c), in addition, it may be dioecious, it lacks an androgynophore, its stamens are sometimes connate, it may have a gynophore, and some species have a true hypanthium (de Wilde 1971b).
Do the sieve tubes have non-dispersive protein bodies?
In Passifloreae, carpel orientation is taken from Le Maout and Decaisne (1868) and Schnizlein (1843-1870: fam. 197); for pollen, see Presting (1965) and Spirlet (1965), for chemistry, see Hegnauer (1969, 1990), for embryology, etc., see Raju (1956a) and Singh (1970), for branching, see de Wilde (1971a), for anatomy, see Harms (1893), for stipules, see Dahlgren and van Wyk (1988), and for general information, see de Wilde (1974) and Feuillet and MacDougal (2006). For general information on Paropsieae, see de Vos and Breteler (2009). Some information on Turneroideae is taken from Raju (1956b) and Vijayaraghavan and Kaur (1967), both embryology and seed, Hegnauer (chemistry), González and Arbo (2005: anatomy) and Gonzalez et al. (2012: Aldenoa, general), and Arbo (2006: general account). General information on Malesherbioideae is taken from Ricardo S. (1967, he suggested that the micropyle is endostomal) and Kubitzki (2006b); for chemistry, see Hegnauer (1969, 1990). Embryologically Malesherbioideae are largely unknown.
For floral anatomy of Passiflora, see Puri (1947), for floral morphology, see Endress (1994b), and for floral development, see Krosnick et al. (2006); for a general account of the genus, see Ulmer and MacDougal (2004). Hansen et al. (2006) discuss chromosome number evolution, n = 12 may be the basal number; see also de Melo and Guerra (2003) and Mayrose et al. (2010).
Phylogeny. Turneraceae were weakly associated with Malesherbiaceae in Chase et al. (2002), the two being strongly associated with Passifloraceae. Although preliminary data seemed to suggest that a paraphyletic Passifloraceae might include Turneraceae and Malesherbiaceae (A.P.G. II 2003), Korotkova et al. (2009: only three taxa from the three families) found that Turnera and Passiflora were sister and with 98% jacknife support. For relationships in the clade that are the same as those shown above, see Tokuoka (2012).
Arbo and Espert (2009: morphological analysis, basally pectinate tree with little support) discuss the morphology and biogeography of Turnera; Thulin et al. (2012a) disentangled relationships within Turneroideae, finding three or so links across the South Atlantic and several African genera forming a distinct clade. For relationships within Malesherbioideae, see Gengler-Novak (2002, 2003), and for those within Passifloroideae, see Tokuoka (2012). For a phylogeny of Passiflora, see Yockteng and Nadot (2004), Krosnick and Freudenstein (2005: also morphology), and Krosnick and Freudenstein (2006). See Hearn (2006) for a phylogeny of Adenia.
Classification. Including Turneraceae and Malesherbiaceae in Passifloraceae s.l. is an optional arrangement in A.P.G. II (2003), and given the basic similarity of the three families, they were placed in one by A.P.G. III (2009).
For a revision of Turnera, see Arbo (2008 and references). Passiflora includes Hollrungia and Tetrapathea (Krosnick & Freudenstein 2006); for a formal infrageneric classification of Passiflora, see Feuillet and Macdougal (2004).
Thanks. I am grateful to J. M. MacDougal for information.
[Lacistemataceae + Salicaceae]: anthers ellipsoid to subglobose; endosperm copious.
LACISTEMATACEAE Martius, nom. cons. Back to Malpighiales
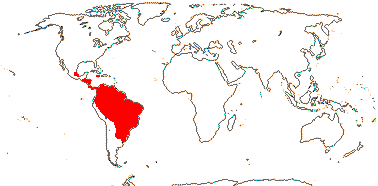
Trees; plants Al accumulators; chemistry?; vessel elements with scalariform perforation plates; sieve tubes?; petiole bundle D or deeply C-shaped, also wing bundles +; leaves two-ranked, (lamina entire); inflorescence a raceme to dense spike; flowers small; P +, cup-like [1-6]; A 1, the thecae ± separated and even stipitate; ?aril; G [2-3], median member adaxial, placentation apical, style branches short, ?stigma; ovules 1-2/carpel, funicles thick, long, ovule type?; fruit a 1(-3)-seeded capsule; testa fleshy or not; embryo (short), with foliaceous cotyledons; n = 22, chromosomes 0.9-2.3 µm long.
2[list]/14. Greater Antilles (Jamaica), Mexico southwards, not in Chile. (map: from Sleumer 1980).[Photo - Flower, Fruit]
Chemistry, Morphology, etc. Chirtoiü (1918) described the flower as having 4-5 free perianth parts and an irregularly lobed disc. The presence of an aril in Lacistemataceae needs to be confirmed (see also Corner 1976). Sleumer (1980) records one for Lacistema, but a fleshy seed coat for Lozania; an aril seems obvious in neither. In the latter genus there appear to be long "hairs" inside the fruit which perhaps support the dangling seed; these hairs are thick-walled but unlignified cells that may be derived from the funicle (see also Casearia, Salicaceae). However, the embryology, etc., of the family are largely unknown.
Additional information from: Lozania - Riviere 270 (anatomy), Gentry et al. 22231 (fruit); Lacistema - Aymard & Delgado 6882 (fruit), Rimachi Y. 11201 (anatomy - stomata tending to anisocytic). See Young (2007 onwards: Lacistemataceae website, focus is on species, nomenclature, etc.
Phylogeny. Lacistemataceae do not cluster with the rest of Salicaceae and Kiggelariaceae (Savolainen et al. 2000a), although they are probably in this area (Chase et al. 2002; see also D. Soltis et al. 1999, 2000). Davis et al. (2005a) place them as sister to Salicaceae s.l. (61% bootstrap, 1.0 posterior probability), as do Korotkova et al. (2009: slightly higher jacknife); as might be expected, they lack salicoid teeth.
Classification. See Sleumer (1980) as Flacourtiaceae: Lacistemeae for a monograph)
SALICACEAE Mirbel, nom. cons. Back to Malpighiales
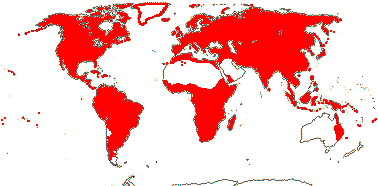
Evergreen (deciduous) trees; cocarcinogens, (gynocardin, ellagic acid +), tanniniferous; cork?; vessel elements with simple or scalariform perforation plates; petiole bundle arcuate or annular with wing bundles; leaves spiral or two-ranked (opposite), lamina vernation supervolute-curved or involute, (margin entire), (venation palmate, (glands +), (stipules 0); inflorescence various; flowers 3-6-merous, (hypanthium +), K (0-)3-8(-15), often valvate, (basally connate; corona +), nectary extrastaminal, often with glands or lobes; A 1 to many (fasciculate, opposite petals), anthers (extrorse), (linear); G [2-5], (placentation axile), styles separate or fused; ovules straight or anatropous, outer and inner integuments 2-5 cells across, micropyle bistomal/zig-zag, parietal tissue 5-7 cells across, nucellar cap + (0), both ± resorbed, funicle short; fruit also a berry (drupe); (embryo green); n = 9, 10-12, 19, x = 10, genome size [1C] (0.018-)0.589(-19.042) pg.
55[list]/1010. Pantropical, also temperate (but few in the Antipodes) to Arctic (map: (Africa incomplete) from Sleumer 1954; Meusel et al. 1975; Sleumer 1980; Hultén & Fries 1986). [Photo - Flower, Fruit.]
1. Samydoideae Reveal
Lamina often punctate or lineate, teeth theoid; hypanthium +; P +, 3-7, basally connate, C 0; nectary on base of P; A 3-many, initiated simultaneously; (seed squeezed from fruit, aril vascularized - Casearia); exotegmen cells laterally flattened, crystalliferous.
13/245: Casearia (180). Pantropical, especially South America.
Synonymy: Bembiciaceae R. C. Keating & Takhtajan, Prockiaceae Bertuch, Samydaceae Ventenat, nom. cons.
[Scyphostegioideae + Salicoideae]: lamina with a small vein proceeding into the tooth, where it expands, the apex of the tooth being a variously coloured spherical gland or stout hair [salicoid teeth].
2. Scyphostegioideae Reveal
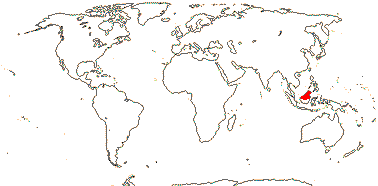
Vessels in radial multiples, with simple (and scalariform) perforations; rays mostly uniseriate; petiole bundle annular and with adaxial mass of xylem and phloem becoming adaxial inverted plate of vascular tissue; stomata paracytic; leaves 2-ranked; plant dioecious, inflorescences terminal, branched, long-lived, bracts large, overlapping, tubular [c.f. Alpinia sect. Myriocrater!], pedicels not articulated; P +, = T 3 + 3, connate; staminate flowers: nectary as lobes opposite A; A 3, opposite inner whorl of T, connate, extrorse; pollen ?tricolpate; carpellate flowers: G [8-13], placentation basal, style 0, stigmas ray-like, with an opening in the middle; ovules with much elongated (exo-?)bistomal micropyle, outer integument 2-3 cells across, lobed, inner integument 3-4 cells across, nucellar cap persistent, funicle well developed; fruit a fleshy capsule with lignified commissural valves; seeds with aril from funicle/outer integument; endosperm slight, perisperm +, very scanty; n = 9.
1/1: Scyphostegia borneensis. Borneo, not the southern part. [Photo - Flower, Leaf, Inflorescence.]
Synonymy: Scyphostegiaceae Hutchinson, nom. cons.
3. Salicoideae Arnott
Benzoylated glycosides, etc. +, cyanogenic glycosides 0 (+ - Banara); (nodes 2:2 - some Azara); petiole often with secondary growth; stomata paracytic; (plant dioecious/monoecious); (C 0; more than K - e.g. some Scolopieae); A initiation centrifugal; (pollen inaperturate - Populus); G [2-5(-13)], (inferior - Homalium); (micropyle exostomal - Idesia; endostomal - Oncoba), (outer integument lobed - Caloncoba); embryo sac elongated, ± protruding into the micropyle, (bisporic, 8-nucleate: Allium-type); (seeds with hairs [of arillate origin]), (testa vascularized, sarcotesta +, endotesta palisade - Oncoba); (endosperm 0 - Salix).
41/765: Salix (450), Homalium (180), Xylosma (85), Scolopia (37), Banara (31). Worldwide, but only Saliceae cold Temperate/Arctic, few Australia, not New Zealand.
Synonymy: Flacourtiaceae Richard, Homaliaceae R. Brown, Poliothyrsidaceae Doweld
Evolution. Divergence & Distribution. Boucher et al. (2003) described Pseudosalix, an Eocene fossil from North America, which is morphologically intermediate between Salix and more florally conventional Salicaceae.
Diversification in Scyphostegioideae seems to have slowed down (Xi et al. 2012: as Scyphostegiaceae).
Pollination Biology. Populus is dioecious and wind-pollinated.
Bacterial/Fungal Associations. Salix and its immediate relatives enter into a variety of associations with fungi, including ectomycorrhiza1 and endomycorrhizal, and there may also be dark septate endophytes (e.g. Van der Heijden 2000; Becerra et al. 2009).
Plant-Animal Interactions. Boeckler et al. (2011) discuss the anti-herbivore properties of the phenolic glycosides characteristic of Salix and its immediate relatives, nevertheless, a number of insects and fungi are associated with these plants and their immediate relatives. Phyllonorycter leaf-mining moths (Lepidoptera: Gracillariidae: Phyllocnistinae) are sometimes found on Populus and Salix (Lopez Vaamonde et al. 2006). Ehrlich and Raven (1964) noted that caterpillars of Atella (Nymphalinae) feed on Flacourtiaceae and Salicaceae, while some Notodontidae moths (Miller 1992); rusts (e.g. Melampsora spp. on Salix, M. idesiae on Idesia - Holm 1979), etc., show similar host patterns.
Some hundreds of species of gall-forming sawflies (Hymenoptera: Tenthredinidae: Nematini) are by far the most common on Salix and Populus among their angiosperm hosts, although they have yet to be recorded from other Salicaceae s.l. (Roininen et al. 2005). Unlike other galls, sawfly-induced galls result largely from stimuli provided by the ovipositing wasp, which may inject fluids into the plant, rather than from the activities of the larvae, so the galls assume their mature forms before the eggs hatch (Redfern 2011).
Genes & Genomes. There is a gene duplication in the common ancestor of Salix and Populus, the salicoid duplication, that has been dated to 65-60 m.y. (Tuskan et al. 2006); it will be interesting to know if other Salicaceae s.l. have it. Populus has lost the PHYC gene (Matthews 2010 and references); again, the phylogenetic extent of this loss is of interest.
The [Salix + Populus] clade is dioecious, and an x/y sex determination system has evolved at least three times (twice in Populus - aspens and poplars) in the clade, maybe within the last 7-6 m.y. - although the whole clade is at least 25 m.y.o. (Geraldes et al. 2015; see also Filatov 2015).
Salix is notorious for extensive interspecific hybridisation .
Chemistry, Morphology, etc. Banara is the only genus reported to have cyanogenic glycosides, but it is well embedded within Salicaceae (Chase et al. 2002). The perforation plates of the tracheary elements are more or less simple and the intervascular pits are small. Xylosma, Flacourtia, etc., have groups of large sclereids in the phloem (Zahur 1959). Xylosma and some Casearia seem to have unilacunar nodes. Leaf traces arise an internode below the leaf they innervate in Hasseltia.
Casearia can have phyllanthoid branching, the orthotropic axes having spirally arranged and reduced leaves while the plagiotropic branches are sylleptic and have fully-expanded and two-ranked leaves. Abatia has opposite leaves with at most very small stipules and marginal glands at the base of the lamina. Salicoid leaf teeth are quite variable, but all have secretory palisade cells over parenchyma and are well supplied by vascular tissue, especially xylem (Wilkinson 2007). Thadeo et al. (2008; see also Thadeo & Meira 2009) discuss the similarity between leaf teeth and foliar nectaries in Salicaceae, in particular the foliar nectaries of Prockia crucis that secrete fructose, glucose, sucrose, etc. Nectaries at the base of the lamina in two species of Populus differed markedly. In one they were persistent, with continuous nectar flow, while in another, large amounts of nectar were produced over a short time and cell death occurred, but new nectaries could be produced (Escalante-Pérez et al. 2012). There are also taxa with pli-nerved leaf blades and foliar glands in Salicaceae - this is a vegetatively rather heterogeneous clade.
The valvate perianth members of Abatia are basally connate and bear many filamentous processes, and the flowers lack a nectary. In those species that have a nectary, its morphology is very variable. It is often made up of a number of lobes, and these may be intrastaminal. It has been claimed that the nectary of Salix represents a member of the perianth, but Alford et al. (2009) suggest that it is like that of other members of the family; there are ± receptacular nectaries associated with the stamens in taxa in the clade sister to [Salix + Populus]. Elongated embryo sacs occur in both Salicaceae and old Flacourtiaceae (Steyn et al. 2005a), indeed, the embryo sac more or less protrudes into the micropyle in Archevaletaia (Maheshwari 1950). Corner (1976) described the micropyle of Scyphostegia as being exostomal. The exotegmen of Dovyalis consists of ribbon-like cells.
Some information is taken from Hegnauer (1973, 1990, also 1966, 1989) and Chai (2009), all as Flacourtiaceae, chemistry, Miller (1975: wood anatomy), Thadeo and Meira (2013: leaf anatomy), Spencer and Seigler (1985b: chemistry), Bernhard and Endress (1999: androecial initiation), Gavrilova (1998: pollen), Steyn et al. (2004, 2005a, b: ovule and seed development, summary in latter paper), and van Heel (1977, 1979: testa anatomy); see also Lemke (1988, Judd (1997a) and especially Chase et al. (2002), general. For Scyphostegia, see Metcalfe (1954: anatomy), van Heel (1967: flowers and fruits), and Hutchinson (1973: various interpretations of the gynoecium).
Phylogeny. It had been observed in the past that Salicaceae s. str. and Flacourtiaceae-Idesieae were very close, despite the catkins of the former - they both have similar, distinctive leaf teeth, phenolic-type compounds such as salicin are found here only, etc. (Miller 1975), and as just noted, rusts and caterpillars, perhaps keying in on chemical characters, show similar distributions (e.g. Meeuse 1975).
Chase et al. (2002) have greatly clarified the phylogenetic situation in the old Salicaceae-Flacourtiaceae area (see also Judd 1997a; Nandi et al. 1998; T. Azuma et al. 2000; Savolainen et al. 2000a), although sampling within tribes still needs to be extended. Casearia, which lacks salicoid leaf teeth and has apetalous flowers with a nectariferous area on the basal-adaxial surface of the perianth tube, is sister to the rest of Salicaceae, although support for this position is weak (Chase et al. 2002, c.f. D. Soltis et al. 1999, 2000). Relationships around Salix are [Microhasseltia, etc. [[Salix + Populus] [Olmediella [Bennettiodendron + Idesia]]]; characters like hairy seeds, sepals deciduous in fruit, loss of corolla, and dioecy are apomorphies at various levels within this clade (Alford et al. 2009). For a phylogeny of Salix, see T. Azuma et al. (2000) and Chen et al. (2010). Trichostephanus (Trichostephaneae) was not assigned to any family (Chase et al. 2002), but in lacking petals and in having a disc at the base of the calyx it is like Casearia (Samydeae). Oncoba is remarkably like other members of the erstwhile Oncobeae (see now Achariaceae - Lindackerieae), but they differ in chemistry, leaf tooth type, and stamen initiation; it is perhaps to be assigned to Flacourtieae.
Leskinen and Alström-Rapaport (1999) discussed the relationships of Salix and its immediate relatives.
Classification. Chase et al. (2002) provide a detailed tribal classification for the clade: Abatieae, Bembicieae, Prockieae (inc. Banareae), Oncobeae (Oncoba only), Homalieae, Saliceae, Samydeae (Casearieae), Scolopieae, and Scyphostegieae. These tribal limits may well have to be adjusted, thus Saliceae will probably have to be expanded, but note Flacourtieae are polyphyletic, etc. The classification is only partly adopted here pending more detailed sampling. Alford (2003) recognised three families for the New World genera previously included in Flacourtiaceae - in addition to Achariaceae, Berberidopsidaceae and Lacistemataceae. Salicaceae have dilated stigmas and sometimes lack sepals, while Flacourtiaceae s. str. have attenuate, lobed, or capitate stigmas and petals present or absent.
Generic limits in the Casearia group neeed attention (Samarakoon et al. 2010).
Previous Relationships. In the Englerian system Salix was often kept with the wind-pollinated Amentiferae, not at all close to Flacourtiaceae, a family that was also recognised at the time and which encompassed the bulk of Salicaceae above and also Achariaceae, etc. See Gilg's (1914) emphatic rejection of Hallier's suggestion that there might be a link between Idesia (Flacourtiaceae) and Salicaceae.
Thanks. I am grateful to S. Zmarzty for comments.
[[Peraceae [Rafflesiaceae + Euphorbiaceae]] [[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]]: fruit a part-septicidal + loculicidal capsule/schizocarp.
Phylogeny. See Xi et al. (2012) for the composition of this clade (= euphorbioids) and relationships within it.
Evolution. Divergence & Distribution. Lee et al. (2011) found that genes involved in oxygen and radical detoxification clustered at this node, but Populus and Bruguiera were the only other members of Malpighiales in their study. On strict parsimony grounds the distinctive fruits of Euphorbiaceae, etc., could be a synapomorphy for the whole clade, and then lost, or acquired twice; in any case, these fruits have been lost within Euphorbiaceae s. str., etc.
[Peraceae [Rafflesiaceae + Euphorbiaceae]]: vessel elements with simple perforation plates; flowers small, imperfect; G [3], styles ± separate; ovule 1/carpel, nucellar cap + [unknown in Peraceae]; fruit with outer layer often separating from the woody layer, valves falling off, central column persistent; seeds large, micropylar caruncle + (0); cotyledons longer and broader than radicle.
Evolution. Divergence & Distribution. This one clade includes the extremes of flower size in angiosperms. Pseudanthia have evolved ca 4 times, and in Euphorbia, for example, a single stamen represents the staminate flower. In Rafflesia, on the other hand, the flower can be up to 1.5 m across (e.g. Barkman et al. 2008).
Chemistry, Morphology, etc. Note that many of the features mentioned above are lost in Rafflesiaceae. The exotegmen there is described as having U-shaped thickenings, and the exotegmen of some Peraceae can also look U-shaped in transverse section (see illustrations in Tokuoka & Tobe 2003).
Phylogeny. Barkman et al. (2004a) sequenced the mitochondrial gene, matR, of Rafflesia and found a strong association with Malpighiales (see below for previous placements of Rafflesiaceae). Although sampling within Malpighiales (only three taxa with parietal placentation were studied) and other rosids was poor, Barkman et al. (2004) noted that the flowers of Rafflesia could be interpreted as having a number of features in common with those of Passifloraceae, including a corona (called a diaphragm by students of Rafflesiaceae), androgynophore, parietal placentation (but this is common in echlorophyllous parasites), etc. As Nickrent et al. (2004a) pointed out, the "homology" of these structures needs careful examination. Davis and Wurdack (2004: two nuclear, one mitochondrial [matR] genes), with considerably more extensive sampling, but also confirming a position in Malpighiales, favoured a position closer to Ochnaceae, Clusiaceae and their relatives, and tenuinucellate ovules are common there, too - however, it is quite common for holoparasitic taxa to lack parietal tissue in their ovules... Many of the analyses carried out by Nickrent et al. (2004a) also suggested a position of Rafflesiaceae in or near Malpighiales. Most recently, Davis et al. (2007), using largely mitochondrial genes, exemplars of all families of Malpighiales, and a good sample of Euphorbiaceae s.l. (including three of the four genera of Peraceae, Chaetocarpus only excluded, Euphorbiaceae-Cheilosioideae also included), located Rafflesiaceae within Euphorbiaceae and with quite good support (see also Wurdack & Davis 2009).
Phylogeny. Merino Sutter and Endress (1995) argue for a rather broadly delimited Euphorbiaceae (inc. both Phyllanthaceae and Putranjivaceae), Huber (1991) for a narrower circumscription, with the biovulate taxa (Phyllanthaceae, Picrodendraceae, Putranjivaceae) being considered to be closer to Linales s. str., while Meeuse (1990) also suggested that the family should be split - into eleven families. There is no molecular evidence yet for a broadly delimited Euphorbiaceae (unless Linaceae et al. are to be included), yet Euphorbiaceae s. str, Phyllanthaceae and Picrodendraceae all have a similar and rather distinctive capsule, etc. (see also Merino Sutter et al. 2006).
Splitting Peroideae from the other Euphorbiaceae s. str. - which actually makes the latter more homogeneous in fruit and testa anatomy - and keeping Rafflesiaceae seems reasonable, there being little enthusiasm for including a Rafflesioideae within an already large Euphorbiaceae.

PERACEAE Klotzsch Back to Malpighiales
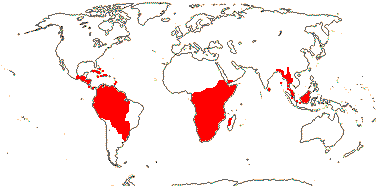
Shrubs to trees; petiole bundles interrupted arcuate to annular, complete annular, (also with medullary plate and wing bundles); stomata?; leaves spiral, lamina vernation involute, margins entire, venation pinnate, stipules (large), small or 0; plant?; (coloured inflorescence bracts +); staminate flowers: C (clawed; 0); nectary lobes opposite K; A 2-8, (?androgynophore, extrorse, opposite C - Clutia); pistillode 0; carpellate flowers: (K 0, C 0); staminodes 0; ovule with both integuments 3-6 cells across; fruit septa membranous and without visible vascularisation, (valves connected at base), perianth [when present] persistent; seeds carunculate or arillate, very shiny; exotesta palisade, lignified, endotesta crystaliferous, exotegmen tracheoidal, oblique, (palisade - Pogonophora); endosperm copious; n = 18, chromosomes 0.5-1.1 µm long.
5/135: Clutia (70), Pera (40). Pantropical, probably not East Malesia (one doubtful report) (map: inaccurate, see van Welzen 1994).
Evolution. Seed Dispersal. Myrmecochory may predominate in this clade (Lengyel et al. 2009).
Chemistry, Morphology, etc. Many details of anatomy, and in particular floral development, ovule morphology, etc., are poorly known.
Neither wood anatomy nor pollen morphology are distinctive (Nowicke et al. 1998; Hayden & Hayden 2000), however, members of Peraceae are variously described as having lysigenous radial canals in the wood, laticiferous cells, and elongated cells with brown contents.
The highly reduced flowers of Pera are surrounded by coloured inflorescence bracts; a pseudanthium of sorts. Pogonophora has adaxially barbellate petals. Style branches are variable in this group, being very short to longer and bifid. The seeds are described as having arils by Tokuoka and Tobe (2003). The anatomy of the seed coat of Peraceae is distinctive compared to that of Euphorbiaceae s. str. - this from an earlier version of /APweb/: "Tokuoka and Tobe (2003) note some Acalyphoideae with a distinctive seed coat anatomy - a more or less tracheoidal exotegmen - unlike that of all other Euphorbiaceae (minus Phyllanthaceae, etc). Interestingly, all the taxa involved belong to other families or are in Euphorbiaceae-Peroideae. These are sister to other Euphorbiaceae, and the distinctive exotegmen structure is more like that common in other Malpighiales. So it is possible that this is plesiomorphic, whereupon the palisade exotegmen of Pogonophora is a parallelism..." The seed coat anatomy of Trigonopleura is unknown.
For pollen, see Nowicke et al. (1998), for wood anatomy, see Hayden and Hayden (2000); for seed coat anatomy, see Huner (1991) and Tokuoka and Tobe (2003); for general information, see Webster (1984) and Radcliffe Smith (2001).
Phylogeny. Tokuoka and Tobe (2006) question the inclusion of Pogonophora in this clade; indeed, if it is to be included and is sister to other Peraceae, then mapping seed coat evolution on the tree does become a bit tricky.
Classification. For a comprehensive checklist and bibliography, see Govaerts et al. (2000, in Euphorbiaceae).
Previous Relationships. Although Airy Shaw (1976) recognised Peraceae as separate from Euphorbiaceae, his Peraceae included only Pera, a genus long considered very distinctive within Euphorbiaceae, even if rarely separated from it. Esser (2003) drew attention to the distinctiveness of the whole group.
[Rafflesiaceae + Euphorbiacaeae]: ?
RAFFLESIACEAE Dumortier, nom. cons. Back to Malpighiales
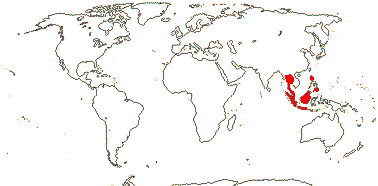
Stem or root parasites, plant endophytic, mycorrhizae, rhizomes and roots 0; vessel elements?; sieve tube plastids lacking starch and protein inclusions; cuticle wax crystalloids 0; plant dioecious [other breeding systems?]; inflorescences various, or flowers single; flowers medium-sized to huge, (perfect); P connate, uniseriate (biseriate in Sapria), 5/10/16-lobed, (valvate - Rhizanthes), with an annular diaphragm , the margins incurved, (absent - Rhizanthes), (nectary on distal part of perianth - Rhizanthes); staminate flowers: A 12-40, adnate to central column [pistillode, = gynostemium], extrorse, anthers sessile, bisporangiate, monothecal to polysporangiate, pores terminal; pollen inaperturate, atectate; carpellate flowers: ovary inferior, nectary at base of style, carpel margins closed by postgenital fusion and secretion, placentation laminar-parietal, loculi irregular, gynostemium short, stigma on outer margin or underside of disc-shaped structure; ovules very many/carpel, parietal tissue 0, inner integument ca 2 cells across, micropyle (exo-)endostomal, nucellar epidermis persists; antipodal cells ephemeral or not; fruit baccate; caruncle 0, seed in two parts, that covered by the testa not enveloping the embryo, exotegmic cells cuboidal, with U thickenings; endosperm slight, embryo undifferentiated; n = 11, 12.
3[list]/20: Rafflesia (16). S. China, Assam, Bhutan, Thailand, W. Malesia (map: from Meijer 1997). [Photo - Flower.]
Evolution. Divergence & Distribution. The age of stem Rafflesiaceae may be (109.5-)95(-83.1) m.y., that of the crown group (95.9-)81.7(-69.5) m.y.a., however, diversification with each genus did not begin until very much later, that in Rafflesia at a mere (15.1-)11.8(-9.2) m.y. being the first - there may have been an extinction event immediately prior to this (Bendiksby et al. 2010, which see for more dates, 95% H.P.D.; see also Barkman et al. 2008). Note that if Sapria split off from other Rafflesiaceae ca 81.7 m.y.a. (Bendiksby et al. 2010) there must have been a major reorganization of the plant body in less than 15 m.y. after Rafflesiaceae split from Euphorbiaceae s. str.
Pollination Biology. The flowers of at least some Rafflesiaceae are thermogenic (Seymour 2001) and pollination, where known, is by flies (Davis et al. 2008). There was a ca 79-fold increase in flower size during the evolution of stem-group Rafflesiaceae over a period of ca 46 m.y., an increase perhaps linked to the adoption of sapropmyophily; size increase in the subsequent ca 60 m.y. was much more modest (Davis et al. 2007, 2008). However, in a more extensive study of Rafflesia, Barkman et al. (2008) suggested that there had been very considerable changes in flower size even within the last 12 m.y. or so, the age of crown group Rafflesia, with repeated both considerable increases and moderate decreases in flower size. Ancestral flower size was (very approximately) 29 cm across (Barkman et al. 2008).
Ecology & Physiology. Rafflesia - Rafflesiaceae in general - are parasitic on species of Tetrastigma (Vitaceae), an association that has evolved more than once (P. Chen et al. 2011a). P. Chen et al. (2011b: HPD) suggest ages of (65.3-)50.6(-36.4) m.y. for stem Tetrastigma, (49.3-)36.9(-25.7) m.y. for the crown group, well before diversification of Rafflesia. However, if the whole family is parasitic on Tetrastigma, there is an interesting timing problem... The vegetative plant body of Rafflesiaceae is endophytic, and Wurdack and Davis (2009) suggested that it might be derived from laticiferous tissue, however, the common ancestor of Euphorbiaceae and Rafflesiaceae are not likely to have had laticifers.
Genes & Genomes. Davis and Wurdack (2004) found that the sequence of another mitochondrial gene (nad1B-c) that they sequenced strongly suggested a relationship between Rafflesiaceae and Vitaceae; the presence of this gene in Rafflesiaceae they reasonably thought was caused by horizontal gene transfer from Vitaceae. Xi et al. (2012a) have since shown that slightly over 2% of the genome may be involved, some Tetrastigma genes functioning in their host, and in a number of vertically transmitted Rafflesia genes, codon usage seemed to be Tetrastigma-like, perhaps facilitating the close association of the host and endophytic parasite - certainly the closest integration of host and parasite genome known in land plants.
Chemistry, Morphology, etc. The plant is tanniniferous (Gottlieb et al. 1989). Although there are stomata in Rafflesia, they are clearly abnormal, having three or more guard cells (Cammerloher 1920).
What the perianth represents is still unclear. If Sapria is interpreted as having a biseriate perianth (the spreading lobes of the flower), then the annular diaphragm in the middle is "coronal" in nature. However, in Rafflesia the tubular structure below the diaphragm seems to have an inner and outer portion, suggesting that the spreading lobes represent the calyx and the diaphragm the connate corolla (D. Boufford, pers. comm.)... Rhizanthes lacks a diaphragm. Furness and Rudall (2004) note a very distinctive combination of microsporogenesis and pollen morphology for the family, but for pollen morphology, see also Blarer et al. (2004). The outer integument, when present, is one cell layer thick, but it is not easy to interpret the ovule (see Solms-Laubach 1874; Ernst & Schmid 1913 for more details). Although the funicle is bent, the integument is not adnate to it; in taxa with a single integument, there is a swelling on the chalaza, perhaps an indication of the other integument. During germination of some Rafflesia, at least, the seed is anchored onto the host by sticky endosperm tubules and also the embryonal primary haustorium, the whole thing looking rather like a T4 bacteriophage (Arekal & Shivamurthy 1976).
For additional information, see Harms (1935a: general), Takhtajan et al. (1985: pollen), Bouman and Meijer (1986: seeds, 1994: ovules and seeds), Meijer (1993: general), Nais (2001: general, superb photographs), the Parasitic Plants website (Nickrent 1998 onwards: general) and also Heide-Jørgensen (2008: general).
Phylogeny. Relationships within Rafflesiaceae are [Sapria [Rhizanthes + Rafflesia]] (Davis et al. 2007).
Previous Relationships. Rafflesiales of some authors included a number of echlorophyllous, parasitic groups including Cytinaceae (here Malvales), Apodanthaceae (Cucurbitales), Mitrastemonaceae (Ericales) and of course Rafflesiaceae itself. Many early and more recent authors have sought an affinity between Rafflesiaceae and taxa like Aristolochiacaeae (references in Takhtajan 1997), perhaps in part because of a belief that the pollen of the former had only a single aperture, as did that of Aristolochiaceae; there is a gynostemium of sorts and extrorse anthers in both. Cocucci and Cocucci (1996) saw connections of Rafflesiaceae first with Apodanthaceae and then with Annonaceae.
Determining the phylogenetic relationships of Rafflesiaceae has been difficult. Apart from the distinctive and often hard-to-interpret morphologies of the families involved, molecular analyses have been problematic in part because of the very long branches in some genes and the general problem of obtaining suitable sequences from holoparasites (e.g. see results from analysing sequences of the mitochondrial atp1 gene - Nickrent et al. 2004a). Indeed, when representatives of all four families are included, an apparently monophyletic Rafflesiales s.l. may be recovered (Nickrent et al. 2004a). Nickrent (2002) had suggested that Rafflesiaceae themselves might be close to Malvales, however, other analyses, including those in which not all members of the erstwhile Rafflesiales were included at one time, suggest a break-up of the group (see also Barkman et al. 2004; Davis & Wurdack 2004; Nickrent et al. 2004a; Davis et al. 2007; Filipowicz & Renner 2010).
EUPHORBIACEAE Jussieu, nom. cons. Back to Malpighiales
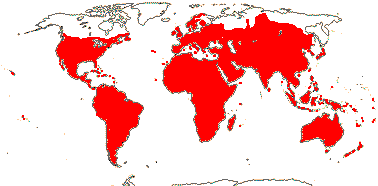
Herbs to trees (lianes), commonly laticiferous; (plants Al-accumulators); cucurbitacins [triterpenes], (polyacetylenes), ellagitannins [geraniin and mallotussic acid], lectins [hemagglutinins], cocarcinogens [phorbol ester diterpenes] +; cork also outer cortical (pericyclic); vessel elements often in multiples, (with scalariform perforation plates); (pits vestured); sieve tubes with non-dispersive protein bodies; (nodes also 1:1 [?Actinostemon] or 5 or more:5 or more); petiole anatomy very variable, often ± annular, etc.; (epidermis silicified); stomata various; leaves spiral, two-ranked or opposite, (palmately compound), lamina vernation variable, margins entire or single veins running into opaque persistent tooth, secondary veins palmate (pinnate), subbasal glands +/0, petioles often apically pulvinate, stipules (0), with axillary colleters; plant monoecious (dioecious); P +, (2-)3-6(-12), (connate), C ?0; nectary disc-like; staminate flowers: A 1-many, (connate), extrorse or introrse; pistillode 0; carpellate flowers: staminodes 0; G [(2) 3(-many)], (alt. with 3 P - Excoecaria), median member usu. abaxial, style (short), branched or not, stigmas prominent, often branched or with adaxial furrow, dry or wet; micropyle exo(bi)stomal, nucellus often projecting beyond micropyle, placental obturator +; (fruit indehiscent; aril +); (seed pachychalazal), exotegmen sclereids laterally flattened, oblique [Malpighian cells]; endosperm + (0), embryo green or white; n = (5-)9(-11+), x = 11 (?10), genome size [1C] (0.045-)1.064(-25.072) pg.
218[list]/5735 - four groups below. Pantropical, also (cool) temperate (map: see Meusel et al. 1978, Canada not very accurate). [Photo - Flower, Flower, Fruit, Fruit.]
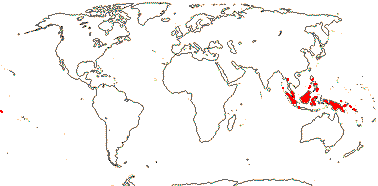
1. Cheilosoideae K. Wurdack & Petra Hoffmann
A 5-12, pollen echinate, (G [2]), outer integument 8-10 cells across, inner integument 8-12 cells across; seeds not carunculate; testa with vascular bundles; endosperm +.
2/7. Burma, Malesia (map: from van Welzen 1994).
Synonymy: Cheilosaceae Doweld
[Acalyphoideae [Crotonoideae + Euphorbioideae]]: (phorbol esters [diterpenes] +); outer integument 6-10 cells or so across.
2. Acalyphoideae Beilschmied
(Tapetum amoeboid); (pollen trinucleate); outer integument 3-6(-16) cells across, inner integument 3-24 cells across, hypostase +; (embryo sac tetrasporic, 12-16 celled - Penaea type); caruncle ?, (testa with vascular bundles); (endosperm 5-10-ploid).
Acalypha (430: stigmas much branched), Macaranga (240), Tragia (170), Mallotus (140), Dalechampia (115), Claoxylon (80). Pantropical.
Synonymy: Acalyphaceae Menge, Mercurialaceae Berchtold & J. Presl, Trewiaceae Lindley.
[Crotonoideae + Euphorbioideae]: laticifers +; (pollen trinucleate - ?level).
3. Crotonoideae Beilschmied
(Herbs); cyanogenesis via the valine/isoleucine pathway; laticifers articulated or not; hairs often stellate or lepidote; (lamina abaxially with paired pale glands near the petiole junction); petals +/0; pollen inaperturate, with supractectal processes attached to muri with short and irregular columellae [Croton-type pollen], or colpate, or porate; (micropyle endostomal), outer integument 4-8 cells across, inner integument (8-)18-25 cells across; micropylar megaspore functional; seeds carunculate/arillate or not, often pachychalazal; (exotesta palisade, endotestal cells ± palisade, thin-walled, slightly lignified), tegmen usu. with vascular bundles; (100+ bp deletion in trnL-F spacer).
Croton (1300+), Jatropha (175), Manihot (100), Trigonostemon (95), Cnidosculus (75). Pantropical, fewer temperate.
Synonymy: Crotonaceae J. Agardh
4. Euphorbioideae Beilschmied
(Herbs); laticifers not articulated, (starch grains much elaborated - Euphorbieae); (inflorescences pseudanthia; P 0 - Euphorbieae); staminate flowers: A not covered by P; (pollen trinucleate); nectary usu. 0; outer integument 3-6 or 8-22 cells across, inner integument 3-7(-22) cells across, parietal tissue 5-6 cells across, (postament +); seeds carunculate or not; (testa mucilaginous), (two layers of tegmic sclereids, fibres between).
Euphorbia (2420), Mabea (40). Pantropical, extending (mostly Euphorbia) into temperate regions.
Synonymy: Bertyaceae J. Agardh, Hippomanaceae J. Agardh, Ricinaceae Martynov, Ricinocarpaceae Hurusawa, Tithymalaceae Ventenat, Tragiaceae Rafinesque
Evolution. Divergence & Distribution. Euphorbiaceae may have diverged from other Malpighiales in the Cretaceous-late Aptian ([119.4-]113.8[-110.7]/[105.9-]101.6[-101.1] m.y.: see Davis et al. 2005a). An estimate for the crown group age of the family is ca 102 m.y. (van Ee et al. 2008).
Diversification within Acalyphoideae occurred within the last ca 70 m.y. (Davis et al. 2005a), while the stem and crown group ages for Croton are ca 55 and ca 40 m.y. respectively (van Ee et al. 2008). A Mallotus-Macaranga-like plant has been found in deposits from New Zealand that are about 23 m.y. old - this clade is not currently known from the islands (Lee et al. 2010).
Divergence within Euphorbieae may have begun (63.5-)48.9(-40.5) m.y.a. (Bruyns et al. 2011); Euphorbia itself started diverging (47.2-)36.6(-29.0) m.y.a., or earlier than 42.5 m.y.a. (van Ee et al. 2008: subgenus Esula not included.) There seems to be but a single origin of the distinctive cyathium here (Park & Backlund 2002; Wurdack et al. 2005), and although the cyathium characterises a very species-rich clade, diversification may also be associated with the evolution of a variety of distinctive life forms and seed dispersal types and the adoption of C4 photosynthesis (see below).
Plant-Animal Interactions. Caterpillars of nymphalid butterflies are quite common on Euphorbiaceae (Ehrlich & Raven 1964), e.g. caterpillars of the ca 340 species of Biblidinae are found on Dalechampia and Tragia (DeVries 1987; Wahlberg et al. 2009). Caterpillars of the spectacular Uraniinae moths can be found on Endospermum, Omphalea and Suregada throughout the tropics (Lees & Smith 1991); the first two are rather closely related, the position of the last is unclear (Wurdack et al. 2005). It would be interesting in this context to clarify both Euphorbiaceae phylogeny and Uraniinae host plant preferences.
In Malesia about 29 species of fast-growing, large-leaved Macaranga are the ecological analogues of the New World Cecropia (Urticaceae); for a phylogeny, see Bänfer et al. (2006). Food bodies (Beccarian bodies) and extra-floral nectaries provide food for the ants (Crematogaster spp.), which live in obligate association with the plants in their hollow stems which are either hollowed out by the ant or become hollow as the stem ages; mymecophytism seems to have evolved more than once here (Hatada et al. 2001 for references; Feldhaar 2003a, b). The association with the ants is some 20-16 m.y. old. Coccus scale insects provide carbohydrates for the ants, but that association is only 9-7 m.y. old (Ueda et al. 2008). Other members of this association include Arhopala (a lycaenid) caterpillars which eat ant larvae but are not attacked by the ants, rather, the products of a tentacle-like gland that is extruded by the caterpillar calm the ants (Maschwitz et al. 1984).
Bacterial/Fungal Associations. Complex maytansinoids, ansamycin antibiotics, that are likely to be synthesized by fungal endophytes or other plant associates, are found in Trewia (Cassady et al. 2004 for references). The fungus Uromyces pisi causes Euphorbia cyparissias to form nectar-producing pseudoflowers that facilitate transmission of the fungus spermatia (Pfunder & Roy 2000).
Pollination Biology & Seed Dispersal. Flowers of Euphorbiaceae are generally small, and pseudanthia have originated more than once (and also in Peraceae). Euphorbia, with some 2,400+ species, is by far the biggest clade with cyathia. The distinctive nectar-secreting glands on the bracts subtending the cyathium may be modified commissural stipules (Steinmann & Porter 2002), while the cyathium itself seems to be a modified cymose inflorescence with a single, terminal, carpellate flower (c.f. Jatropha, etc.); details of its development are provided by Prenner and Rudall (2007), although they found that the morphological nature of both cyathial glands and the petal-like bracts surrounding the cyathium was unclear (see also Hoppe 1985 and Prenner et al. 2008b). Indeed, at the level of gene expression, "floral" genes may be expressed in the cyathium as a whole (Prenner et al. 2011).
The ca 115 species of Dalechampia (Acalyphoideae) also have remarkable pseudanthia. Here female megachilid, euglossine and stingless Trigona (meliponine) bees visit the "flowers" for resin they use to build their nests (resin is a very uncommon reward in flowering plants, see Armbruster 1984, 1996, 2012 for details and references). The resin has secondarily become used for defence in some species, probably its original fuction (Armbruster et al. 2009).
The evolution of the distinctive red, spurred, monosymmetric cyathia in New World species of Euphorbia that used to be segregated as Pedilanthus has been studied from the point of view of whether or not they represent a "key innovation" (Cacho et al. 2010): Bird pollination occurs here. The jury is out, but there aren't many species in this clade... Pollination by thrips (Thysanoptera) seems to be particularly common in myrmecophytic species of Macaranga (Fiala et al. 2011); 24/29 species may be so pollinated, based on floral morphology, about double the frequency when compared with non-myrmecophytic species of the genus growing in the West Malesian localities that Fiala et al. visited.
Seed dispersal is initially usually by the explosion of the capsule, the seed being hurled quite some distance - to 45 m in Hura crepitans, the sand-box tree (Swaine & Beer 1976). The seeds especially of Euphorbia may have nutritive arils or caruncles (e.g. Rössler 1943) which facilitate further local dispersal of the seeds, especially by ants; a total of ca 2300 species in the family, many in Euphorbia, are likely to be myrmecochorous (Lengyel et al. 2010), and caruncles may have evolved ca 13 times there (Horn et al. 2012). In Euphorbia subgenus Chamaesyce section Anisophyllum in particular a number of species have testas that become mucilaginous when wetted (Jordan & Hayden 1992).
Ecology & Physiology. "Euphorbiaceae", i.e. including Phyllanthaceae, Putranjivaceae, etc., are often the second most abundant family in tropical rainforests in South-East Asia and Africa (Gentry 1988).
Both growth patterns and carbon fixation pathways show much diversity, particularly in Euphorbia. Basic relationships here are [Esula [Rhizanthium [Euphorbia + Chamaesyce]]] (subgeneric names, see Bruyns et al. 2011, also below). Diversification is estimated to have begun ca 42.5 m.y.a. (van Ee et al. 2008: probably earlier, subgenus Esula not included; Horn et al. 2012), but other estimates are (47.2-)36.6(-29.0) m.y.a. (Bruyns et al,. 2011). Divergence within the largely succulent subgenera Rhizanthium and Euphorbia began somewhat less than 30 m.y.a., (38.2-(28.1(-22) and (39.6-)29.9(-22.5) m.y. respectively, with much speciation in the latter subgenuus in particular occuring within the last ca 13 m.y. (Bruyns et al. 2011). Euphorbia s. str. (i.e. not including Chamaesyce, etc.) alone is extremely variable (Keller 1996), but with the inclusion of Chamaesyce and other segregate genera (e.g. Horn et al. 2009a, 2012) there is yet more diversity.
Both the cactiform life form and the annual habit, the latter in Euphorbia subgenera Chamaesyce and Esula, seem to be associated with much speciation (Horn et al. 2009a, 2010b); the annual habit has evolved eight times or more in subgenus Esula alone (Frajman & Schönswetter 2011). Stem succulence in Euphorbia - some are quite massive stem succulents - is associated with the evolution of a monopodial growth form and axillary inflorescences (Horn et al. 2012); these plants are cactus-analogues of drier areas throughout Africa and into India and even beyond (Steinmann & Porter 2002; Bruyns et al. 2006, 2011). Succulent Euphorbia is a particularly prominent component of the winter rainfall vegetation of the Succulent Karoo of south west Africa (Nyffeler & Eggli 2010b). All told, some 850 species belonging to all four subgenera are succulents; xeromorphism of one form or another has evolved ca 14 times (Horn et al. 2012), but subgenera Rhizanthium and Euphorbia are made up mostly of succulent species, and they lack any annuals (Bruyns et al. 2011; Morawetz & Riina 2011). Succulence is of various types. The plants may have variously articulated or simply pencil-like stems, the latter as in some species of subgenus Chamaesyce, or they may be medusoid, with relatively slender but succulent branches radiating from a stout central axis, and they may be spiny (in subgenus Euphorbia there are spine shields; there are also stipular spines, as well as spines in the stipular position that are not actually vascularized) or thorny (see e.g. Park & Jansen 2007; Carter 2002, esp. illustrations; Bruyns 2010). Bruyns et al. (2011) describe the different forms of stem succulence found in the genus in detail, and they note that in a number of species of subgenus Euphorbia the branches become permanently differentiated; orthotropic axes will not develop from branches.
Given the prevalence of succulence in Euphorbia, it is not surprising that crassulacean acid metabolism is quite common in the genus. It has evolved ten times or so, especially notable cases being in subgenera Rhizanthium and Euphorbia, the two subgenera in which the succulent habit is particularly common (Horn et al. 2010b; Bruyns et al. 2011).
Most of the ca 350 species of subgenus Chamaesyce section Anisophyllum carry out C4 photosynthesis, a syndrome which probably originated a single time there (Yang & Berry 2011a; Yang et al. 2012). Maximum estimates of the age of origin of the pathway are (13.1-)10.4(-7.3) m.y. (Christin et al. 2011b). Some C4 species grow in the more arid parts of Africa and are also succulents. Horn et al. (2012) emphasize that it is not simply the evolution of the C4 pathway, but also the adoption of a plagiotropic branching habit, etc., that may have made this clade successful, as well as its movement from the Old to the New World (see also Yang & Berry 2011b). Species of section Anisophyllum growing on Hawaii form a distinctive woody radiation and include some of the largest C4 plants anywhere, being trees up to 9 m tall growing in mesic forests (Pearcy & Troughton 1975; see Winter 1981 for slightly larger C4 Chenopodioideae); Horn et al. (2012) discuss how the tree habit might evolve in a clade in which all branches are basically plagiotropic. This Hawaiian clade appears to have evolved from within a small clade of weedy annuals now found in Southern USA, Mexico, and the Caribbean (see also Yang et al. 2009); hybridisation is involved in their origin, and it is also known from elsewhere in the C4 clade (Yang & Berry 2011b). The whole plant body of some species in section Anisophyllum can be compared with the inflorescence of other species of Euphorbia; the seedling apex aborts after the production of a single pair of leaves, and several axillary shoots showing complex determinate branching produce the adult plant (Hayden 1988). There are many different starch grain morphologies in the latex of species in this clade (Biesboer & Mahlberg 1981).
The C4 clade probably evolved in drier areas of North America, and it is sister to the small subsection Acutae from southwest North America in which there are C4, C3, and C2 species, although only species with the latter two kinds of photoynthesis have been sequenced (Yang & Berry 2011a, esp. b; Yang et al. 2012). Sage et al. (2011) described C2 photosynthesis in subsection Acutae, and they noted that E. angusta, with C3 photosynthesis, had some anatomical similarities with C2 and C4 taxa; it is in a small clade from southwest North America sister to the other C4 species.
Chemistry, Morphology, etc. There is great diversity in phorbol esters in the [Crotonoideae + Euphorbioideae] clade. Cyanogens in Euphorbiaceae s. str. can be derived from nicotinic acid or valine/isoleucine (Seigler 1994); latex and cocarcinogens are both apparently restricted to Euphorbioideae and Crotonoideae. Distinctive fatty acids in the seed oils are quite common in the family (Badami & Patil 1981); for lectins, see Vandenborre et al. (2011).
Laticifers in Euphorbiaceae are discussed by Rudall (1987, 1994a) and Wiedenhoft et al. (2009); see also the cautionary comments in Wurdack et al. (2005). Biesboer and Mahlberg (1981) describe the complex morphology of the starch grains found in the latex of Euphorbia and laticifer evolution there. Claoxylon has distinctively rough leaves when dry because of the styloids in their tissues (Kabouw et al. 2008). Prismatic crystals in wood parenchyma and/or ray cells are common, and these also occur in Putranjivaceae and Picrodendraceae (Hayden 1994).
Prenner et al. (2008a) described the development of the distinctive androecium of Ricinus with its branched stamens which, however, are not cauline as has been suggested. Some Euphorbiaceae are reported to have two vascular traces supplying each ovule (Venkata Rao & Ramalkshmi 1967). Johri and Kapil (1953) describe the vascular tissue in Acalypha indica as proceeding one third the way up the nucellus. The projecting nucellus in Codiaeum variegatum, at least, seems to result from periclinal divisions of the epidermal layer of the nucellus, the nucellar cap, although in Euphorbia less striking projections seem to be the result of divisions of the underlying nucellar cells (Bor & Kapil 1975 and references). Mennega (1990) suggested that the subdermal initiation of the inner integument separated Euphorbiaceae from other families.
Van Welzen (1994) described Neoscortechinia (Cheilosoideae) as having a thin, red aril, but no aril was mentioned by Tokuoka and Tobe (2003) or Tokuoka (2007).
For general information on Euphorbiaceae, see Webster (1967, 1994a, b - also other papers in Ann. Missouri Bot. Gard. 81. 1994), Hegnauer (1966, 1989: chemistry), Hans (1973: chromosomes), Evans and Taylor (1983: phorbol esters), Jury et al. (1987: chemistry), Beutler et al. (1989, 1996: chemistry), Kapil (1960: embryology), Tokuoka and Tobe (1993: general embryology, 1998: ovules and seeds in Crotonoideae, 2002: ovules and seeds in Euphorbioideae, 2003: ovules and seeds in Acalyphoideae), Bor & Bouman (1975: ovules and seeds in Euphorbioideae), Pammel (1892: seeds in Euphorbia), Merino Sutter and Endress (1995: floral morphology), De-Paula and Sajo (2011: anthers and ovules in Croton), Singh (1969), and Stuppy (1996), all seed anatomy, Hayden and Hayden (2000: wood anatomy of Acalyphoideae), Radcliffe-Smith and Esser (2001: morphology of genera), Esser (2001: general), Westra and Koek-Noorman (2004: wood end-grain), Mennega (2005: wood anatomy of Euphorbioideae), Tokuoka (2007: character evolution), Cervantes et al. (2009: leaf anatomy of some Acalyphoideae), and Fiser Pecnikar et al. (2012: leaf anatomy of Mallotus and relatives). For pollen morphology of Crotonoideae in particular, see Lobreau-Callen et al. (2000), for that of Acalyphoideae s.l., see Nowicke and Takahashi (2002) and references, for that of Acalypha, see Sagun et al. (2006), and for that of Plukenetieae and Euphorbieae, see Suárez-Cerbera et al. (2001).
Phylogeny. Molecular analyses by Wurdack and Chase (2002) and especially Wurdack et al. (2005, see also Tokuoka 2007) suggest that substantial changes may be needed in the groupings currently recognised in the family. The beginning of the reclassification they suggest is given here, with the interpolation of Rafflesiaceae as suggested by Davis et al. (2007) and the associated recognition of Peraceae; the main problem remaining is the circumscription of Crotonoideae.
Although there are number of distinctive features in the Crotonoideae as broadly construed, there is as yet no evidence that it is monophyletic (see the C1-5 clades in the tree, the C1-2 clades are the same as in Wurdack et al. 2005), and many of the characters in the subfamilial characterization above are synapomorphies either for individual clades or groups within them (the latter the trnL-F spacer deletion - a feature of part of Crotonoideae s. str.). Crotonoideae s. str., the C1-2 clades, may expand from this minimalist circumscription as details of phylogenetic relationships are clarified, and for the most part they have petals, distinctive inaperturate pollen with supratectal processes, and the tegmen is usually vascularized. The pollen of Suregada (Crotonoideae: C1 clade) is pantoporate and it also lacks columellae.
The large genus Croton is being actively studied by Berry and collaborators (see Berry et al. 2005; van Ee et al. 2008, 2011; van Ee & Berry 2009; Riina et al. 2009, 2010; Caruzo et al. 2011 for phylogenies, dates, biogeography, and more). Croton and immediate relatives can be quite readily recognised: Some leaves turn orange with age, the indumentum is stellate-lepidote, there are distinctive often yellowish glands on the petiole, and the stamens are inflexed in bud.
Although Acalyphoideae in the old sense is paraphyletic, the great bulk of the subfamily is included in a single, strongly-supported clade, Acalyphoideae s. str. The plesiomorphic thickness of the outer integument may be 6-10 or so cells across, but Tokuoka (2007) noted that Adenoclineae and Gelonieae have a thinner outer integument, and they may form a paraphyletic grade at the base of Acalyphoideae, albeit currently this topology has very little support. Slik et al. (2001: morphology), Sierra et al. (2006, 2010 and references [the latter with symmetric resampling values and both qualitative and quantitative variation used]) and Kulju et al. (2007a, b) evaluate the phylogeny of the Macaranga-Mallotus complex; there are three main clades, and in Mallotus s. str. in particular some small, segregate genera are embedded.
For a phylogeny of Euphorbia, see Molero et al. (2002: Macaronesian taxa), Bruyns et al. (2006), Park and Jansen (2007), Zimmermann et al. (2010) and Wurdack et al. (2011). The last three authors found that subgenus Esula was sister to The Rest, although not always with very strong support - relationships are [Esula [Rhizanthium [Euphorbia + Chamaesyce]]] (see also the extensive study by Bruyns et al. 2011; Horn et al. 2012). Sampling was initially poor, however, this has steadily improved. Some characters are particularly common/important within the subgenera, although there is very extensive homoplasy. A few of the major characters are [Esula (annuals, cyathium with 4 glands) [Rhizanthium (cyathium with 5 glands, inflorescences sometimes lateral, plants succulents) [Euphorbia (inflorescences often lateral, plants succulents) + Chamaesyce (annuals, C4 photosynthesis, leaves often opposite)]]] (Horn et al. 2012, q.v. for references and many more details).
For relationships in Euphorbia subgenus Esula in particular, see Frajman and Schönswetter (2011: position of E. lathyris unclear; also summary in Geltman et al. 2011), and for those in subgenus Rhizanthium, in which the only Malagasy species, E. antso, is sister to the rest, see Morawetz and Riina (2011). For relationships in the mostly New World subgenus Chamaesyce, see Yang and Berry (2007, 2011) and Yang et al. (2012), and for those in subgenus Euphorbia, see Dorsey et al. (2011: also growth form evolution). See also above for cyathia, succulence, thorns, and photosynthetic pathways.
Classification. For a comprehensive checklist and bibliography of the family, now dated, see Govaerts et al. (2000), while Radcliffe-Smith and Esser (2001) describe the genera. For an outline of the classification of the speciose Macaranga, seee Whitmore (2008). For information on Euphorbia, see the developing site EuphORBia (Esser et al. 2009) and there is an inventory, etc., of the genus, at Tolkin (Riina & Berry 2012), but not if you use Internet Explorer only... Euphorbia is best broadly circumscribed, so including the whole of the Euphorbiinae of Webster (1994b), i.e. genera like Chamaesyce, Pedilanthus, Monadenium, Synadenium, etc., and is well characterized by the possession of a cyathium (e.g. Bruyns 2010 and references); Yang et al. (2012) provide a classification of Euphorbia subgenus Chamaesyce. For generic limits in the Macaranga-Mallotus area, see Kulju et al. (2007a) and Sierra et al. (2007), and for a sectional classification of Croton, see van Ee et al. (2011).
Previous Relationships. Cronquist's Euphorbiales included Simmondsiaceae (Caryophyllales here), Pandaceae (Malpighiales) and Buxaceae (Buxales), whilst Takhtajan's (1997) Euphorbianae included Pandaceae and Dichapetalaceae (Malpighiales), as well as Thymelaeaceae (Malvales) and Aextoxicaceae (Berberidopsidales), but these are clearly groups that have little to recommend them.
Botanical Trivia. Croton has got nothing to do with the cultivated croton, which is Codiaeum.
Thanks. I am grateful to Hajo Esser for comments.
[[Phyllanthaceae + Picrodendraceae] [Ixonanthaceae + Linaceae]]: stomata paracytic; ovules 2/carpel.
[Phyllanthaceae + Picrodendraceae]: plant monoecious; flowers small; style +; micropyle bistomal, parietal tissue 10 or more cells across, obturator +, nucellar beak +; fruit with outer layer often separating from the woody layer, valves falling off, central column persistent; x = 13.
Evolution. Divergence & Distribution. Phyllanthaceae and Picrodendraceae (= phyllanthoids: Xi et al. 2012) may have diverged in the Cretaceous-Albian 111-100 m.y.a., or a little later at (114.0-)108.1(-105.8)/(101.9-)97.1(-95.6) m.y. (Davis et al. 2005a).
Merino Sutter et al. (2006) suggest additional possible similarities between the two families.
Chemistry, Morphology, etc. For chemistry, see Hegnauer (1966, 1989, as Euphorbiaceae) and Jury et al. (1987), see also Webster (1994a, b: general), Radcliffe-Smith and Esser (2001: description of genera), and Wurdack et al. (2004: phylogeny and morphology).
Phylogeny. The clade [Phyllanthaceae + Picrodendraceae] had only very slight support in a rbcL analysis, although the two families have morphological similarities (Wurdack et al. 2004). Support is stronger (75% bootstrap, 1.0 posterior probablility) in a recent 4-gene analysis (Davis et al. 2005a; see also Tokuoka & Tobe 2006; Korotkova et al. 2009) and even stronger in Soltis et al. (2011). Thus the two are probably sister taxa.
PHYLLANTHACEAE Martynov Back to Malpighiales
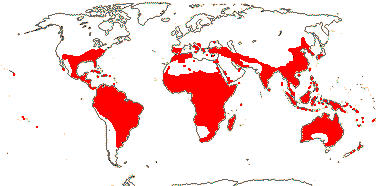
Herbs to trees; (plants Al accumulators); cyanogenesis via the tyrosine pathway, tropane, piperidine and pyrrolizidine alkaloids, cucurbitacins [triterpenes], nonhydrolysable tannins [geraniin] +, ellagic acid 0; cork?; vessel elements with simple [Glochidion] or scalariform [Aporosa] perforation plates; (axial parenchyma 0); (nodes 1:1); (mucilage cells [epidermis] +); (stomata anisocytic); leaves on orthotropic axes often reduced, spiral, on plagiotropic axes two-ranked (spiral), lamina vernation involute or conduplicate, margins entire (toothed); (plant dioecious); K 2-8(-12), often basally connate, C (0, 3-)5(-9), (small); nectary extrastaminal, variously lobed, (0, central); staminate flowers: A 2-35, often ± connate, anthers extrorse; pollen reticulate; (pistillode +); carpellate flowers: (staminodes +); G 1 [2-5(-15)], styles usu. bifid, stigmas with adaxial furrow, wet; ovules with outer integument 2-many cells across, inner integument 2-3(-10) cells across, parietal tissue protruding through micropyle, placental obturator +; fruit a septicidal capsule/schizocarp; seeds large, (caruncle +); (vascular bundles in testa), tegmen 2-5(-20) cells thick, exotegmen with (radially-elongated) ribbon-like cells; endosperm copious (0); n = (6-9, 11, 14).
59[list]/1745. Pantropical, but esp. Malesia, some temperate (map: from Webster 1970, 1984, 1994a, etc.; FloraBase 1.2011 in part - not yet v. accurate...). Two subfamilies below. [Photo - Flower.]
1. Phyllanthoideae Beilschmied
Growth continuous; (plant dioecious); inflorescence fasciculate; (pollen to 16-colporate; colpi diploporate); G [2-6(-15)]; (ovules hemitropous); (fruit indehiscent).
38/ : Phyllanthus (1270), Cleistanthus (140), Bridelia (50). Tropical to Temperate.
Synonymy: Porantheraceae Hurusawa
2. Antidesmatoideae Hurusawa
Growth rhythmic; plant tanniniferous; (pits vestured - Bridelieae); (sieve tubes with non-dispersive protein bodies - Bischofia); (petiole pulvinate; leaves trifoliolate, spiral, teeth deciduous - Bischofia); plant often dioecious; C often 0; G [1-5], (stigma plumose); fruit often indehiscent.
21/ : Antidesma (100), Aporosa (90), Uapaca (60), Baccaurea (50). Tropics and subtropics.
Synonymy: Antidesmataceae Loudon, Aporosaceae Planchon, Bischofiaceae Airy Shaw (tapetal cells 2-5-nuclear), Hymenocardiaceae Airy Shaw, Scepaceae Lindley, Stilaginaceae C. Agardh, Uapacaceae Airy Shaw
Evolution. Divergence & Distribution. Lachnostylis, a small Cape genus, seems to be a relict element there, being dated to as much as 97 m.y.a. (Warren & Hawkins 2006), while Haegens (2000) discusses the distribution of the Baccaurea group as being initially the result of drift events occuring ca 80 m.y.a.. These ages should be re-examined. For the possible (post-)Miocene E->W dispersal of Bridelia across the Indian ocean, see Li et al. (2009).
Bacterial/Fungal Associations. Species of the African-Madagascan Uapaca are ectomycorrhizal (Ducousso et al. 2008) and can locally dominate the vegetation in Madagascar; on Africa they are often a component of Detarieae-dominated woodlands and savannas (White 1983). Ectomycorrhizae sometimes occur along with arbuscular mycorrhizae (Ba et al. 2012).
Pollination Biology. In a clade of some 500+ species of Breynia, Phyllanthus and Glochidion pollination mutualisms involve the moth genus Epicephala (Gracillariidae: Kato et al. 2003; Kawakita & Kato 2004a, b, 2006; Kawakita et al. 2004; Svensson et al. 2010); the plants involved are all part of Phyllanthus s.l. (see below). Flowers in which the moth lays eggs lose all their seeds to the growing caterpillars, but the moth pollinates more flowers than it lays eggs in. These mutualisms seem to have evolved both more than once and also some time after the divergence of the clade in which they are found - 55.2-33.4 m.y.a. (plant) versus 35-20 m.y. (moth) ago - and they have also been lost (Kawakita & Kato 2009; Kawakita 2010).
Vegetative Variation. Phyllanthoid branching occurs in many, but not all, species of Phyllanthus s.l. (Kathriarachchi et al. 2006). The orthotropic axes have reduced, spirally-arranged leaves and the plagiotropic axes usually have two-ranked, photosynthetic leaves and flowers in the axils of those leaves; the latter axes are of more or less limited growth. The plagiotropic lateral branches of P. acidus may be short-lived and lack flowers, so being the functional equivalent of compound leaves; the flowers themselves are borne on short branches lacking photosynthetic leaves and which arise from separate axillary buds. Some Caribbean species are yet more modified. Thus the plagiotropic lateral branches of P. epiphyllanthus are cladodes and bear flowers and fruits in the axils of scale leaves on either side of the cladode.
Chemistry, Morphology, etc. The only record of cocarcinogens is from one species of Antidesma (Beuteler et al. 1989). Wood anatomy is variable (Hayden & Brandt 1984) and the nodes are quite often unilacunar (Thakur & Patil 2002).
The inflorescence of Uapaca is a pseudanthium. The pollen is very variable in Phyllanthus in particular (Webster 1956). The outer integument is variable in thickness. The fruit type of the ancestor of Phyllanthaceae is unclear (Kathriarachchi et al. 2005). The exotegmen is most often described as being ribbon-like or tracheoidal. However, Hymenocardieae (Didymocistus, Hymenocardium) have a collapsed tracheoidal exotegmen and large tanniniferous endotegmic cells; do they belong here (Tokuoka & Tobe 2001)? - yes, say Wurdack et al. (2004) and Kathriarachchi et al. (2005).
For additional information, see also Köhler (1965: pollen), Hans (1973: chromosomes), Levin (1986: leaves), Mennega (1987: wood anatomy), Tokuoka and Tobe (2001: ovules and seeds), Westra and Koek-Noorman (2004: wood end-grain),León Enriquez et al. (2008: architectural variation in the family), and Zhang et al. (2012: floral morphology of Phyllanthus. For pollen, see Webster and Carpenter (2008) and Chen et al. (2009, both Phyllanthus, and Sagun and van der Ham (2003: pollen morphology of Flueggeinae).
Phylogeny. Phyllanthaceae include most of the old Euphorbiaceae-Phyllanthoideae, minus Drypetes and relatives, for which see Putranjivaceae. For phylogenetic relationships, see Wurdack et al. (2004: morphology also discussed, the odd genus Croizatia included), and Samuel et al. (2005: two gene analysis). Kathriarachchi et al. (2005: five-genes; see also Hoffmann et al. 2006) divide the family into two main clades, one largely with fasciculate inflorescences (it includes Lingelsheimia, sometimes previously associated with Putranjivaceae, and Dicoelia, ditto with Pandaceae), the Phyllanthoideae above, and the other (including Hymenocardieae) tanniniferous, the Antidesmatoideae above. For the phylogeny of Phyllanthus, see (Kathriarachchi et al. 2006; Pruesapan et al. 2008), for that of Poranthereae, see Voronstova et al. (2007).
Classification. Govaerts et al. (2000: as Euphorbiaceae) provide a checklist and bibliography, but this is now dated - e.g. see Vorontsova and Hoffmann (2008) for genera in Phyllanthoideae-Poranthereae, while Hoffmann et al. (2006) provide the phylogenetic classification followed here.
The limits of Cleistanthus will have to be adjusted, and most species will need a new name (Li et al. 2009), while the large genus Phyllanthus is paraphyletic, and its limits should probably be broadened to include Glochidion (some 300 species), Breynia, Sauropus (70 spp.), etc. (Kathriarachchi et al. 2006; Lorence & Wagner 2011 - but c.f. Pruesapan et al. 2008). For a monograph of Baccaurea and relatives, see Haegens (2000), and of Aporosa, see Schot (2004).
PICRODENDRACEAE Small, nom. cons. Back to Malpighiales
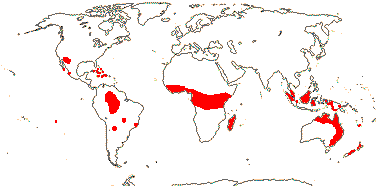
Picrotoxanes +, otherwise chemistry?; cork?; vessel elements with simple perforation plates; ?nodes; mucilage cells [epidermis] + (0); subsidiary cells piggy back [on top of guard cells]; hairs unicellular or unbranched uniseriate; leaves spiral, opposite, (two-ranked), lamina with secondary venation ± palmate or leaves palmate, margin toothed [teeth with deciduous apex] or entire, stipules petiolar, cauline, with axillary colleters, or 0; (plant dioecious); (disc interstaminal); staminate flowers: P 3(-13); A 2-many, (connate basally), often extrorse; nectary between or inside A; pollen 4-brevicolporate, 5-6-porate, zoni- or pantoaperturate, echinate to verrucose; pistillode +/0; carpellate flowers: P (3-)4-8(-13); G (2-)3(-5), stigmas stout, entire (multifid), dry (wet); (micropyle endostomal - Austrobuxus), outer integument 5-6 across, inner integument 3-6 cells across, nucellar cap +, (nucellar beak +), hypostase +, funicular obturator +, with hairs; (fruit indehiscent); seeds carunculate (0), vascular bundle branching in chalaza; exotegmen cuboid or fibrous, [palisade, subprocumbent, mesotegmen ± thickened, endotegmen 2-layered, with banded thickenings- Oldfieldia]; endosperm + (0), (ruminate - Picrodendron), embryo green, cotyledons broader than radicle (plicate; lobed - Picrodendron); (n = 12 - Pseudanthus).
24[list]/80 - three groups below. Many small genera; tropical, esp. New Guinea - Australia - New Caledonia or America plus Africa - Madagascar (map: from Webster 1994a; van Welzen & Forster 2011; FloraBase i.2012 - incomplete). [Photo - Picrodendron Fruit © A. Gentry.]
1. Podocalyx
?
1/1. Podocalyx loranthoides. Amazonia.
[Caletieae + Picrodendreae]: seeds carunculate.
2. Caletieae
?
Largely Australasian, New Caledonia.
Synonymy: Androstachyaceae Airy Shaw, Micrantheaceae J. Agardh, Paivaeusaceae A. Meeuse, Pseudanthaceae Endlicher
3. Picrodendreae
?
Africa, America
Evolution. Ecology & Physiology. The New World Piranhea may dominate flooded forests (Connell & Lowman 1989).
Seed Dispersal. Myrmecochory occurs in the majority of the species of the family (Lengyel et al. 2009, 2010).
Chemistry, Morphology, etc. The family is poorly known. Picrodendron may have a perianth of two whorls (Hakki 1985), or perhaps it is modified quincuncial.
Some information is taken from van Welzen & Forster (2010: general), Hayden (1977, 1994: wood anatomy), Levin and Simpson (1994: pollen), Huber (1991) and Tokuoka and Tobe (1999: seed anatomy), Wurdack et al. (2004: some morphology), Westra and Koek-Noorman (2004: wood end-grain), and Merino Sutter et al. (2006: morphology of carpellate flowers).
Phylogeny. For the taxa included in this family, see Euphorbiaceae-Oldfieldioideae (Webster 1994b). Paradrypetes is probably to be included in Rhizophoraceae, although it is like Podocalyx in particular in wood anatomy and pollen morphology (Levin 1992). Croizatia (see Levin 1992) is also odd, with 5 petals, an extrastaminal disc and style with distinct branches, and it lacks the distinctive pollen of the family; it is here placed in Phyllanthaceae (confirmed by molecular data: see Wurdack et al. 2004; Wurdack 2008). Stuppy (1996) noted that both Picrodendron and Oldfieldia were rather different from other taxa included in his Oldfieldioideae, and he compared the latter with Meliaceae because of similarity in seed characters. However, both genera are firmly in Picrodendraceae-Picrodendreae.
Classification. Govaerts et al. (2000) provides a checklist and bibliography (as Euphorbiaceae).
[Ixonanthaceae + Linaceae]: C contorted; ovules with endothelium, parietal tissue 2-5 cells across, hypostase +, placental obturator +; cotyledons large.
Phylogeny. This family pair has strong support (Xi et al. 2012: = linoids).
IXONANTHACEAE Miquel, nom. cons. Back to Malpighiales
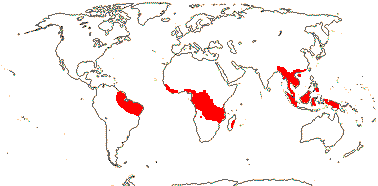
Woody; ellagic acid, myricetin; vessel elements with simple perforation plates; mucilage cells 0; cuticle waxes as variously arranged platelets; stomata paracytic; petiole bundle arcuate; branching from previous flush; leaves spiral, lamina vernation involute, (margins entire), stipules lateral; inflorescences axillary, corymbose; K usu. basally connate, (C imbricate); A folded in bud, 5, opposite sepals [no staminodes], -20 [in triplets opposite K]; pollen with supratectal spines; disc at bases of filaments, vascularized (unvascularized pads adaxial to filaments); G [(2) 5], (carpels subdivided), style undivided, slender, stigma capitate or discoid; micropyle bistomal, (outer integument very long); fruit a septicidal (and loculicidal) capsule opening adaxially as well, columella persistent or not, K and C persistent; seeds basally winged, or aril arising between the hilum and micropyle; endotegmen with sinuous anticlinal walls; endosperm scanty; n = 14, chromosomes 0.4-1.1 µm long.
3[list]/22. Pantropical, often in swamps, on white sand, etc. (map: from Aubréville 1974; Kool 1988).
Evolution. Divergence & Distribution. Stem Ixonanthaceae may have diverged in the Cretaceous-Albian 111-100 m.y.a. (Davis et al. 2005a).
Chemistry, Morphology, etc. The stamens opposite the petals in Ixonanthes are paired, but arise from a single trace (Narayana & Rao 1966). Narayana (1970) depicts a tegmen ca 4 cells thick, the innermost layer of which is an endothelium.
See also Kool (1980: revision of Ixonanthes, 1988), Forman (1965) and Kubitzki (2014), general, Nooteboom (1967: chemistry), Weberling et al. (1980: stipules), Narayana (1970) and Narayana and Rao (1966), embryology, etc., and Link (1992d: nectary).
Previous Relationships. Ixonanthaceae have been associated with a variety of families, although Van Hooren and Nooteboom (1988) noted that they had indeed often been assoociated with Linaceae. Thus Robson and Airy Shaw (1962) thought that the "spiral convolution of the filaments and style" of Cyrillopsis (Ixonanthaceae) was a point of similarity between this genus and Irvingiaceae. However, some species of Ochthocosmus also have flowers very similar to those of Cyrillopsis, with the thin calyx reflexed after anthesis (Phyllocosmus, Ixonanthes), while other species of Ochthocosmus have persistent, erect, almost scarious-looking sepals, as is common in Linaceae.
Although in Soltis et al. (2007a) Ixonanthes grouped strongly with the families around Salicaceae, this was because of misidentified material (= Hydnocarpus: K. Wurdack, pers. comm.).
LINACEAE Perleb, nom. cons. Back to Malpighiales
Cork?; vessel elements with simple or scalariform perforation plates; true tracheids +; petiole bundle(s) arcuate; cristarque cells + [rare in Linoideae]; epidermal wax crystals as parallel platelets; branching from previous innovation; lamina vernation usu. involute, tooth ?type, petiole short; pedicels articulated; (flowers distylous); K quincuncial, C postgenitally connate above base, caducous; A basally connate, basally adnate to C, anthers basifixed; pollen starchy; G [2-5], opposite petals, or median member adaxial, style more or less divided, stigmas ± capitate; ovule (1/carpel), epitropous, mictopyle endostomal (bistomal), endothelium +; (megaspore mother cells several); fruit often septicidal, K persistent; tegmen strongly multiplicative; endosperm with chalazal haustorium, variable, (embryo slightly curved), cotyledons large.
10-12[list]/300 - two groups below. World-wide.
1. Linoideae
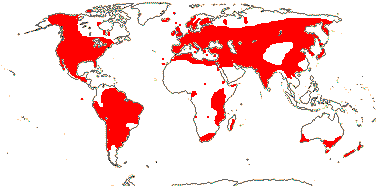
Usu. herbs (shrubs); ellagic acid 0; vessel elements with simple perforation plates; rays uniseriate; nodes 1:1 [Linum]; cuticle waxes as parallel platelets; leaves opposite or spiral, lamina margins entire or toothed, (stipules 0); K ± equal, C clawed, protective in late bud; nectary outside filaments or at base of C or A; A 5, opposite sepals, alternating with staminodes; pollen trinucleate, 3-many colpate or many colporate, or inaperturate, surface ± intectate, gemmate or echinate; G loculi usu. divided, stigma wet or dry; ovules with outer integument 2(-3) cells across, inner integument 3-12 cells across, parietal tissue 0, hypostase?, (lateral tissue scanty), (obturator with papillae); (2-seeded mericarps also splitting along false septae, units opening adaxially); seeds often mucilaginous, exotesta with outer walls massively thickened, cross cells beneath exotegmen, endosperm scanty, xyloglucans +, (helobial), embryo green [Linum]; n = 6, (8), 9, (11-18, etc.).
6/240: Linum (180). Worldwide, but esp. N. temperate and subtropical (map: from Hultén & Fries 1986; Diderichsen & Richards 2003; McDill et al. 2009 - East Asia?). [Photo - Flower]
2. Hugonioideae
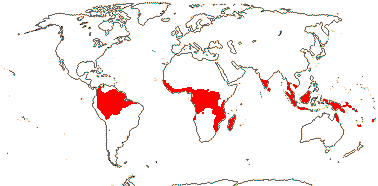
Woody, evergreen often lianes with branch grapnels; ellagic acid?; vessel elements with scalariform perforation plates; sclereids +; stomata accessory cells usu. lignified, lobed beneath the guard cells; branching from previous flush; leaves spiral or two-ranked, (stipules pectinate); (flowers tristylous); K often unequal, C at most slightly clawed, often yellow, (0); disc at base of filaments; A 10, of two lengths, (obdiplostemonous); (G with 1 of 3 loculi better developed than others/fertile; opposite K), styles impressed; ovule with micropyle endostomal [Roucheria], outer integument 2-3 cells thick, inner integument 3-5 cells thick, obturator with papillae; fruit a drupe or with mericarps; seed with an at most slight arillode, testa multiplicative, mesotesta with sclerotic cells, endotesta lignified, exotegmen barely lignified or tegmen obliterated; endosperm copious to scanty; n = 6, 12, 13.
4-6/61. Pantropical (map: from van Hooren & Nooteboom 1984a; Jardim 1999; McDill et al. 2009). [Photo - Flower]
Synonymy: Hugoniaceae Arnott
Evolution. Divergence & Distribution. Linaceae may have diverged from other Malpighiales in the Cretaceous-Albian 111-100 m.y.a. (Davis et al. 2005a), while McDill and Simspon (2011, q.v. for more details, Irvingiaceae taken as closest to Linaceae) suggest that crown group divergence is early Caenozoic (range 82-43 m.y.), Linoideae beginning to diversify in the Eocene and Hugonioideae in the Oligocene-Miocene.
Pollination Biology. Heterostylous flowers are scattered in Linoideae, and tristyly is reported from at least some Hugonioideae (Meeus et al. 2011), but whether heterostyly is an apomorphy for Linaceae is unclear; breeding systems have certainly been labile (Thompson et al. 1996; McDill et al. 2008, 2009).
Economic Importance. Seeds of flax (Linum usitatissimum) have been used for oil, etc., for about 10,000 years (Vaisey-Genser & Morris 2003).
Chemistry, Morphology, etc. Ellagic acid is not reported from Linoideae, but members of this subfamily are largely herbaceous. Flat vernation is reported from Linum narbonense by Cullen (1978), other taxa may be conduplicate. Bracts and bracteoles have a single vascular trace. Tirpitzia bilocularis has a corolla tube over 2 cm long. The "staminodes" may lack any vascularization (Al-Nowaihi & Khalifa 1973). Tobe and Raven (2011) suggest that the inner integument is multiplicative. In Hugonioideae, only half of the ovules may develop and produce seeds. Guinard (1893) drew the ovules of Linum as having a single parietal layer of cells; at the base of the embryo sac there was a persistent narrow column of cells. Note that septicidal dehiscence, presumably liberating pyrenes, may occur in this subfamily (Spichiger et al. 2002).
For information on the family and its possible segregates see Hegnauer (1966, 1989: chemistry), Boesewinkel (1980), L. L. Narayana (1970) and Narayana and Rao (1966, 1969, 1978a), all embryology, floral anatomy, etc., Robertson (1971: Linoideae), van Hooren and Nooteboom (1984, 1988a, b: general), van Welzen and Baas (1984: anatomy), Jardim (1999: New World Hugonioideae), Schmidt et al. (2010: lignans, in most Linum alone), and Schewe et al. (2011: floral development in Linum.
Phylogeny. McDill et al. (2009: focus on Linoideae) oulined phylogenetic relationships in the family; Linoideae may be monophyletic, but support is from posterior probabilities only; the status of Hugonioideae is unclear. However, in the more extended sudy of McDill and Simpson (2011) Linoideae are well supported, Hugonioideae monophyletic, but still lacking strong support,
Classification. If the topology suggested by McDill and Simpson (2011) is maintained in future studies, nomenclatural adjustments will be needed - perhaps best to expand Linum?
Previous relationships. For an early detailed discussion on relationships of Linaceae, then thought to be a "central" family, see Hallier (1923). Linaceae are weakly associated with Picrodendraceae in Chase et al. (2002a), and with Irvingiaceae in Tokuoka and Tobe (2006). Linaceae have been linked with Erythroxylaceae and Humiriaceae, and thence to Geraniales (Narayana & Rao 1978b), or the three families together are placed in Linales (Cronquist 1981).
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02362_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04996_001.jpg',600,500)){kind=link}
![[Photo - Fruit]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-03966_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04919_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04833_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-03677_001.jpg',600,500)){kind=link}
![[Photo - Flower]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-05131_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02075_001.jpg',600,500)){kind=link}
){kind=link}
![Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02103_001.jpg',600,500)){kind=link}
![[Photo - Flower]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04480_001.jpg',600,500)){kind=link}
![[Photo - Marathrum Flower]](javascript:showImage('http://images.mobot.org/mrsid/257/25700039_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02387_001.jpg',600,500)){kind=link}
![[Photo - Flower]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02009_001.jpg',600,500)){kind=link}
![[Photo - Flower, Fruit]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02012_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-03095_001.jpg',600,500)){kind=link}
){kind=link}
![[Photo - Flower]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04120_001.jpg',600,500)){kind=link}
![[Photo - Flower]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02269_001.jpg',600,500)){kind=link}
![[Photo - Fruit]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02265_001.jpg',600,500)){kind=link}
![[Photo - Flower]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02056_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02862_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04125_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04685_001.jpg',600,500)){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02667_001.jpg',600,500)){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04985_001.jpg',600,500)){kind=link}
){kind=link}
){kind=link}
){kind=link}
![Fruit.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02434_001.jpg',600,500)){kind=link}
![[Photo - Flower.]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02421_001.jpg',600,500)){kind=link}
![Photo - Picrodendron Fruit © A. Gentry.]](javascript:showImage('http://images.mobot.org/mrsid/128/MOA-02424_001.jpg',600,500)){kind=link}
![[Photo - Flower]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-04731_001.jpg',600,500)){kind=link}
![[Photo - Flower]](javascript:showImage('http://mobot.mobot.org/cgi-bin/search_vast?w3till=MOA-02994_001.jpg',600,500)){kind=link}